
Diversity of Heparan Sulfate and HSV Entry: Basic Understanding and Treatment Strategies


Abstract
:1. Introduction


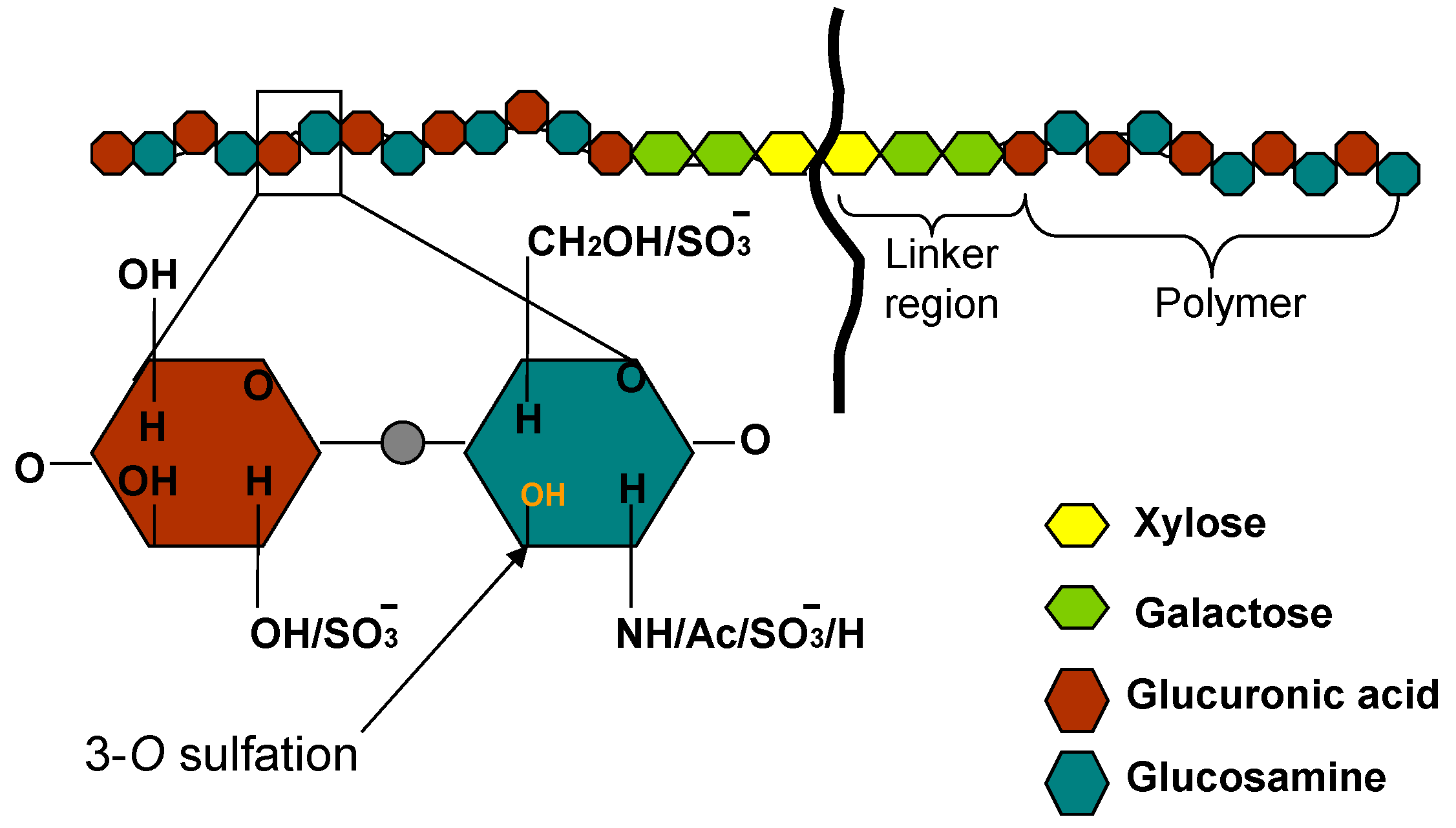{kind=link}
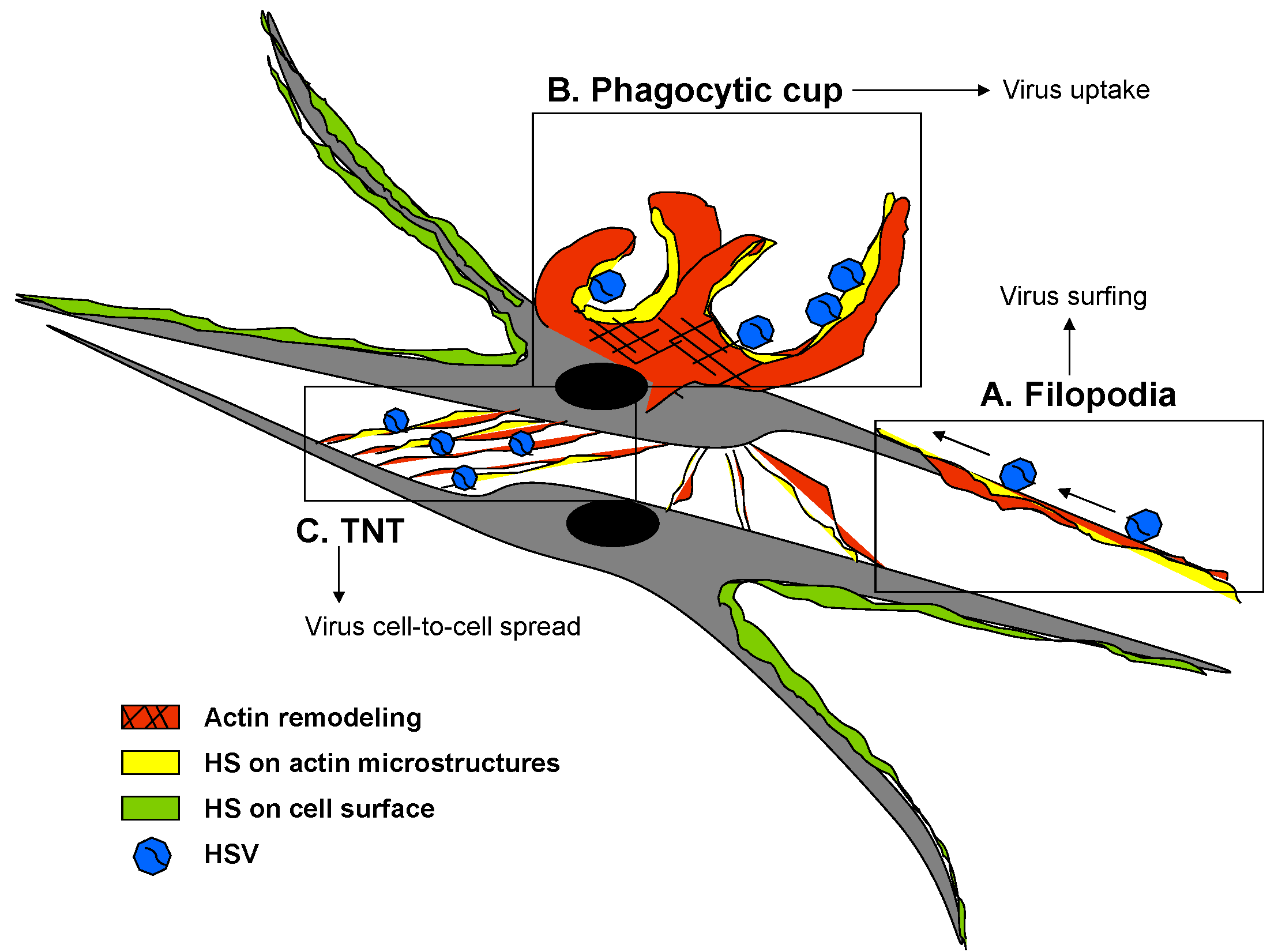{kind=link}
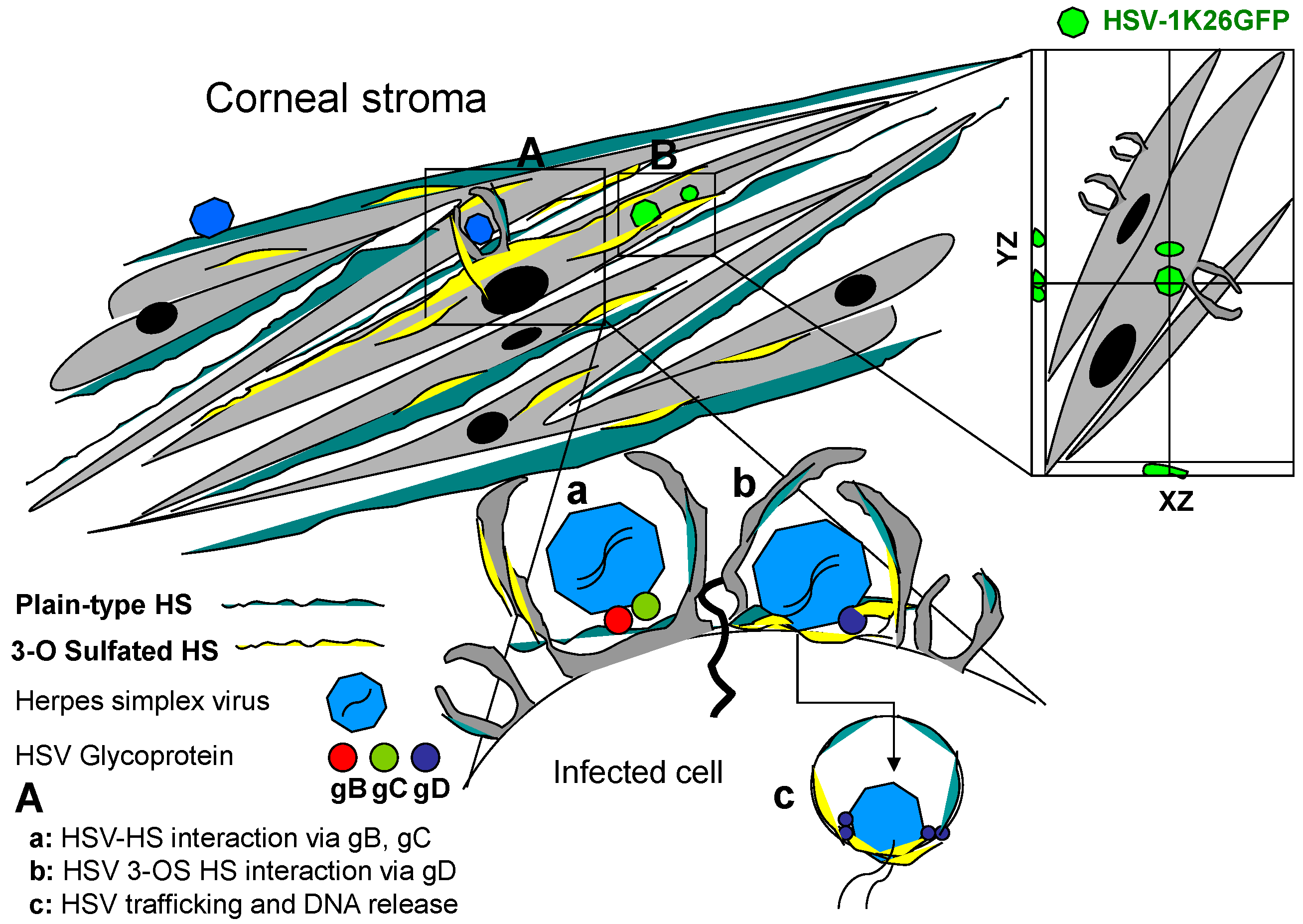{kind=link}
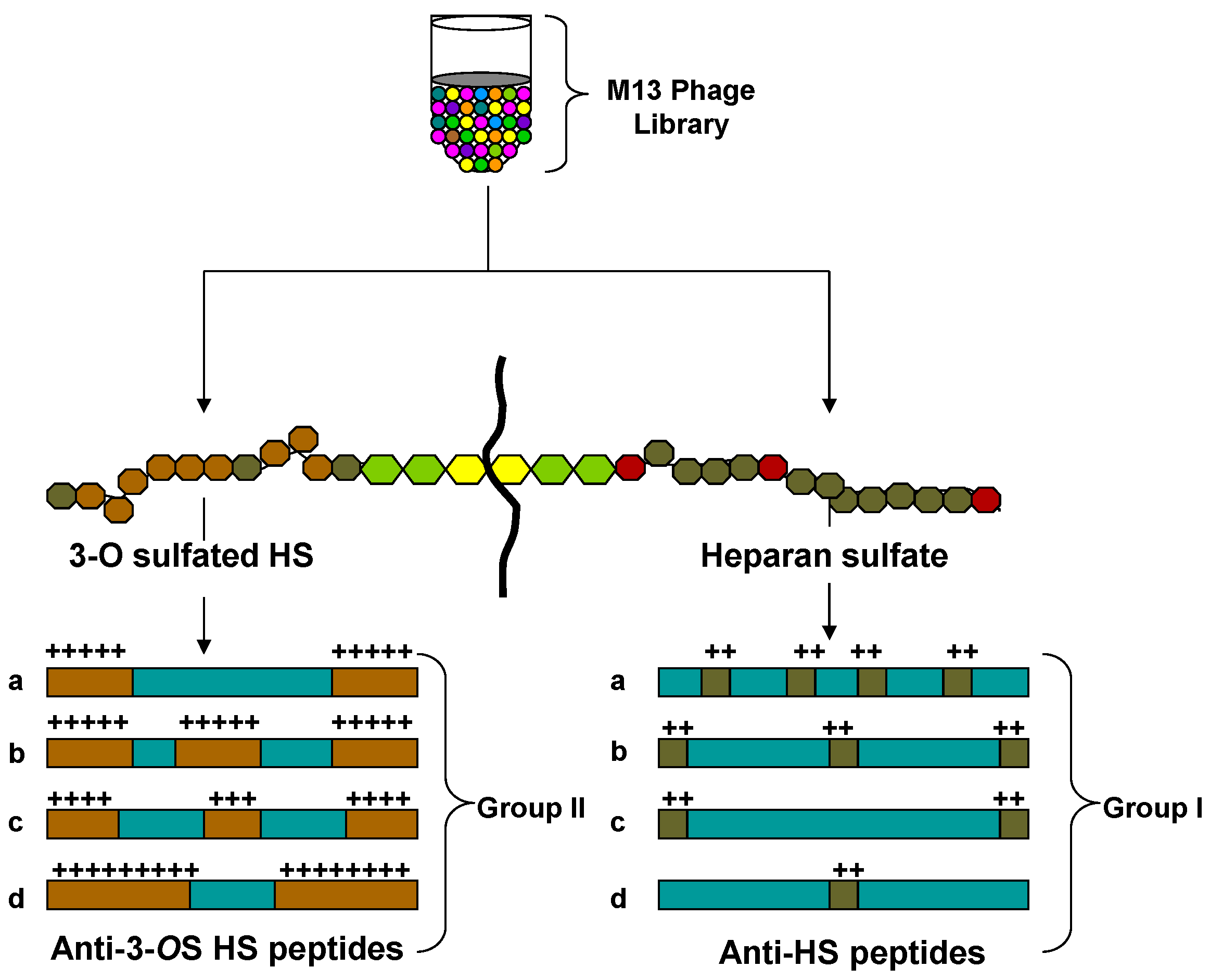{kind=link}
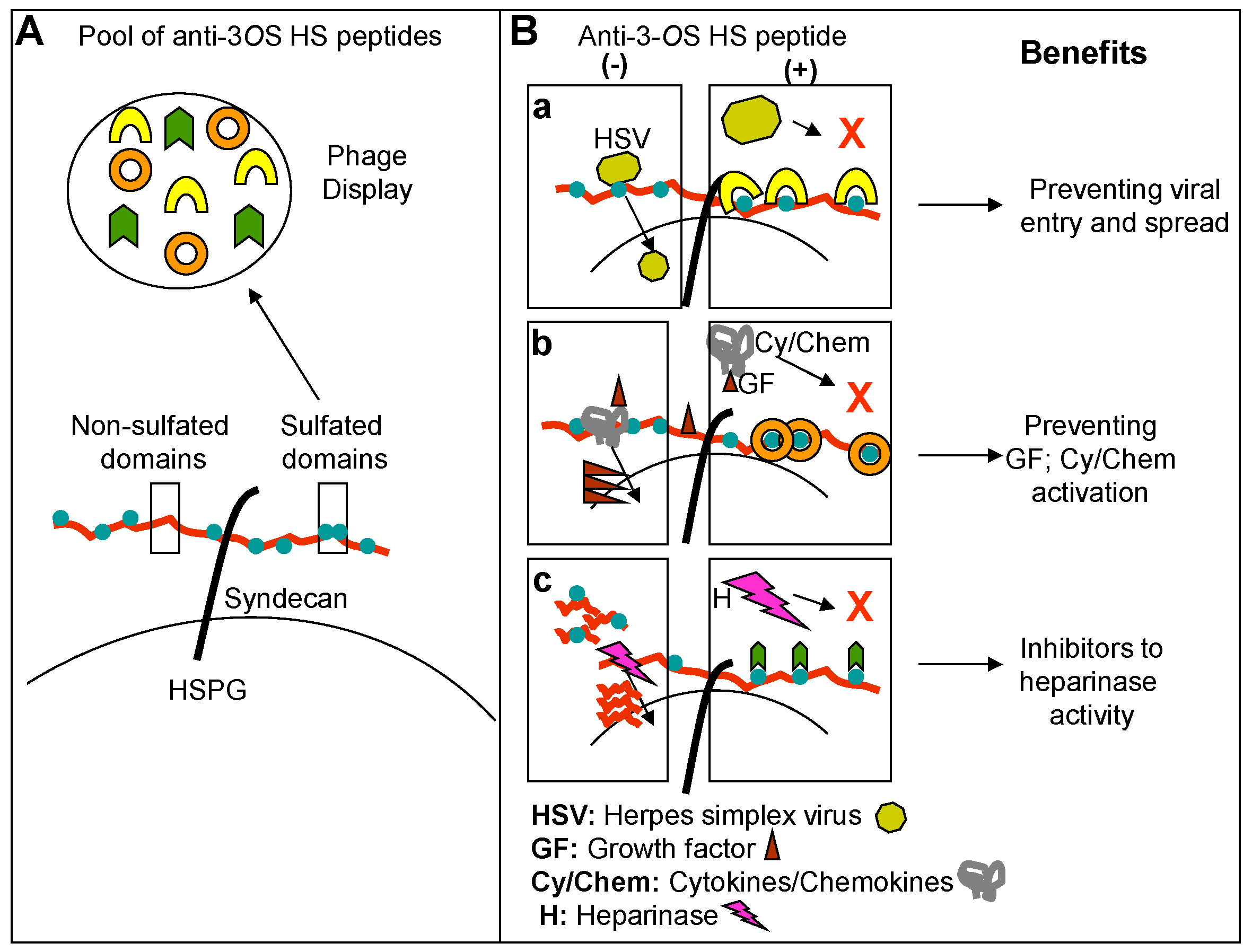{kind=link}
| HS Modifying Enzymes | Viral Infections | References |
|---|---|---|
| * H 3-O-Sulfotransferases-2, -3, -4, -5, -6 | Herpes simplex virus infection | [6,39,41] |
| * ZF 3-O-Sulfotransferases-2, -3, -4, -5, -6 | Herpes simplex virus infection | [48,49,50] |
| 3-O-Sulfotransferase-1 | Herpetic infection of the eye | [51] |
| 3-O-Sulfotransferase-1 | Hepatitis B replication | [46] |
| 6-O-Sulfotransferase | Cytomegalovirus infection | [47] |
| 6-O-Sulfotransferase | Coxsackievirus B3 internalization | [52] |
| 6-O-Sulfotransferase | Baculovirus binding and entry | [53] |
| 6-O-Sulfotransferase | Hepatitis C virus tropism | [54] |
| 2-O-Sulfotransferase | Human immunodeficiency virus entry | [55] |

2. Structural Diversity of HS and Implications in Corneal HSV Infection

3. Zebrafish 3-OST Generated HS: A Tool to Study HSV Corneal Damage
4. Phage Display Library Screening Targeting Heterogeneous HS to Isolate Unique Peptides that Inhibit HSV Pathogenesis

5. Anti-HS and Anti-3-OS HS Peptide: A Novel Tool to Study Virus Associated Inflammation

6. Conclusions
Acknowledgments
Conflicts of Interest
References
- Sarrazin, S.; Lamanna, W.C.; Esko, J.D. Heparan sulfate proteoglycans. Cold Spring Harb. Perspect. Biol. 2011, 3. [Google Scholar] [CrossRef]
- Nadanaka, S.; Kitagawa, H. Heparan sulphate biosynthesis and disease. J. Biochem. 2008, 144, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Bishop, J.R.; Schuksz, M.; Esko, J.D. Heparan sulphate proteoglycans fine-tune mammalian physiology. Nature 2007, 446, 1030–1037. [Google Scholar] [CrossRef] [PubMed]
- Esko, J.D.; Lindahl, U. Molecular diversity of heparan sulfate. J. Clin. Investig. 2001, 108, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Thacker, B.E.; Xu, D.; Lawrence, R.; Esko, J.D. Heparan sulfate 3-O-sulfation: A rare modification in search of a function. Matrix Biol. 2014, 35, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Shukla, D.; Spear, P.G. Herpesviruses and heparan sulfate: An intimate relationship in aid of viral entry. J. Clin. Investig. 2001, 108, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, V.; Maus, E.; Sigar, I.M.; Ramsey, K.H.; Shukla, D. Role of heparan sulfate in sexually transmitted infections. Glycobiology 2012, 22, 1402–1412. [Google Scholar] [CrossRef] [PubMed]
- Park, P.J.; Shukla, D. Role of heparan sulfate in ocular diseases. Exp. Eye Res. 2013, 110, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, U.; Kusche-Gullberg, M.; Kjellén, L. Regulated diversity of heparan sulfate. J. Biol. Chem. 1998, 273, 24979–24982. [Google Scholar] [CrossRef]
- Lindahl, U.; Li, J.P. Interactions between heparan sulfate and proteins-design and functional implications. Int. Rev. Cell Mol. Biol. 2009, 276, 105–159. [Google Scholar] [PubMed]
- Kreuger, J.; Jemth, P.; Sanders-Lindberg, E.; Eliahu, L.; Ron, D.; Basilico, C.; Salmivirta, M.; Lindahl, U. Fibroblast growth factors share binding sites in heparan sulphate. Biochem. J. 2005, 389 (Pt 1), 145–150. [Google Scholar] [CrossRef]
- Kreuger, J.; Spillmann, D.; Li, J.P.; Lindahl, U. Interactions between heparan sulfate and proteins: The concept of specificity. J. Cell Biol. 2006, 174, 323–327. [Google Scholar] [CrossRef]
- Lembo, D.; Donalisio, M.; Laine, C.; Cagno, V.; Civra, A.; Bianchini, E.P.; Zeghbib, N.; Bouchemal, K. Auto-associative heparin nanoassemblies: A biomimetic platform against the heparan sulfate-dependent viruses HSV-1, HSV-2, HPV-16 and RSV. Eur. J. Pharm. Biopharm. 2014, 88, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Connell, B.J.; Lortat-Jacob, H. Human immunodeficiency virus and heparan sulfate: From attachment to entry inhibition. Front. Immunol. 2013, 4, 385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Francesco, M.A.; Baronio, M.; Poiesi, C. HIV-1 p17 matrix protein interacts with heparan sulfate side chain of CD44v3, syndecan-2, and syndecan-4 proteoglycans expressed on human activated CD4+ T cells affecting tumor necrosis factor alpha and interleukin 2 production. J. Biol. Chem. 2011, 286, 19541–19548. [Google Scholar] [CrossRef] [PubMed]
