Fluorescent Lectins for Local in Vivo Visualization of Peripheral Nerves


Abstract
:1. Introduction

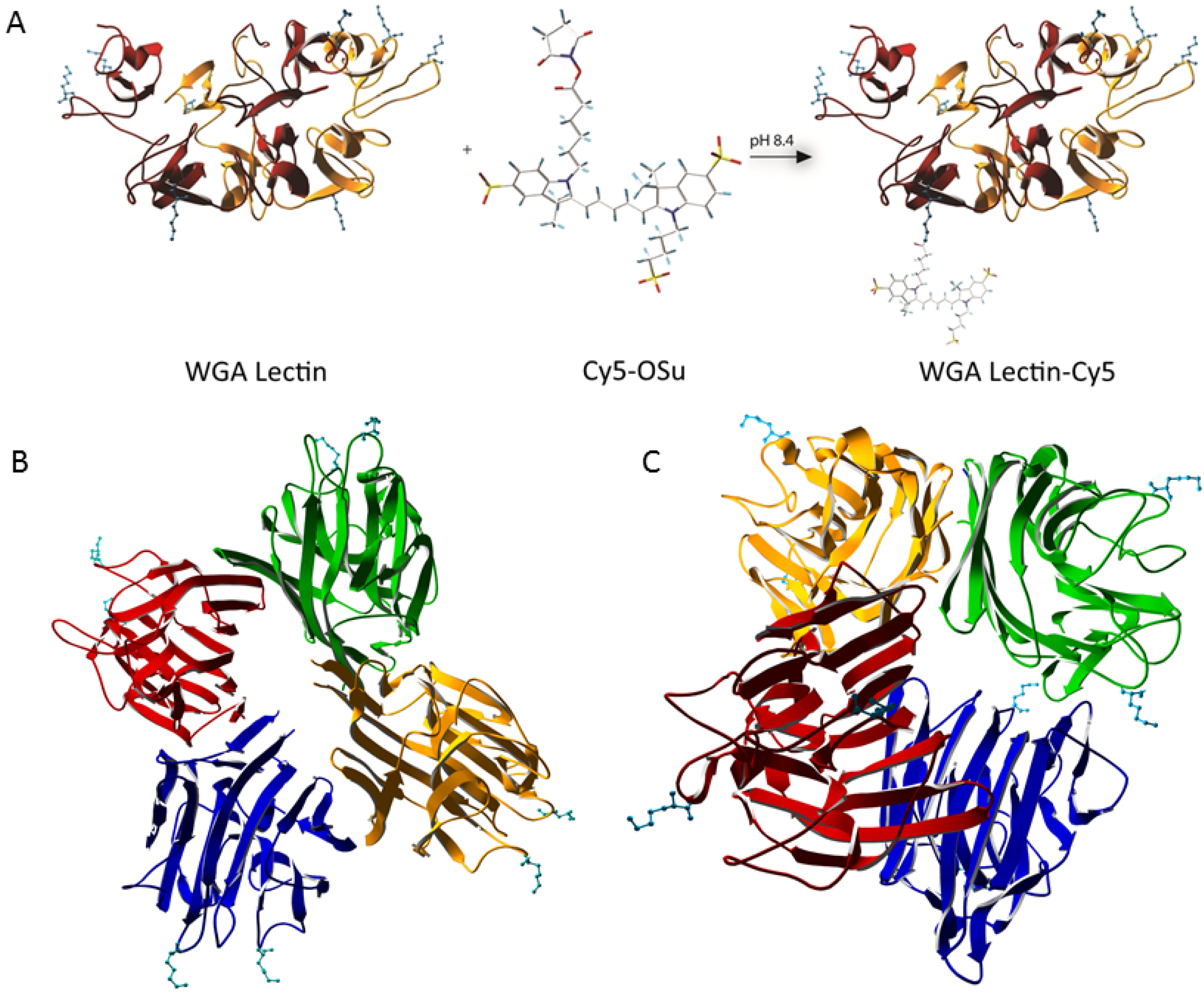{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lectin | Sugar Moiety | PG | Molecular Weight (KD) |
|---|---|---|---|
| Triticum Vulgaris (Wheat germ agglutinin; WGA); | β-d-GlcNAc, Neu5Ac | HSPG, KSPG, HP | 36 |
| Arachis Hypogaea (Peanut lectin; PNA) | Gal β (1–3)GalNAc | CSPG, DSPG | 110 |
| Phaseolus Vulgaris Leucoagglutinin(Red kidney bean; PHA-L) | Antennary branched β (1–6) GlcNAc | HSPG, KSPG, HP | 120 |
| Lycopersicon Esculentum (Tomato lectin; LEL) | [GlcNAc β (1–4)]2−4 | HSPG, KSPG, HP | 71 |
2. Results and Discussion
2.1. Labeling Efficacy
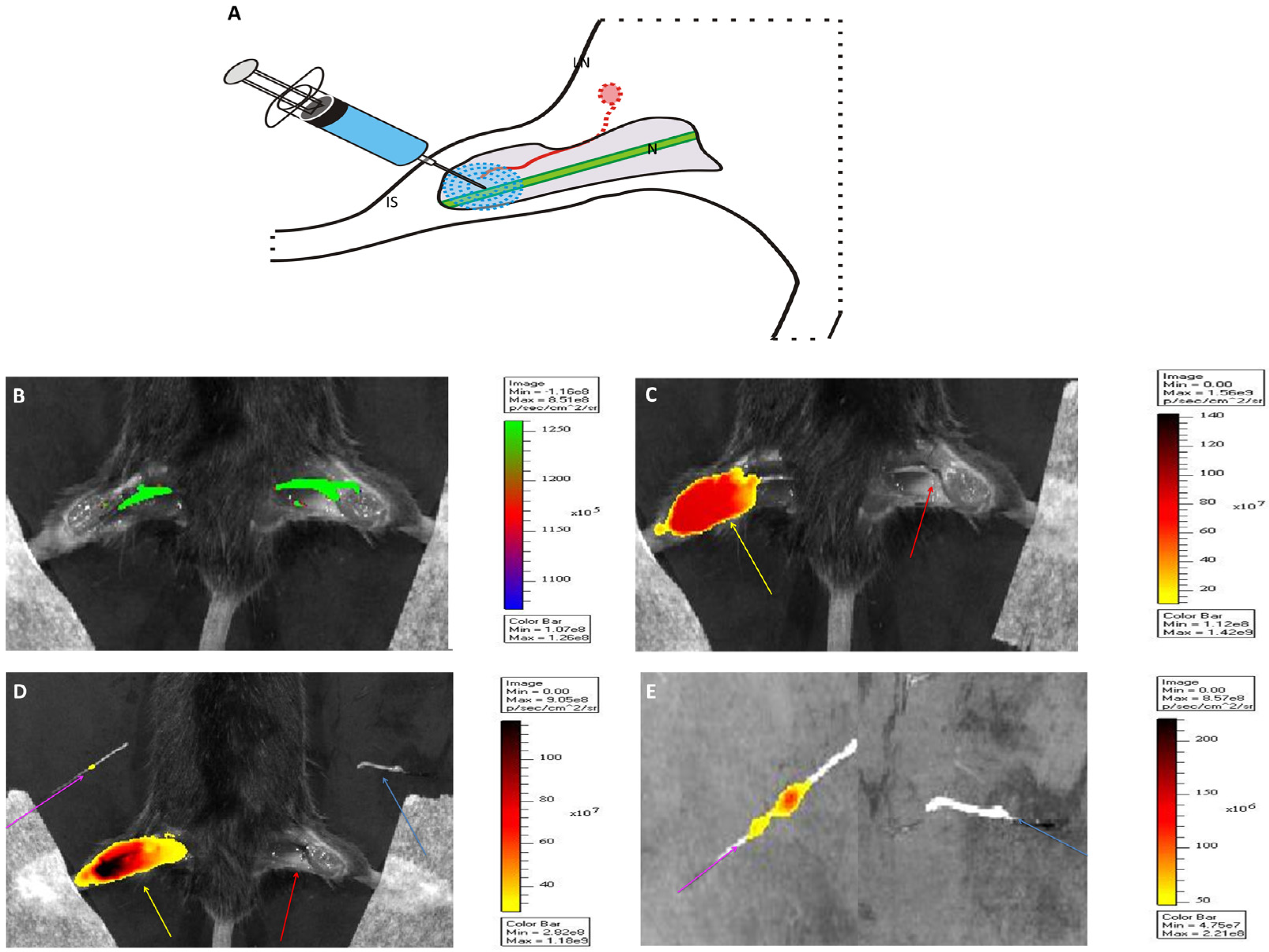
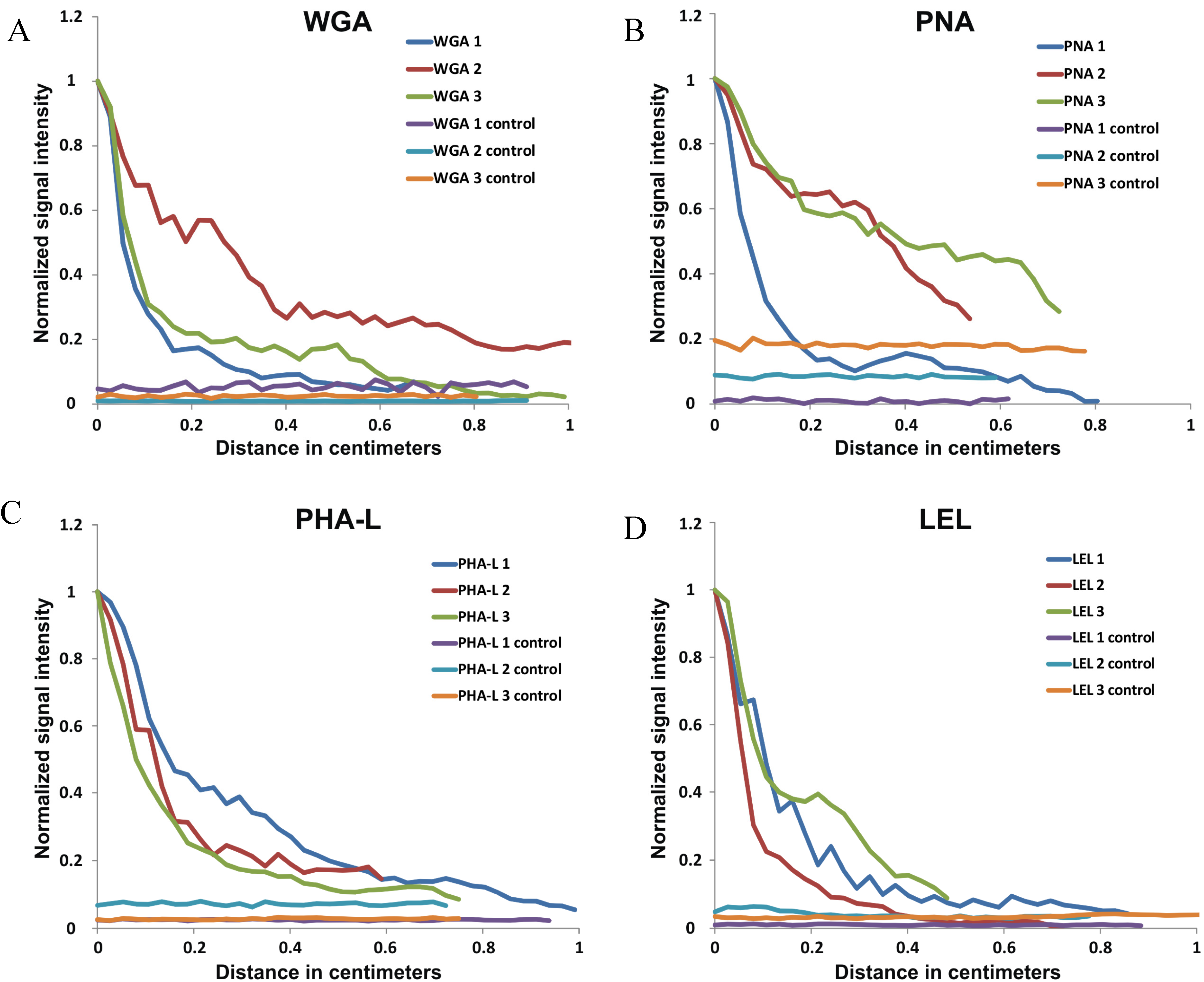
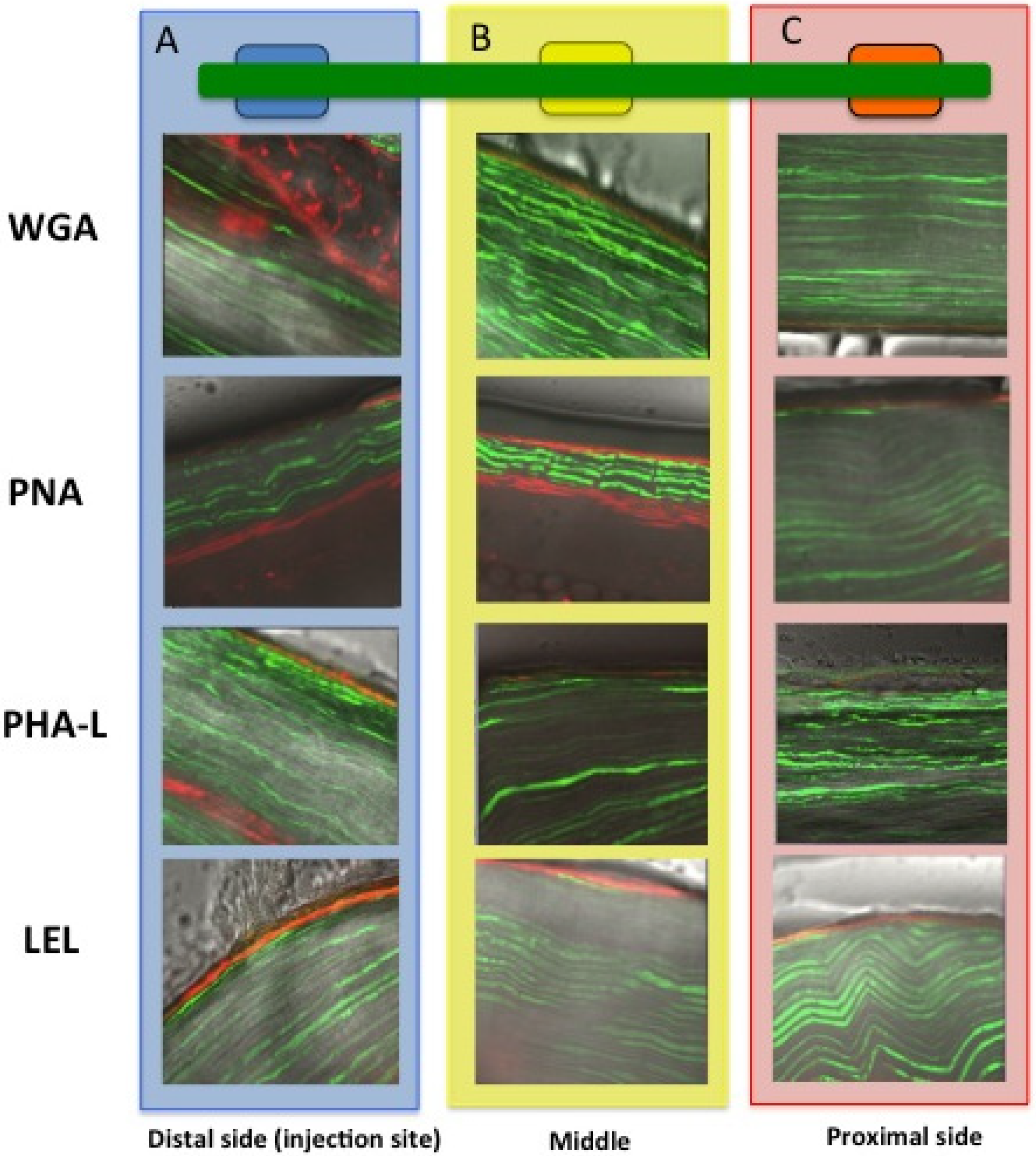
2.2. In Vivo Migration



| WGA | PNA | PHA-L | LEL | |
|---|---|---|---|---|
| Average migration distance (cm) (SD) | 0.95 (0.20) | 0.72 (0.13 | 0.81 (0.20) | 0.72 (0.20) |
| SBR nervecontrol (Average + SD) | 2.08 (1.11) | 1.72 (0.32) | 1.86 (0.70) | 4.88 (2.48) |
| SBR nervemuscle (Average + SD) | 1.86 (1.00) | 1.42 (0.12) | 1.12 (0.13) | 1.26 (0.46) |
| SBRIpsilateral lymph node (Average + SD) | 0.61 (0.26) | 0.70 (0.06) | 3.73 (1.36) | 1.11 (0.87) |
| SBRContralateral lymph node (Average + SD) | 0.45 (0.42) | 0.55 (0.17) | 0.79 (0.33) | 0.86 (0.62) |

2.3. Signal Intensity
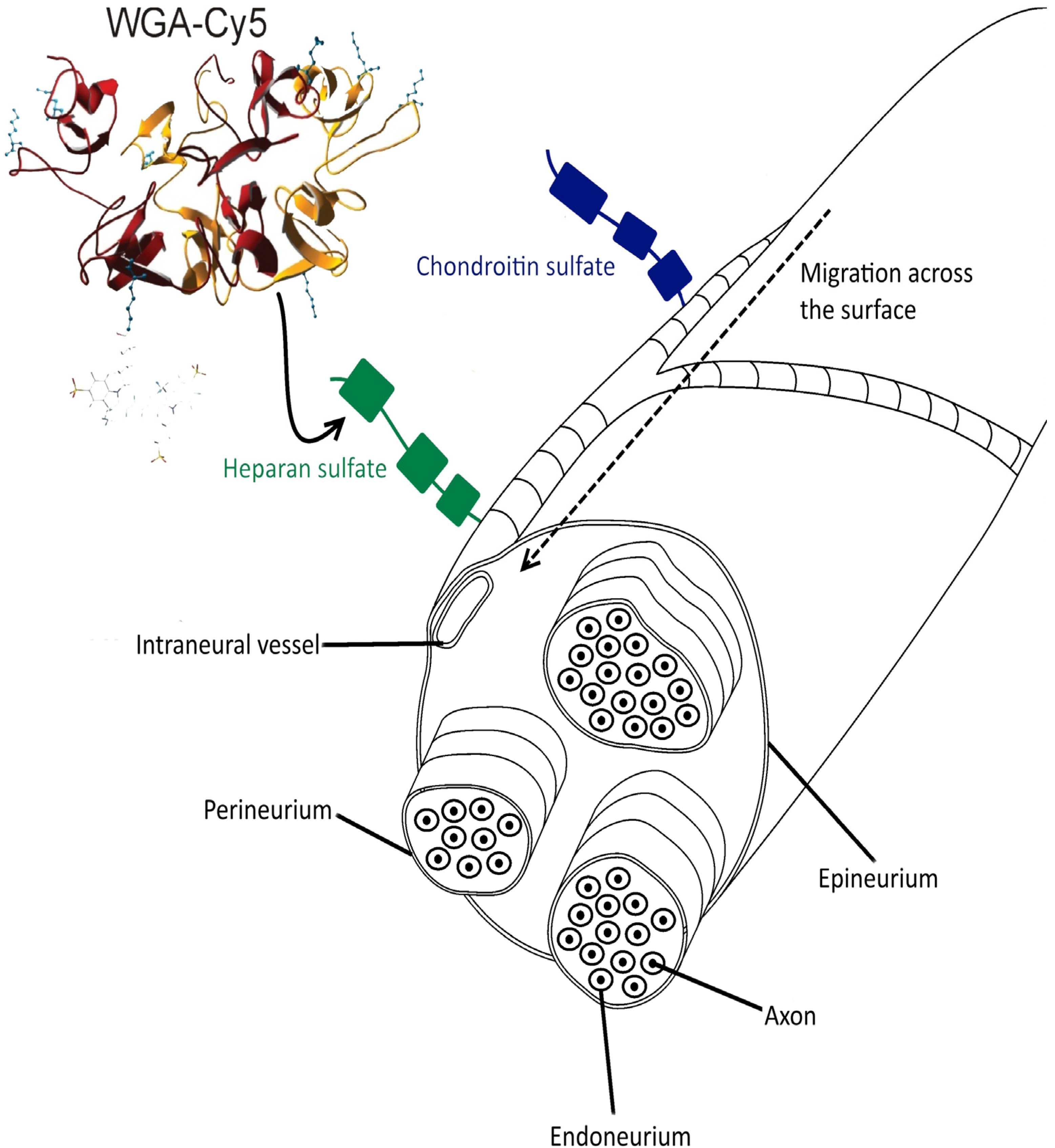
2.4. Evaluation of Binding Mode



3. Experimental Section
3.1. Experimental Equipment
3.2. Synthesis of Sulphonated Cy5-OSu
3.3. General Conjugation Procedure

| WGA | PNA | PHA-L | LEL | |
|---|---|---|---|---|
| Labeling ratio from test-conjugation | 1.08 | 1.25 | 0.75 | 1.48 |
| Amount of lectin (nmol) | 27.8 | 9.1 | 4.2 | 14.1 |
| Amount of fluorophore (nmol) | 191.8 | 54.6 | 35.3 | 70.5 |
