
Nitric Oxide Plays a Central Role in Water Stress-Induced Tanshinone Production in Salvia miltiorrhiza Hairy Roots

Abstract
:1. Introduction
2. Results
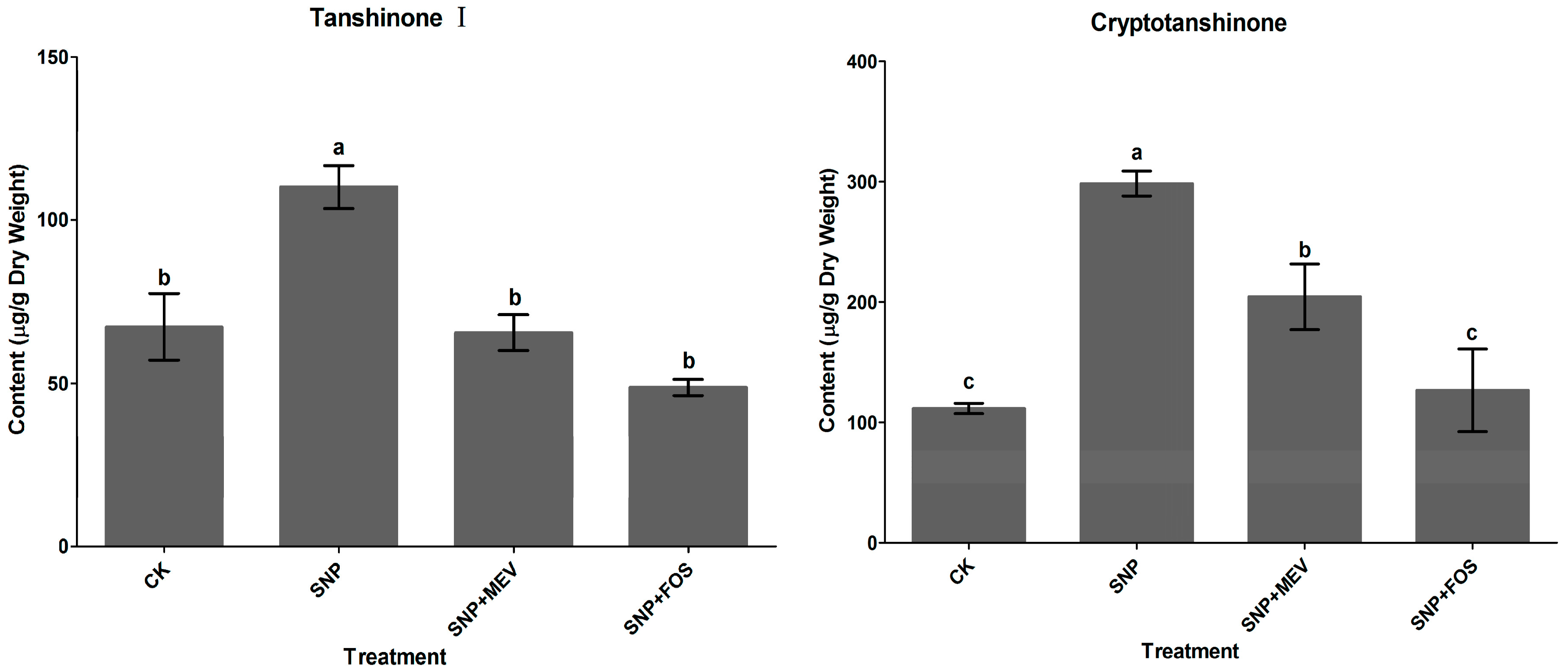
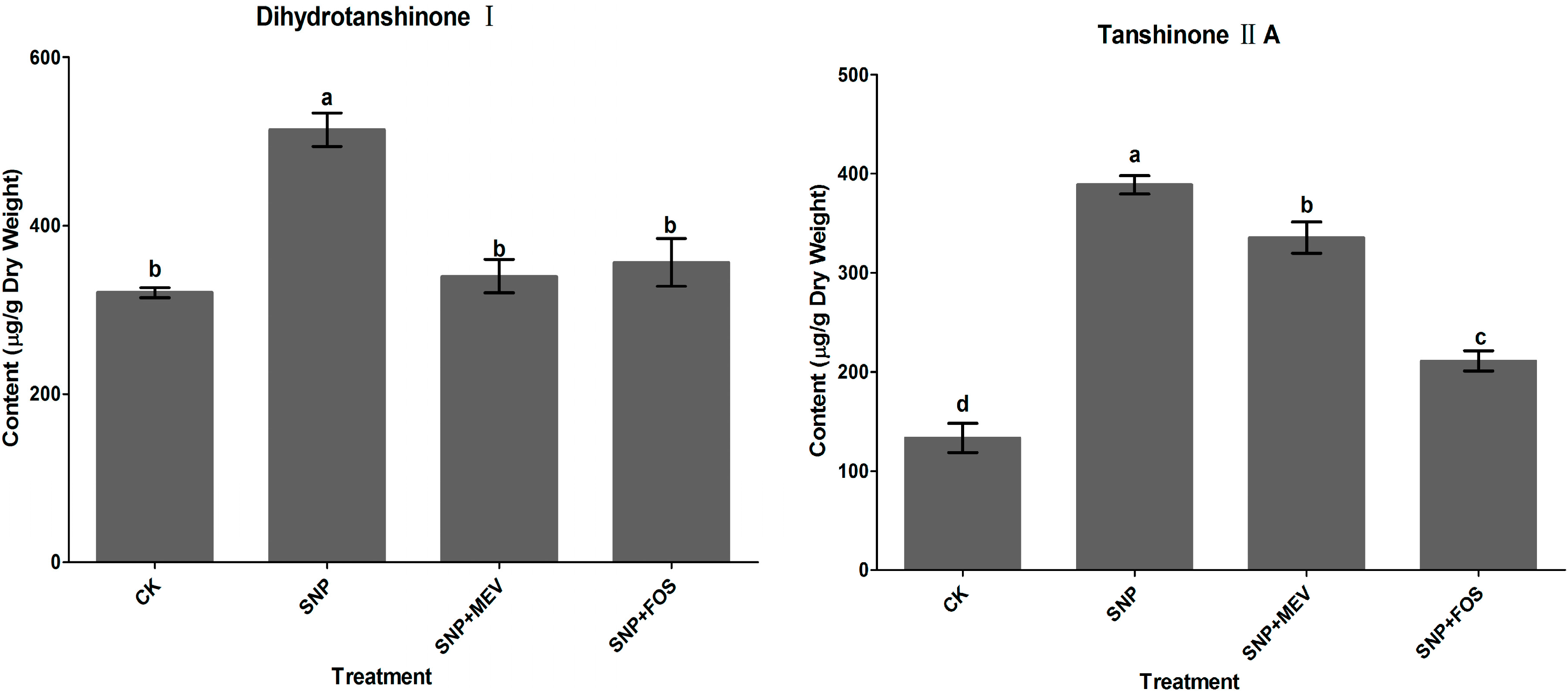
2.1. Effects of SNP on Tanshinone Production in S. miltiorrhiza Hairy Roots


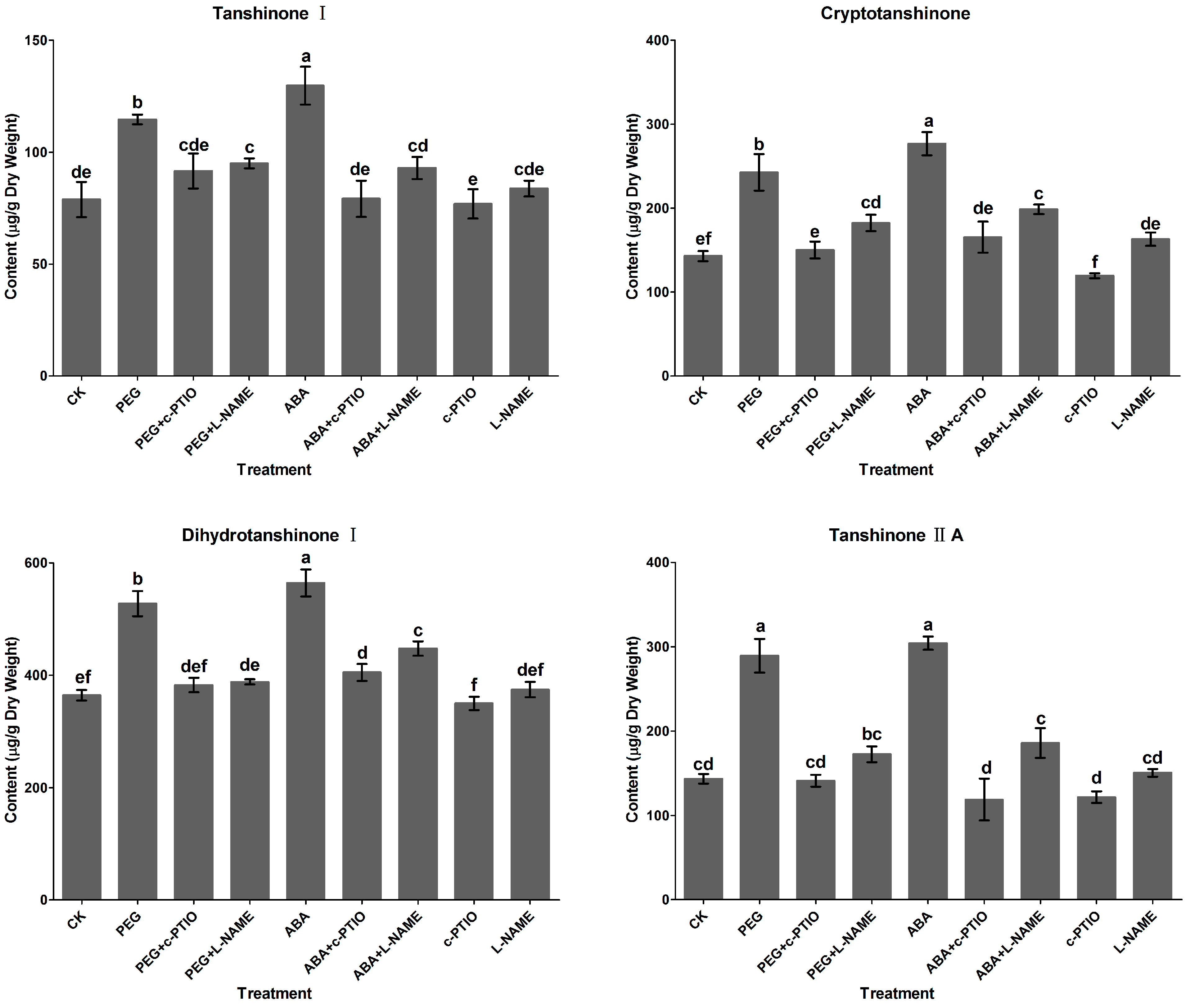
2.2. Effects of c-PTIO and l-NAME on Water Stress-Induced Tanshinone Production

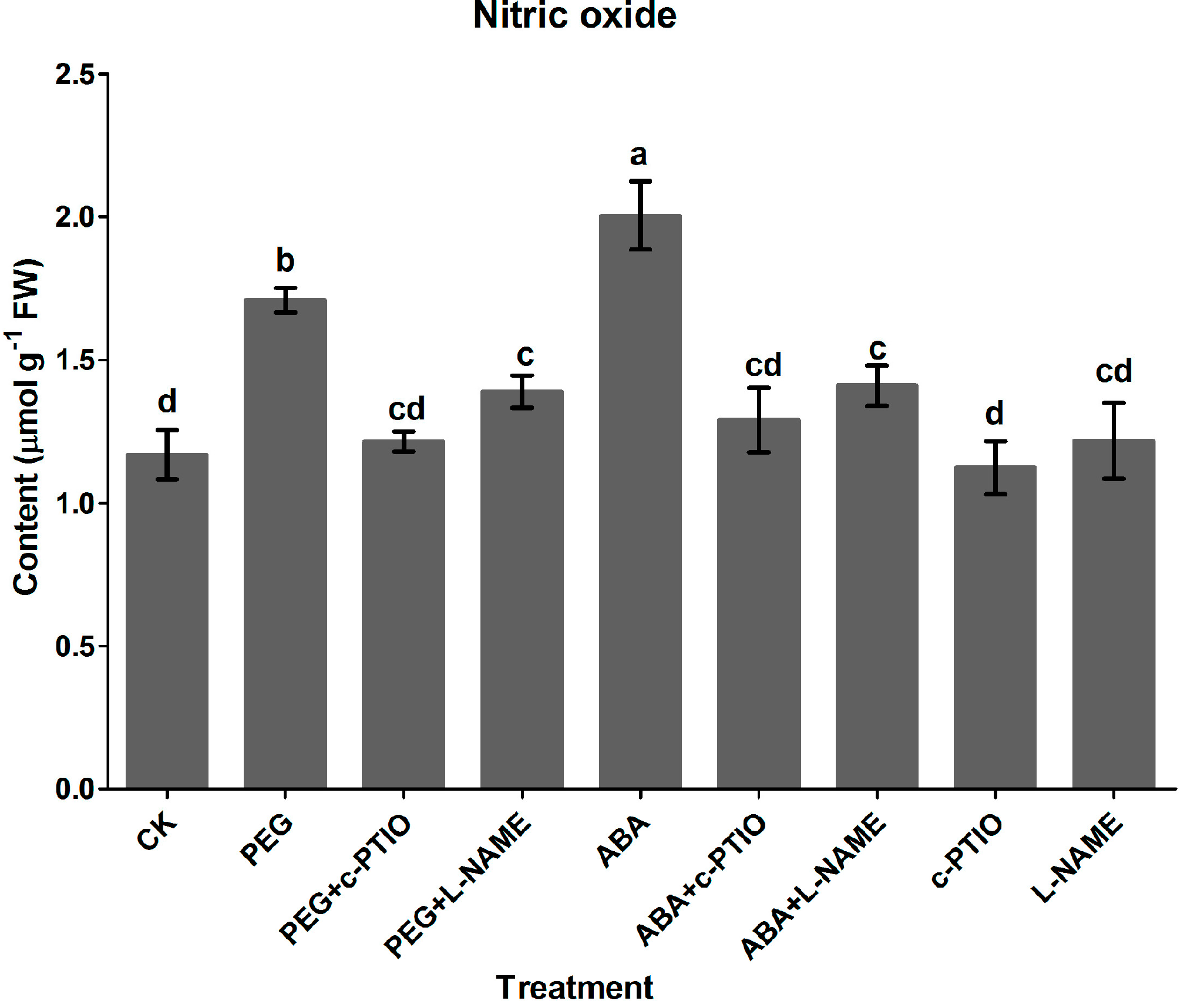
2.3. Generation of Nitric Oxide Induced by Water Stress


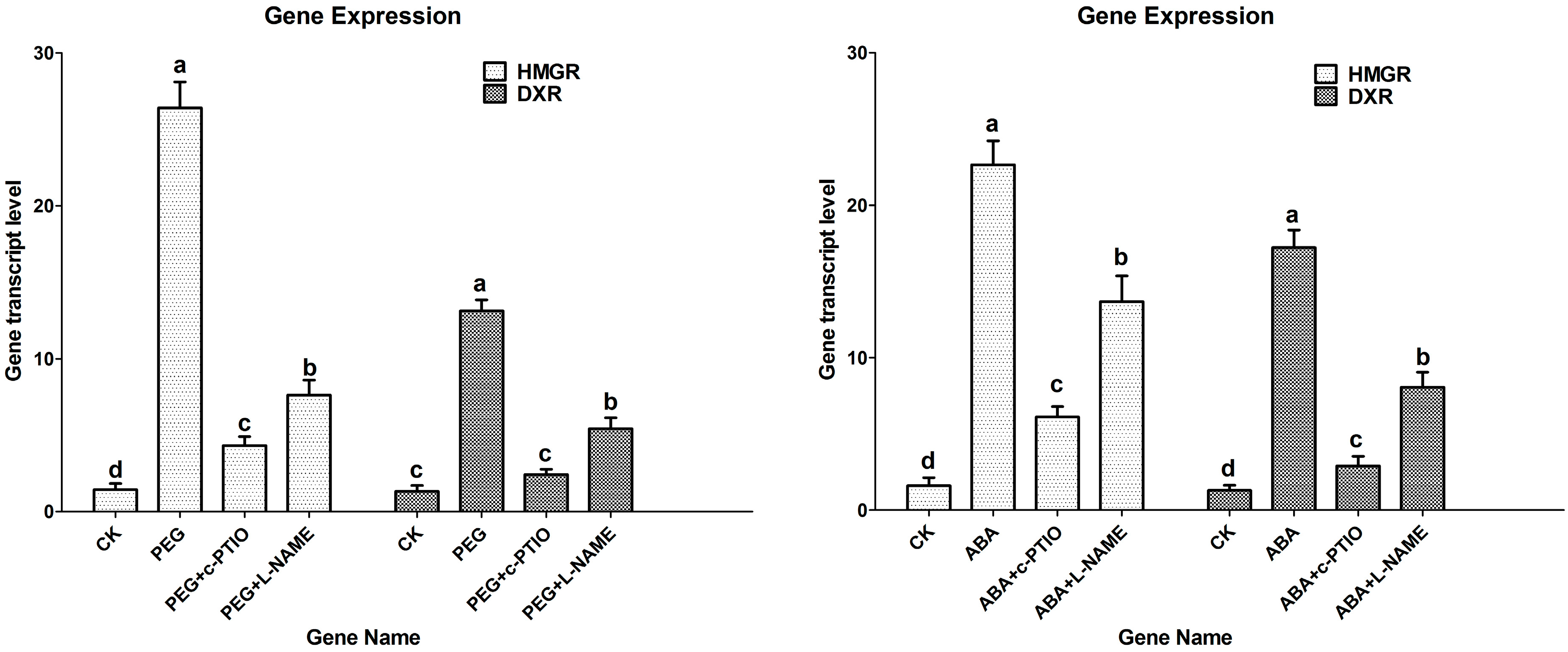
2.4. Effects of Water Stress on Expressions of HMGR and DXR