- Imamura, J.; Suzuki, Y.; Gonda, K.; Roy, C.N.; Gatanaga, H.; Ohuchi, N.; Higuchi, H. Single particle tracking confirms that multivalent Tat protein transduction domain-induced heparan sulfate proteoglycan cross-linkage activates Rac1 for internalization. J. Biol. Chem. 2011, 286, 10581–10592. [Google Scholar] [CrossRef] [PubMed]
- Richards, K.F.; Bienkowska-Haba, M.; Dasgupta, J.; Chen, X.S.; Sapp, M. Multiple heparan sulfate binding site engagements are required for the infectious entry of human papillomavirus type 16. J. Virol. 2013, 87, 11426–11437. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.L.; Chung, C.S.; Heine, H.G.; Chang, W. Vaccinia virus envelope H3L protein binds to cell surface heparan sulfate and is important for intracellular mature virion morphogenesis and virus infection in vitro and in vivo. J. Virol. 2000, 74, 3353–3365. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.W.; Poh, C.L.; Sam, I.C.; Chan, Y.F. Enterovirus 71 uses cell surface heparan sulfate glycosaminoglycan as an attachment receptor. J. Virol. 2013, 87, 611–620. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Jiang, J.; Luo, G. Syndecan-1 serves as the major receptor for attachment of hepatitis C virus to the surfaces of hepatocytes. J. Virol. 2013, 87, 6866–6875. [Google Scholar] [CrossRef] [PubMed]
- Dalrymple, N.; Mackow, E.R. Productive dengue virus infection of human endothelial cells is directed by heparan sulfate-containing proteoglycan receptors. J. Virol. 2011, 85, 9478–9485. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.A.; Khomandiak, S.; Ashbrook, A.W.; Weller, R.; Heise, M.T.; Morrison, T.E.; Dermody, T.S. A single-amino-acid polymorphism in Chikungunya virus E2 glycoprotein influences glycosaminoglycan utilization. J. Virol. 2014, 88, 2385–2397. [Google Scholar] [CrossRef] [PubMed]
- Koski, A.; Karli, E.; Kipar, A.; Escutenaire, S.; Kanerva, A.; Hemminki, A. Mutation of the fiber shaft heparan sulphate binding site of a 5/3 chimeric adenovirus reduces liver tropism. PLoS One 2013, 8, e60032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, A.; Jinno-Oue, A.; Shimizu, N.; Hoque, A.; Mori, T.; Islam, S.; Nakatani, Y.; Shinagawa, M.; Hoshino, H. Entry of human T-cell leukemia virus type 1 is augmented by heparin sulfate proteoglycans bearing short heparin-like structures. J. Virol. 2012, 86, 2959–2969. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, K.; Zhao, Z.; Date, T.; Miyamoto, M.; Murayama, A.; Akazawa, D.; Tanabe, J.; Sone, S.; Wakita, T. The roles of CD81 and glycosaminoglycans in the adsorption and uptake of infectious HCV particles. J. Med. Virol. 2007, 79, 714–723. [Google Scholar] [CrossRef] [PubMed]
- Immergluck, L.C.; Domowicz, M.S.; Schwartz, N.B.; Herold, B.C. Viral and cellular requirements for entry of herpes simplex virus type 1 into primary neuronal cells. J. Gen. Virol. 1998, 79 (Pt 3), 549–559. [Google Scholar]
- Choudhary, S.; Burnham, L.; Thompson, J.M.; Shukla, D.; Tiwari, V. Role of Filopodia in HSV-1 Entry into Zebrafish 3-O-Sulfotransferase-3-Expressing Cells. Open Virol. J. 2013, 7, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, V.; Clement, C.; Xu, D.; Valyi-Nagy, T.; Yue, B.Y.; Liu, J.; Shukla, D. Role for 3-O-sulfated heparan sulfate as the receptor for herpes simplex virus type 1 entry into primary human corneal fibroblasts. J. Virol. 2006, 80, 8970–8980. [Google Scholar] [CrossRef] [PubMed]
- De Boer, S.M.; Kortekaas, J.; de Haan, C.A.; Rottier, P.J.; Moormann, R.J.; Bosch, B.J. Heparan sulfate facilitates Rift Valley fever virus entry into the cell. J. Virol. 2012, 86, 13767–13771. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, V.; Liu, J.; Valyi-Nagy, T.; Shukla, D. Anti-heparan sulfate peptides that block herpes simplex virus infection in vivo. J. Biol. Chem. 2011, 286, 25406–25415. [Google Scholar] [CrossRef] [PubMed]