| Equivalents of dye/lectin | 6.9 | 6.0 | 9.9 | 5.0 |


| WGA | PNA | PHA-L | LEL | |
|---|---|---|---|---|
| Measured absorption | 0.068 | 0.019 | 0.008 | 0.028 |
| Fluorophore concentration (nmol/μL) | 0.272 | 0.076 | 0.032 | 0.112 |
| Total sample volume (µL) | 139 | 139 | 128 | 150 |
| Total amount of fluorophore (nmoL) | 37.81 | 10.56 | 4.10 | 16.80 |
| Total amount of lectin (nmoL) | 27.80 | 9.10 | 4.15 | 14.10 |
3.4. Labeling Ratio Analysis
3.5. In Vivo Distribution
3.6. Ex Vivo Incubation and Fluorescence Confocal Microscopy
4. Conclusions
Acknowledgements
Author Contributions
Conflicts of Interests
References
- Mauroy, B.; Demondion, X.; Drizenko, A.; Goullet, E.; Bonnal, J.L.; Biserte, J.; Abbou, C. The inferior hypogastric plexus (pelvic plexus): Its importance in neural preservation techniques. Surg. Radiol. Anat. 2003, 25, 6–15. [Google Scholar] [CrossRef]
- Stelzner, S.; Holm, T.; Moran, B.J.; Heald, R.J.; Witzigmann, H.; Zorenkov, D.; Wedel, T. Deep pelvic anatomy revisited for a description of crucial steps in extralevator abdominoperineal excision for rectal cancer. Dis. Colon. Rectum. 2011, 54, 947–957. [Google Scholar] [CrossRef]
- Tewari, A.K.; Srivastava, A.; Huang, M.W.; Robinson, B.D.; Shevchuk, M.M.; Durand, M.; Sooriakumaran, P.; Grover, S.; Yadav, R.; Mishra, N.; et al. Anatomical grades of nerve sparing: A risk-stratified approach to neural-hammock sparing during robot-assisted radical prostatectomy (RARP). BJU Int. 2011, 108, 984–992. [Google Scholar] [CrossRef]
- Pfister, B.J.; Gordon, T.; Loverde, J.R.; Kochar, A.S.; Mackinnon, S.E.; Cullen, D.K. Biomedical engineering strategies for peripheral nerve repair: Surgical applications, state of the art, and future challenges. Crit. Rev. Biomed. Eng. 2011, 39, 81–124. [Google Scholar] [CrossRef]
- Myssiorek, D. Recurrent laryngeal nerve paralysis: Anatomy and etiology. Otolaryngol. Clin. North. Am. 2004, 37, 25–44. [Google Scholar] [CrossRef]