3. Discussion
4. Experimental Section
4.1. Hairy Root Culture and Treatment
4.2. Determination of Nitric Oxide
4.3. Extraction and HPLC Analysis of Tanshinone
4.4. Quantitative Real Time PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence | Gene Bank ID |
|---|---|---|
| HMGR-F | 5'-GCAACATCGTCTCCGCCGTCTACA-3' | FJ747636 |
| HMGR-R | 5'-GATGGTGGCCAGCAGCCTGGAGTT-3' | FJ747636 |
| DXR-F | 5'-CATGCGTTTGCCTATTCTGTAC-3' | DQ991431 |
| DXR-R | 5'-ACTAAGAACTCCGGTCATGGTG-3' | DQ991431 |
| β-actin-F | 5'-AGGAACCACCGATCCAGACA-3' | DQ243702.1 |
| β-actin-R | 5'-GGTGCCCTGAGGTCCTGTT-3' | DQ243702.1 |
4.5. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Abbreviations
| ABA | abscisic acid |
| c-PTIO | 2-(4-Carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide |
| DAD | Photodiode Array Detector |
| DXR | 1-deoxy-d-xylulose 5-phosphate reductoisomerase |
| DXS | 1-deoxy-d-xylulose-5-phosphatesynthase |
| FOS | fosmidomycin |
| HMGR | 3-hydroxy-3-methylglutaryl coenzyme A reductase |
| HPLC | high performance liquid chromatography |
| l-NAME | Nω-nitro-l-arginine methyl ester |
| MVA | mevalonate |
| MEP | 2-C-methyl-d-erythritol-4-phosphate |
| MEV | mevinolin |
| NO | Nitric oxide |