- Raghuraman, A.; Tiwari, V.; Thakkar, J.N.; Gunnarsson, G.T.; Shukla, D.; Hindle, M.; Desai, U.R. Structural characterization of a serendipitously discovered bioactive macromolecule, lignin sulfate. Biomacromolecules 2005, 6, 2822–2832. [Google Scholar] [CrossRef] [PubMed]
- Raghuraman, A.; Tiwari, V.; Zhao, Q.; Shukla, D.; Debnath, A.K.; Desai, U.R. Viral inhibition studies on sulfated lignin, a chemically modified biopolymer and a potential mimic of heparan sulfate. Biomacromolecules 2007, 8, 1759–1763. [Google Scholar] [CrossRef] [PubMed]
- Thakkar, J.N.; Tiwari, V.; Desai, U.R. Nonsulfated, cinnamic acid-based lignins are potent antagonists of HSV-1 entry into cells. Biomacromolecules 2010, 11, 1412–1416. [Google Scholar] [CrossRef] [PubMed]
- Nyberg, K.; Ekblad, M.; Bergström, T.; Freeman, C.; Parish, C.R.; Ferro, V.; Trybala, E. The low molecular weight heparan sulfate-mimetic, PI-88, inhibits cell-to-cell spread of herpes simplex virus. Antivir. Res. 2004, 63, 15–24. [Google Scholar] [CrossRef]
- Damonte, E.B.; Matulewicz, M.C.; Cerezo, A.S. Sulfated seaweed polysaccharides as antiviral agents. Curr. Med. Chem. 2004, 11, 2399–2419. [Google Scholar] [CrossRef] [PubMed]
- Copeland, R.; Balasubramaniam, A.; Tiwari, V.; Zhang, F.; Bridges, A.; Linhardt, R.J.; Shukla, D.; Liu, J. Using a 3-O-sulfated heparin octasaccharide to inhibit the entry of herpes simplex virus type 1. Biochemistry 2008, 47, 5774–5783. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, A.; Remes Lenicov, F.; Sabatté, J.; Rodríguez Rodrígues, C.; Cabrini, M.; Jancic, C.; Raiden, S.; Donaldson, M.; Agustín Pasqualini, R., Jr.; Marin-Briggiler, C.; et al. Spermatozoa capture HIV-1 through heparan sulfate and efficiently transmit the virus to dendritic cells. J. Exp. Med. 2009, 206, 2717–2733. [Google Scholar] [CrossRef]
- De Witte, L.; Zoughlami, Y.; Aengeneyndt, B.; David, G.; van Kooyk, Y.; Gissmann, L.; Geijtenbeek, T.B. Binding of human papilloma virus L1 virus-like particles to dendritic cells is mediated through heparan sulfates and induces immune activation. Immunobiology 2007, 212, 679–691. [Google Scholar] [CrossRef] [PubMed]
- Shukla, D.; Liu, J.; Blaiklock, P.; Shworak, N.W.; Bai, X.; Esko, J.D.; Cohen, G.H.; Eisenberg, R.J.; Rosenberg, R.D.; Spear, P.G. A novel role for 3-O-sulfated heparan sulfate in herpes simplex virus 1 entry. Cell 1999, 99, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, R.; Yabe, T.; Hajmohammadi, S.; Rhodes, J.; McNeely, M.; Liu, J.; Lamperti, E.D.; Toselli, P.A.; Lech, M.; Spear, P.G.; et al. The principal neuronal gD-type 3-O-sulfotransferases and their products in central and peripheral nervous system tissues. Matrix Biol. 2007, 26, 442–455. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, C.D.; Kovacs, M.; Akhtar, J.; Valyi-Nagy, T.; Shukla, D. Expanding the role of 3-O sulfated heparan sulfate in herpes simplex virus type-1 entry. Virology 2010, 397, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, V.; ten Dam, G.B.; Yue, B.Y.; van Kuppevelt, T.H.; Shukla, D. Role of 3-O-sulfated heparan sulfate in virus-induced polykaryocyte formation. FEBS Lett. 2007, 581, 4468–4472. [Google Scholar] [CrossRef] [PubMed]
- Spear, P.G. Herpes simplex virus: Receptors and ligands for cell entry. Cell. Microbiol. 2004, 6, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Spear, P.G.; Longnecker, R. Herpesvirus entry: An update. J. Virol. 2003, 77, 10179–10185. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, J.; Shukla, D. Viral entry mechanisms: cellular and viral mediators of herpes simplex virus entry. FEBS J. 2009, 276, 7228–7236. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, X.; Chen, J.; Su, H.; Luo, Q.; Ye, J.; Tang, N.; Zhang, W.; Chen, W.; Ko, B.C.; et al. Heparin sulphate d-glucosaminyl 3-O-sulfotransferase 3B1 plays a role in HBV replication. Virology 2010, 406, 280–285. [Google Scholar] [CrossRef] [PubMed]