- Wei, J.T.; Dunn, R.L.; Marcovich, R.; Montie, J.E.; Sanda, M.G. Prospective assessment of patient reported urinary continence after radical prostatectomy. J. Urol. 2000, 164, 744–748. [Google Scholar] [CrossRef]
- Guntinas-Lichius, O. The facial nerve in the presence of a head and neck neoplasm: Assessment and outcome after surgical management. Curr. Opin. Otolaryngol. Head Neck Surg. 2004, 12, 133–141. [Google Scholar]
- Polom, K.; Murawa, D.; Rho, Y.S.; Nowaczyk, P.; Hünerbein, M.; Murawa, P. Current trends and emerging future of indocyanine green usage in surgery and oncology: A literature review. Cancer 2011, 117, 4812–4822. [Google Scholar] [CrossRef]
- Van der Poel, H.G.; Buckle, T.; Brouwer, O.R.; Valdés Olmos, R.A.; van Leeuwen, F.W.B. Intraoperative laparoscopic fluorescence guidance to the sentinel lymph node in prostate cancer patients: Clinical proof of concept of an integrated functional imaging approach using a multimodal tracer. Eur. Urol. 2011, 60, 826–833. [Google Scholar] [CrossRef]
- Brouwer, O.R.; van den Berg, N.S.; Mathéron, H.M.; van der Poel, H.G.; van Rhijn, B.W.; Bex, A.; van Tinteren, H.; Valdés Olmos, R.A.; van Leeuwen, F.W.; Horenblas, S. A hybrid radioactive and fluorescent tracer for sentinel node biopsy in penile carcinoma as a potential replacement for blue dye. Eur. Urol. 2014, 65, 600–609. [Google Scholar] [CrossRef]
- Gibbs-Strauss, S.L.; Nasr, K.A.; Fish, K.M.; Khullar, O.; Ashitate, Y.; Siclovan, T.M.; Johnson, B.F.; Barnhardt, N.E.; Tan Hehir, C.A.; Frangioni, J.V. Nerve-highlighting fluorescent contrast agents for image-guided surgery. Mol. Imaging. 2011, 10, 91–101. [Google Scholar]
- Kuil, J.; Buckle, T.; Oldenburg, J.; Yuan, H.; Borowsky, A.D.; Josephson, L.; van Leeuwen, F.W. Hybrid peptide dendrimers for imaging of chemokine receptor 4 (CXCR4) expression. Mol. Pharm. 2011, 8, 2444–2532. [Google Scholar] [CrossRef]
- Bunschoten, A.; Buckle, T.; Visser, N.L.; Kuil, J.; Yuan, H.; Josephson, L.; Vahrmeijer, A.L.; van Leeuwen, F.W. Multimodal interventional molecular imaging of tumor margins and distant metastases by targeting αvβ3 integrin. Chembiochem 2012, 13, 1039–1045. [Google Scholar] [CrossRef]
- Whitney, M.A.; Crisp, J.L.; Nguyen, L.T.; Friedman, B.; Gross, L.A.; Steinbach, P.; Tsien, R.Y.; Nguyen, Q.T. Fluorescent peptides highlight peripheral nerves during surgery in mice. Nat. Biotechnol. 2011, 29, 352–356. [Google Scholar] [CrossRef]
- Schellingerhout, D.; le Roux, L.G.; Bredow, S.; Gelovani, J.G. Fluorescence imaging of fast retrograde axonal transport in living animals. Mol. Imaging 2009, 8, 319–329. [Google Scholar]
- Streit, W.J.; Schulte, B.A.; Balentine, D.J.; Spicer, S.S. Histochemical localization of galactose-containing glycoconjugates in sensory neurons and their processes in the central and peripheral nervous system of the rat. J. Histochem. Cytochem. 1985, 33, 1042–1052. [Google Scholar] [CrossRef]
- Borges, L.F.; Sidman, R.L. Axonal transport of lectins in the peripheral nervous system. J. Neurosci. 1982, 2, 647–653. [Google Scholar]
- Zhou, C.J.; Kawabuchi, M.; He, J.W.; Kuraoka, A.; Hirata, K.; Wang, S.; Nada, O. Changes in the distribution of peanut agglutinin (PNA) binding molecules during muscle reinnervation following nerve crush injury. Arch. Histol. Cytol. 1999, 62, 261–272. [Google Scholar] [CrossRef]
- Gerfen, C.R.; Sawchenko, P.E. An anterograde neuroanatomical tracing method that shows the detailed morphology of neurons, their axons and terminals: Immunohistochemical localization of an axonally transported plant lectin, Phaseolus vulgaris leucoagglutinin (PHA-L). Brain Res. 1984, 290, 219–238. [Google Scholar] [CrossRef]