- Borst, E.M.; Ständker, L.; Wagner, K.; Schulz, T.F.; Forssmann, W.G.; Messerle, M. A peptide inhibitor of cytomegalovirus infection from human hemofiltrate. Antimicrob. Agents Chemother. 2013, 57, 4751–4760. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, J.; Shukla, D.; Tiwari, V. Members of 3-O-Sulfotransferases (3-OST) Family: A Valuable Tool from Zebrafish to Humans for Understanding Herpes Simplex Virus Entry. Open Virol. J. 2013, 7, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, S.; Darmani, N.A.; Thrush, G.R.; Dey, D.; Burnham, L.; Thompson, J.M.; Jones, K.; Tiwari, V. Zebrafish-encoded 3-O-sulfotransferase-3 isoform mediates herpes simplex virus type 1 entry and spread. Zebrafish 2010, 7, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Yakoub, A.M.; Rawal, N.; Maus, E.; Baldwin, J.; Shukla, D.; Tiwari, V. Comprehensive analysis of herpes simplex virus 1 (HSV-1) entry mediated by zebrafish 3-O-Sulfotransferase isoforms: implications for the development of a zebrafish model of HSV-1 infection. J. Virol. 2014, 88, 12915–12922. [Google Scholar] [CrossRef] [PubMed]
- Norose, K.; Yano, A.; Zhang, X.M.; Blankenhorn, E.; Heber-Katz, E. Mapping of genes involved in murine herpes simplex virus keratitis: identification of genes and their modifiers. J. Virol. 2002, 76, 3502–3510. [Google Scholar] [CrossRef] [PubMed]
- Zautner, A.E.; Jahn, B.; Hammerschmidt, E.; Wutzler, P.; Schmidtke, M. N- and 6-O-sulfated heparan sulfates mediate internalization of coxsackievirus B3 variant PD into CHO-K1 cells. J. Virol. 2006, 80, 629–636. [Google Scholar] [CrossRef]
- Makkonen, K.E.; Turkki, P.; Laakkonen, J.P.; Ylä-Herttuala, S.; Marjomäki, V.; Airenne, K.J. 6-O- and N-sulfated syndecan-1 promotes baculovirus binding and entry into Mammalian cells. J. Virol. 2013, 87, 11148–11159. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, F.; Yamada, S.; Taguwa, S.; Kataoka, C.; Naito, S.; Hama, Y.; Tani, H.; Matsuura, Y.; Sugahara, K. Specific interaction of the envelope glycoproteins E1 and E2 with liver heparan sulfate involved in the tissue tropismatic infection by hepatitis C virus. Glycoconj. J. 2012, 29, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Matos, P.M.; Andreu, D.; Santos, N.C.; Gutiérrez-Gallego, R. Structural requirements of glycosaminoglycans for their interaction with HIV-1 envelope glycoprotein gp120. Arch. Virol. 2014, 159, 555–560. [Google Scholar] [CrossRef] [PubMed]
- Rider, C.C.; Coombe, D.R.; Harrop, H.A.; Hounsell, E.F.; Bauer, C.; Feeney, J.; Mulloy, B.; Mahmood, N.; Hay, A.; Parish, C.R. Anti-HIV-1 activity of chemically modified heparins: Correlation between binding to the V3 loop of gp120 and inhibition of cellular HIV-1 infection in vitro. Biochemistry 1994, 33, 6974–6980. [Google Scholar] [CrossRef] [PubMed]
- Trybala, E.; Bergström, T.; Spillmann, D.; Svennerholm, B.; Olofsson, S.; Flynn, S.J.; Ryan, P. Mode of interaction between pseudorabies virus and heparan sulfate/heparin. Virology 1996, 218, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Trybala, E.; Liljeqvist, J.A.; Svennerholm, B.; Bergström, T. Herpes simplex virus types 1 and 2 differ in their interaction with heparan sulfate. J. Virol. 2000, 74, 9106–9114. [Google Scholar] [CrossRef] [PubMed]
- Seki, Y.; Mizukura, M.; Ichimiya, T.; Suda, Y.; Nishihara, S.; Masuda, M.; Takase-Yoden, S. O-sulfate groups of heparin are critical for inhibition of ecotropic murine leukemia virus infection by heparin. Virology 2012, 424, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Hallak, L.K.; Spillmann, D.; Collins, P.L.; Peeples, M.E. Glycosaminoglycan sulfation requirements for respiratory syncytial virus infection. J. Virol. 2000, 74, 10508–10513. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.J.; Akhtar, J.; Desai, P.; Shukla, D. A role for heparan sulfate in viral surfing. Biochem. Biophys. Res. Commun. 2010, 391, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Shworak, N.W.; Sinaÿ, P.; Schwartz, J.J.; Zhang, L.; Fritze, L.M.; Rosenberg, R.D. Expression of heparan sulfate d-glucosaminyl 3-O-sulfotransferase isoforms reveals novel substrate specificities. J. Biol. Chem. 1999, 274, 5185–5192. [Google Scholar] [CrossRef] [PubMed]