- Roth, J.; Binder, M.; Gerhard, U.J. Conjugation of lectins with fluorochromes: An approach to histochemical double labeling of carbohydrate components. Histochemistry 1978, 56, 265–273. [Google Scholar] [CrossRef]
- Shehab, S.A.S.; Hughes, D.I. Simultaneous identification of unmyelinated and myelinated primary somatic afferents by co-injection of isolectin B4 and Cholera toxin subunit B into the sciatic nerve of the rat. J. Neurosci. Methods 2011, 198, 213–221. [Google Scholar] [CrossRef]
- Mislovičová, D.; Gemeiner, P.; Kozarova, A.; Kožár, T.; Lectinomics, I. Relevance of exogenous plant lectins in biomedical diagnostics. Biologia 2009, 64, 1–19. [Google Scholar] [CrossRef]
- Ruoslahti, E. Structure and biology of proteoglycans. Ann. Rev. Cell. Biol. 1988, 4, 229–255. [Google Scholar] [CrossRef]
- Scott, J.E. Proteoglycan-fibrillar collagen interactions. Biochem. J. 1988, 252, 313–323. [Google Scholar]
- Reitsma, S.; Slaaf, D.W.; Vink, H.; van Zandvoort, M.A.J.M.; Oude Egbrink, M.G.A. The endothelial glycocalyx: Composition, functions, and visualization. Pflugers Arch. Eur. J. Physiol. 2007, 454, 345–359. [Google Scholar] [CrossRef]
- Schwefel, D.; Maierhofer, C.; Beck, J.G.; Seeberger, S.; Diederichs, K.; Moller, H.M.; Welte, W.; Wittman, V. Structural basis of multivalent binding to wheat germ agglutinin. J. Am. Chem. Soc. 2010, 132, 8704–8719. [Google Scholar] [CrossRef]
- Natchiar, S.K.; Srinivas, O.; Mitra, N.; Surolia, A.; Jayaraman, N.; Vijayan, M. Structural studies on peanut lectin complexed with disaccharides involving different linkages: Further insights into the structure and interactions of the lectin. Acta Crystallogr. Sect. D: Biol. Crystallogr. 2006, 62, 1413–1421. [Google Scholar] [CrossRef]
- Hamelryck, T.W.; Dao-Thi, M.H.; Poortmans, F.; Chrispeels, M.J.; Wyns, L.; Loris, R. The crystallographic structure of phytohemagglutinin-L. J. Biol. Chem. 1996, 27, 20479–20485. [Google Scholar]