- Xia, G.; Chen, J.; Tiwari, V.; Ju, W.; Li, J.P.; Malmstrom, A.; Shukla, D.; Liu, J. Heparan sulfate 3-O-sulfotransferase isoform 5 generates both an antithrombin-binding site and an entry receptor for herpes simplex virus, type 1. J. Biol. Chem. 2002, 277, 37912–37919. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Tiwari, V.; Xia, G.; Clement, C.; Shukla, D.; Liu, J. Characterization of heparan sulphate 3-O-sulphotransferase isoform 6 and its role in assisting the entry of herpes simplex virus type 1. Biochem. J. 2005, 385 (Pt 2), 451–459. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, C.D.; Tiwari, V.; Oh, M.J.; Shukla, D. A role for heparan sulfate 3-O-sulfotransferase isoform 2 in herpes simplex virus type 1 entry and spread. Virology 2006, 346, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, V.; O’Donnell, C.D.; Oh, M.J.; Valyi-Nagy, T.; Shukla, D. A role for 3-O-sulfotransferase isoform-4 in assisting HSV-1 entry and spread. Biochem. Biophys. Res. Commun. 2005, 338, 930–937. [Google Scholar] [CrossRef] [PubMed]
- Shworak, N.W.; Liu, J.; Petros, L.M.; Zhang, L.; Kobayashi, M.; Copeland, N.G.; Jenkins, N.A.; Rosenberg, R.D. Multiple isoforms of heparan sulfate d-glucosaminyl 3-O-sulfotransferase. Isolation, characterization, and expression of human cdnas and identification of distinct genomic loci. J. Biol. Chem. 1999, 274, 5170–5184. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, H.; Yoshida, K.; Gotoh, M.; Sugioka, S.; Kikuchi, N.; Kwon, Y.D.; Tawada, A.; Maeyama, K.; Inaba, N.; Hiruma, T.; et al. Characterization of a heparan sulfate 3-O-sulfotransferase-5, an enzyme synthesizing a tetrasulfated disaccharide. J. Biol. Chem. 2003, 278, 26780–26787. [Google Scholar] [CrossRef] [PubMed]
- Yabe, T.; Hata, T.; He, J.; Maeda, N. Developmental and regional expression of heparan sulfate sulfotransferase genes in the mouse brain. Glycobiology 2005, 15, 982–993. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, B.; Li, J.P. Implications of heparan sulfate and heparanase in neuroinflammation. Matrix Biol. 2014, 35, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Yabe, T.; Maeda, N. Histochemical analysis of heparan sulfate 3-O-sulfotransferase expression in mouse brain. Methods Mol. Biol. 2015, 1229, 377–387. [Google Scholar] [PubMed]
- Park, P.J.; Chang, M.; Garg, N.; Zhu, J.; Chang, J.H.; Shukla, D. Corneal lymphangiogenesis in herpetic stromal keratitis. Surv. Ophthalmol. 2015, 60, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, K.; Biswas, P.S.; Kumaraguru, U.; Schoenberger, S.P.; Rouse, B.T. Protective and pathological roles of virus-specific and bystander CD8+ T cells in herpetic stromal keratitis. J. Immunol. 2004, 173, 7575–7583. [Google Scholar] [CrossRef] [PubMed]
- Shen, F.H.; Wang, S.W.; Yeh, T.M.; Tung, Y.Y.; Hsu, S.M.; Chen, S.H. Absence of CXCL10 aggravates herpes stromal keratitis with reduced primary neutrophil influx in mice. J. Virol. 2013, 87, 8502–8510. [Google Scholar] [CrossRef] [PubMed]
- Giménez, F.; Suryawanshi, A.; Rouse, B.T. Pathogenesis of herpes stromal keratitis—A focus on corneal neovascularization. Prog. Retin. Eye Res. 2013, 33, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rolinski, J.; Hus, I. Immunological aspects of acute and recurrent herpes simplex keratitis. J. Immunol. Res. 2014, 2014, 513560. [Google Scholar] [CrossRef] [PubMed]
- Van Wijk, X.M.; van Kuppevelt, T.H. Heparan sulfate in angiogenesis: A target for therapy. Angiogenesis 2014, 17, 443–462. [Google Scholar] [PubMed]
- Iozzo, R.V.; san Antonio, J.D. Heparan sulfate proteoglycans: Heavy hitters in the angiogenesis arena. J. Clin. Investig. 2001, 108, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Fuster, M.M.; Lawrence, R.; Esko, J.D. Heparan sulfate regulates VEGF165- and VEGF121-mediated vascular hyperpermeability. J. Biol. Chem. 2011, 286, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Robinson, C.J.; Mulloy, B.; Gallagher, J.T.; Stringer, S.E. VEGF165-binding sites within heparan sulfate encompass two highly sulfated domains and can be liberated by K5 lyase. J. Biol. Chem. 2006, 281, 1731–1740. [Google Scholar] [CrossRef] [PubMed]
- Maccarana, M.; Casu, B.; Lindahl, U. Minimal sequence in heparin/heparan sulfate required for binding of basic fibroblast growth factor. J. Biol. Chem. 1993, 268, 23898–23905. [Google Scholar] [PubMed]
- Guimond, S.; Maccarana, M.; Olwin, B.B.; Lindahl, U.; Rapraeger, A.C. Activating and inhibitory heparin sequences for FGF-2 (basic FGF). Distinct requirements for FGF-1, FGF-2, and FGF-4. J. Biol. Chem. 1993, 268, 23906–23914. [Google Scholar] [PubMed]
- Suryawanshi, A.; Veiga-Parga, T.; Reddy, P.B.; Rajasagi, N.K.; Rouse, B.T. IL-17A differentially regulates corneal vascular endothelial growth factor (VEGF)-A and soluble VEGF receptor 1 expression and promotes corneal angiogenesis after herpes simplex virus infection. J. Immunol. 2012, 188, 3434–3446. [Google Scholar] [CrossRef] [PubMed]
- Young, R.D.; Knupp, C.; Pinali, C.; Png, K.M.; Ralphs, J.R.; Bushby, A.J.; Starborg, T.; Kadler, K.E.; Quantock, A.J. Three-dimensional aspects of matrix assembly by cells in the developing cornea. Proc. Natl. Acad. Sci. USA 2014, 111, 687–692. [Google Scholar] [CrossRef] [PubMed]
- Clement, C.; Tiwari, V.; Scanlan, P.M.; Valyi-Nagy, T.; Yue, B.Y.; Shukla, D. A novel role for phagocytosis-like uptake in herpes simplex virus entry. J. Cell Biol. 2006, 174, 1009–1021. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, V.; Shukla, D. Nonprofessional phagocytosis can facilitate herpesvirus entry into ocular cells. Clin. Dev. Immunol. 2012, 2012, 651691. [Google Scholar] [CrossRef] [PubMed]
- Antoine, T.E.; Jones, K.S.; Dale, R.M.; Shukla, D.; Tiwari, V. Zebrafish: Modeling for herpes simplex virus infections. Zebrafish 2014, 11, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Meijer, A.H.; Spaink, H.P. Host-pathogen interactions made transparent with the zebrafish model. Curr. Drug Targets 2011, 12, 1000–1017. [Google Scholar] [CrossRef] [PubMed]
- Van der Sar, A.M.; Appelmelk, B.J.; Vandenbroucke-Grauls, C.M.; Bitter, W. A star with stripes: Zebrafish as an infection model. Trends Microbiol. 2004, 12, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, R.N.; van Eeden, F.J. The zebrafish as a model of vascular development and disease. Prog. Mol. Biol. Transl. Sci. 2014, 124, 93–122. [Google Scholar] [PubMed]
- Cadwallader, A.B.; Yost, H.J. Combinatorial expression patterns of heparan sulfate sulfotransferases in zebrafish: I. The 3-O-sulfotransferase family. Dev. Dyn. 2006, 235, 3423–3431. [Google Scholar] [CrossRef] [PubMed]
- Samson, S.C.; Ferrer, T.; Jou, C.J.; Sachse, F.B.; Shankaran, S.S.; Shaw, R.M.; Chi, N.C.; Tristani-Firouzi, M.; Yost, H.J. 3-OST-7 regulates BMP-dependent cardiac contraction. PLoS Biol. 2013, 11, e1001727. [Google Scholar] [CrossRef] [PubMed]
- Gabor, K.A.; Goody, M.F.; Mowel, W.K.; Breitbach, M.E.; Gratacap, R.L.; Witten, P.E.; Kim, C.H. Influenza A virus infection in zebrafish recapitulates mammalian infection and sensitivity to anti-influenza drug treatment. Dis. Model. Mech. 2014, 7, 1227–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heur, M.; Jiao, S.; Schindler, S.; Crump, J.G. Regenerative potential of the zebrafish corneal endothelium. Exp. Eye Res. 2013, 106, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Whiteford, J.R.; Ko, S.; Lee, W.; Couchman, J.R. Structural and cell adhesion properties of zebrafish syndecan-4 are shared with higher vertebrates. J. Biol. Chem. 2008, 283, 29322–29330. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, J.; Antoine, T.E.; Shukla, D.; Tiwari, V. Zebrafish encoded 3-O-sulfotransferase-2 generated heparan sulfate serves as a receptor during HSV-1 entry and spread. Biochem. Biophys. Res. Commun. 2013, 432, 672–676. [Google Scholar] [CrossRef] [PubMed]
- Antoine, T.E.; Yakoub, A.; Maus, E.; Shukla, D.; Tiwari, V. Zebrafish 3-O-sulfotransferase-4 generated heparan sulfate mediates HSV-1 entry and spread. PLoS One 2014, 9, e87302. [Google Scholar] [CrossRef] [PubMed]
- Burgos, J.S.; Ripoll-Gomez, J.; Alfaro, J.M.; Sastre, I.; Valdivieso, F. Zebrafish as a new model for herpes simplex virus type 1 infection. Zebrafish 2008, 5, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Salameh, S.; Sheth, U.; Shukla, D. Early events in herpes simplex virus lifecycle with implications for an infection of lifetime. Open Virol. J. 2012, 6, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Karasneh, G.A.; Shukla, D. Herpes simplex virus infects most cell types in vitro: Clues to its success. Virol. J. 2011, 8, p. 481. Available online: http://www.biomedcentral.com/content/pdf/1743-422X-8-481.pdf (accessed on 3 February 2015). [CrossRef] [PubMed]