- Burggraaf, J.; Kamerling, I.M.C.; Gordon, P.B.; Schrier, L.; de Kam, M.L.; Kales, A.J.; Bendiksen, R.; Indrevoll, B.; Bjerke, R.M.; Moestue, S.A.; et al. Detection of colorectal neoplasia in vivo in humans using an intravenously administered fluoresecent peptide targeted against c-Met and fluorescent colonoscopy; a proof of concept study. Nature Med. 2014. accepted for publishing. [Google Scholar]
- Stryer, L. Fluorescence energy transfer as a spectroscopic ruler. Annu. Rev. Biochem. 1978, 47, 819–846. [Google Scholar] [CrossRef]
- Nieweg, O.E.; Tanis, P.J.; Kroon, B.B. The definition of a sentinel node. Ann. Surg. Oncol. 2001, 8, 538–541. [Google Scholar] [CrossRef]
- KleinJan, G.H.; Bunschoten, A.; Brouwer, O.R.; Berg, N.S.; Valdés-Olmos, R.A.; Leeuwen, F.W.B. Multimodal imaging in radioguided surgery. Clin. Transl. Imaging. 2013, 1, 433–444. [Google Scholar] [CrossRef]
- Stoffels, I.; Poeppel, T.; Boy, C.; Mueller, S.; Wichmann, F.; Dissemond, J.; Schadendorf, D.; Rosenbaum-Krumme, S.; Klode, J. Radio-guided surgery: Advantages of a new portable γ-camera (Sentinella) for intraoperative real time imaging and detection of sentinel lymph nodes in cutaneous malignancies. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 308–313. [Google Scholar] [CrossRef]
- Vermeeren, L.; Valdés Olmos, R.A.; Meinhardt, W.; Horenblas, S. Intraoperative imaging for sentinel node identification in prostate carcinoma: Its use in combination with other techniques. J. Nucl. Med. 2011, 52, 741–744. [Google Scholar] [CrossRef]
- Brouwer, O.R.; Klop, W.M.C.; Buckle, T.; Vermeeren, L.; van den Brekel, M.W.M.; Balm, A.J.M.; Nieweg, O.E.; Valdés Olmos, R.A.; van Leeuwen, F.W. Feasibility of sentinel node biopsy in head and neck melanoma using a hybrid radioactive and fluorescent tracer. Ann. Surg. Oncol. 2012, 19, 1988–1994. [Google Scholar] [CrossRef]
- Van den Berg, N.S.; Brouwer, O.R.; Klop, W.M.; Karakullukcu, B.; Zuur, C.L.; Tan, I.B.; Balm, A.J.; van den Brekel, M.W.; Valdés Olmos, R.A.; van Leeuwen, F.W. Concomitant radio- and fluorescence-guided sentinel lymph node biopsy in squamous cell carcinoma of the oral cavity using ICG-(99m)Tc-nanocolloid. Eur. J. Nucl. Med. Mol. Imaging. 2012, 39, 1128–1136. [Google Scholar] [CrossRef]
- Buckle, T.; Chin, P.T.K.; van den Berg, N.S.; Loo, C.E.; Koops, W.; Gilhuijs, K.G.A.; van Leeuwen, F.W. Tumor bracketing and safety margin estimation using multimodal marker seeds: A proof of concept. J. Biomed. Opt. 2010, 15, 056021. [Google Scholar] [CrossRef]
- Maravilla, K.R.; Bowen, B.C. Imaging of the peripheral nervous system: Evaluation of peripheral neuropathy and plexopathy. AJNR Am. J. Neuroradiol. 1998, 19, 1011–1023. [Google Scholar]
- Kostrominova, T.Y. Application of WGA lectin staining for visualization of the connective tissue in skeletal muscle, bone, and ligament/tendon studies. Microsc. Res. Tech. 2011, 74, 18–22. [Google Scholar] [CrossRef]
- Kaemmer, D.; Bozkurt, A.; Otto, J.; Junge, K.; Klink, C.; Weis, J.; Sellhaus, B.; O’Dey, D.M.; Pallua, N.; Jansen, M.; Schumpelick, V.; Klinge, U. Evaluation of tissue components in the peripheral nervous system using Sirius red staining and immunohistochemistry: A comparative study (human, pig, rat). J. Neurosci. Methods 2010, 190, 112–116. [Google Scholar] [CrossRef]
- Aquino, D.A.; Margolis, R.U.; Margolis, R.K. Immunocytochemical localization of a chondroitin sulfate proteoglycan in nervous tissue. I. Adult brain, retina, and peripheral nerve. J. Cell. Biol. 1984, 99, 1117–1129. [Google Scholar] [CrossRef]
- Carey, D.J.; Stahl, R.C.; Asundi, V.K.; Tucker, B. Processing and subcellular distribution of the Schwann cell lipid-floated heparan sulfate proteoglycan and identification as glypican. Exp. Cell. Res. 1993, 208, 10–18. [Google Scholar] [CrossRef]