- Esko, J.D.; Selleck, S.B. Order out of chaos: Assembly of ligand binding sites in heparan sulfate. Annu. Rev. Biochem. 2002, 71, 435–471. [Google Scholar] [CrossRef] [PubMed]
- Presto, J.; Thuveson, M.; Carlsson, P.; Busse, M.; Wilén, M.; Eriksson, I.; Kusche-Gullberg, M.; Kjellén, L. Heparan sulfate biosynthesis enzymes EXT1 and EXT2 affect NDST1 expression and heparan sulfate sulfation. Proc. Natl. Acad. Sci. USA 2008, 105, 4751–4756. [Google Scholar] [CrossRef] [PubMed]
- Ekblad, M.; Adamiak, B.; Bergstrom, T.; Johnstone, K.D.; Karoli, T.; Liu, L.; Ferro, V.; Trybala, E. A highly lipophilic sulfated tetrasaccharide glycoside related to muparfostat (PI-88) exhibits virucidal activity against herpes simplex virus. Antivir. Res. 2010, 86, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Ekblad, M.; Bergström, T.; Banwell, M.G.; Bonnet, M.; Renner, J.; Ferro, V.; Trybala, E. Anti-herpes simplex virus activities of two novel disulphated cyclitols. Antivir. Chem. Chemother. 2006, 17, 97–106. [Google Scholar] [PubMed]
- Ferro, V.; Liu, L.; Johnstone, K.D.; Wimmer, N.; Karoli, T.; Handley, P.; Rowley, J.; Dredge, K.; Li, C.P.; Hammond, E.; et al. Discovery of PG545: A highly potent and simultaneous inhibitor of angiogenesis, tumor growth, and metastasis. J. Med. Chem. 2012, 55, 3804–3813. [Google Scholar] [CrossRef] [PubMed]
- Said, J.; Trybala, E.; Andersson, E.; Johnstone, K.; Liu, L.; Wimmer, N.; Ferro, V.; Bergström, T. Lipophile-conjugated sulfated oligosaccharides as novel microbicides against HIV-1. Antivir. Res. 2010, 86, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Lundin, A.; Bergström, T.; Andrighetti-Fröhner, C.R.; Bendrioua, L.; Ferro, V.; Trybala, E. Potent anti-respiratory syncytial virus activity of a cholestanol-sulfated tetrasaccharide conjugate. Antivir. Res. 2012, 93, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, N.S.; Mancera, R.L. Heparin/heparan sulphate-based drugs. Drug Discov. Today 2010, 15, 1058–1069. [Google Scholar] [CrossRef] [PubMed]
- Khachigian, L.M.; Parish, C.R. Phosphomannopentaose sulfate (PI-88): Heparan sulfate mimetic with clinical potential in multiple vascular pathologies. Cardiovasc. Drug Rev. 2004, 22, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Knelson, E.H.; Nee, J.C.; Blobe, G.C. Heparan sulfate signaling in cancer. Trends Biochem. Sci. 2014, 39, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Ferro, V. Heparan sulfate inhibitors and their therapeutic implications in inflammatory illnesses. Expert Opin. Ther. Targets 2013, 17, 965–975. [Google Scholar] [CrossRef] [PubMed]
- Ferro, V.; Dredge, K.; Liu, L.; Hammond, E.; Bytheway, I.; Li, C.; Johnstone, K.; Karoli, T.; Davis, K.; Copeman, E.; et al. PI-88 and novel heparan sulfate mimetics inhibit angiogenesis. Semin. Thromb. Hemost. 2007, 33, 557–568. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, U.; Kjellén, L. Pathophysiology of heparan sulphate: many diseases, few drugs. J. Intern. Med. 2013, 273, 555–571. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.M.; Karasneh, G.A.; Jarding, M.J.; Tiwari, V.; Shukla, D. A 3-O-sulfated heparan sulfate binding peptide preferentially targets herpes simplex virus 2-infected cells. J. Virol. 2012, 86, 6434–6443. [Google Scholar] [CrossRef] [PubMed]
- Luganini, A.; Nicoletto, S.F.; Pizzuto, L.; Pirri, G.; Giuliani, A.; Landolfo, S.; Gribaudo, G. Inhibition of herpes simplex virus type 1 and type 2 infections by peptide-derivatized dendrimers. Antimicrob. Agents Chemother. 2011, 55, 3231–3239. [Google Scholar] [CrossRef] [PubMed]
- Connell, B.J.; Baleux, F.; Coic, Y.M.; Clayette, P.; Bonnaffé, D.; Lortat-Jacob, H. A synthetic heparan sulfate-mimetic peptide conjugated to a mini CD4 displays very high anti-HIV-1 activity independently of coreceptor usage. Chem. Biol. 2012, 19, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Baleux, F.; Loureiro-Morais, L.; Hersant, Y.; Clayette, P.; Arenzana-Seisdedos, F.; Bonnaffé, D.; Lortat-Jacob, H. A synthetic CD4-heparan sulfate glycoconjugate inhibits CCR5 and CXCR4 HIV-1 attachment and entry. Nat. Chem. Biol. 2009, 5, 743–748. [Google Scholar] [CrossRef] [PubMed]