- Hoedt-Schmidt, S.; McClure, J.; Jasani, M.K.; Kalbhen, D.A. Immunohistochemical localization of articular cartilage proteoglycan and link protein in situ using monoclonal antibodies and lectin-binding methods. Histochemistry 1993, 99, 391–403. [Google Scholar] [CrossRef]
- Mujumdar, R.B.; Ernst, L.A.; Mujumdar, S.R.; Lewis, C.J.; Waggoner, A.S. Cyanine dye labeling reagents: Sulfoindocyanine succinimidyl esters. Bioconjug Chem. 1993, 4, 105–111. [Google Scholar] [CrossRef]
- Sample Availability: Only via a research agreement with the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
KleinJan, G.H.; Buckle, T.; Van Willigen, D.M.; Oosterom, M.N.v.; Spa, S.J.; Kloosterboer, H.E.; Van Leeuwen, F.W.B. Fluorescent Lectins for Local in Vivo Visualization of Peripheral Nerves. Molecules 2014, 19, 9876-9892. https://doi.org/10.3390/molecules19079876
KleinJan GH, Buckle T, Van Willigen DM, Oosterom MNv, Spa SJ, Kloosterboer HE, Van Leeuwen FWB. Fluorescent Lectins for Local in Vivo Visualization of Peripheral Nerves. Molecules. 2014; 19(7):9876-9892. https://doi.org/10.3390/molecules19079876
Chicago/Turabian StyleKleinJan, Gijs Hendrik, Tessa Buckle, Danny Michel Van Willigen, Matthias Nathanaël van Oosterom, Silvia Johara Spa, Harmen Egbert Kloosterboer, and Fijs Willem Bernhard Van Leeuwen. 2014. "Fluorescent Lectins for Local in Vivo Visualization of Peripheral Nerves" Molecules 19, no. 7: 9876-9892. https://doi.org/10.3390/molecules19079876
APA StyleKleinJan, G. H., Buckle, T., Van Willigen, D. M., Oosterom, M. N. v., Spa, S. J., Kloosterboer, H. E., & Van Leeuwen, F. W. B. (2014). Fluorescent Lectins for Local in Vivo Visualization of Peripheral Nerves. Molecules, 19(7), 9876-9892. https://doi.org/10.3390/molecules19079876