- Bacsa, S.; Karasneh, G.; Dosa, S.; Liu, J.; Valyi-Nagy, T.; Shukla, D. Syndecan-1 and syndecan-2 play key roles in herpes simplex virus type-1 infection. J. Gen. Virol. 2011, 92 (Pt 4), 733–743. [Google Scholar] [CrossRef] [PubMed]
- Karasneh, G.A.; Ali, M.; Shukla, D. An important role for syndecan-1 in herpes simplex virus type-1 induced cell-to-cell fusion and virus spread. PLoS One 2011, 6, e25252. [Google Scholar] [CrossRef] [PubMed]
- Lortat-Jacob, H. The molecular basis and functional implications of chemokine interactions with heparan sulphate. Curr. Opin. Struct. Biol. 2009, 19, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Parish, C.R. The role of heparan sulphate in inflammation. Nat. Rev. Immunol. 2006, 6, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Götte, M. Syndecans in inflammation. FASEB J. 2003, 17, 575–591. [Google Scholar] [CrossRef] [PubMed]
- Webb, L.M.; Ehrengruber, M.U.; Clark-Lewis, I.; Baggiolini, M.; Rot, A. Binding to heparan sulfate or heparin enhances neutrophil responses to interleukin 8. Proc. Natl. Acad. Sci. USA 1993, 90, 7158–7162. [Google Scholar] [CrossRef] [PubMed]
- Duan, R.; Remeijer, L.; van Dun, J.M.; Osterhaus, A.D.; Verjans, G.M. Granulocyte macrophage colony-stimulating factor expression in human herpetic stromal keratitis: Implications for the role of neutrophils in HSK. Investig. Ophthalmol. Vis. Sci. 2007, 48, 277–284. [Google Scholar] [CrossRef]
- Turnbull, J.E.; Miller, R.L.; Ahmed, Y.; Puvirajesinghe, T.M.; Guimond, S.E. Glycomics profiling of heparan sulfate structure and activity. Methods Enzymol. 2010, 480, 65–85. [Google Scholar] [PubMed]
- Taylor, K.R.; Gallo, R.L. Glycosaminoglycans and their proteoglycans: host-associated molecular patterns for initiation and modulation of inflammation. FASEB J. 2006, 20, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Stringer, S.E. The role of heparan sulphate proteoglycans in angiogenesis. Biochem. Soc. Trans. 2006, 34 (Pt 3), 451–453. [Google Scholar] [CrossRef] [PubMed]
- Adamiak, B.; Ekblad, M.; Bergström, T.; Ferro, V.; Trybala, E. Herpes simplex virus type 2 glycoprotein G is targeted by the sulfated oligo- and polysaccharide inhibitors of virus attachment to cells. J. Virol. 2007, 81, 13424–13434. [Google Scholar] [CrossRef] [PubMed]
- Tuve, S.; Wang, H.; Jacobs, J.D.; Yumul, R.C.; Smith, D.F.; Lieber, A. Role of cellular heparan sulfate proteoglycans in infection of human adenovirus serotype 3 and 35. PLoS Pathog. 2008, 4, e1000189. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.Q.; Wang, X.Q.; Guo, Y.N.; Wu, J.; Li, S.; Yu, P.; Wang, Z. HS N-sulfation and iduronic acids play an important role in the infection of respiratory syncytial virus in vitro. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 1864–1868. [Google Scholar] [PubMed]
- Schowalter, R.M.; Pastrana, D.V.; Buck, C.B. Glycosaminoglycans and sialylated glycans sequentially facilitate Merkel cell polyomavirus infectious entry. PLoS Pathog. 2011, 7, e1002161. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Thorp, S.C. Cell surface heparan sulfate and its roles in assisting viral infections. Med. Res. Rev. 2002, 22, 1–25. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tiwari, V.; Tarbutton, M.S.; Shukla, D. Diversity of Heparan Sulfate and HSV Entry: Basic Understanding and Treatment Strategies. Molecules 2015, 20, 2707-2727. https://doi.org/10.3390/molecules20022707
Tiwari V, Tarbutton MS, Shukla D. Diversity of Heparan Sulfate and HSV Entry: Basic Understanding and Treatment Strategies. Molecules. 2015; 20(2):2707-2727. https://doi.org/10.3390/molecules20022707
Chicago/Turabian StyleTiwari, Vaibhav, Morgan S. Tarbutton, and Deepak Shukla. 2015. "Diversity of Heparan Sulfate and HSV Entry: Basic Understanding and Treatment Strategies" Molecules 20, no. 2: 2707-2727. https://doi.org/10.3390/molecules20022707
APA StyleTiwari, V., Tarbutton, M. S., & Shukla, D. (2015). Diversity of Heparan Sulfate and HSV Entry: Basic Understanding and Treatment Strategies. Molecules, 20(2), 2707-2727. https://doi.org/10.3390/molecules20022707