| PEG | polyethylene glycol |
| SNP | sodium nitroprusside |
Conflicts of Interest
References
- Dixon, R.A. Natural products and plant disease resistance. Nature 2001, 411, 843–847. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.B.; Liang, Z.S.; Han, R.L. Saikosaponin accumulation and antioxidative protection in drought-stressed Bupleurum chinense DC. plants. Environ. Exp. Bot. 2009, 66, 326–333. [Google Scholar] [CrossRef]
- Zhang, J.; Jia, W.; Yang, J.; Ismail, A.M. Role of ABA in integrating plant responses to drought. Field Crops Res. 2006, 97, 111–119. [Google Scholar] [CrossRef]
- Mansouri, H.; Asrar, Z.; Szopa, J. Effects of ABA on primary terpenoids and Δ9-tetrahydrocannabinol in Cannabis sativa L. at flowering stage. Plant Growth Regul. 2009, 58, 269–277. [Google Scholar] [CrossRef]
- Song, L.L.; Ding, W.; Shen, J.; Zhang, Z.G.; Bi, Y.R.; Zhang, L.X. Nitric oxide mediates abscisic acid induced thermotolerance in the calluses from two ecotypes of reed under heat stress. Plant Sci. 2008, 175, 826–832. [Google Scholar] [CrossRef]
- Zhang, A.Y.; Zhang, J.; Zhang, J.H.; Ye, N.H.; Zhang, H.; Tan, M.P.; Jiang, M.Y. Nitric Oxide Mediates Brassinosteroid-Induced ABA Biosynthesis Involved in Oxidative Stress Tolerance in Maize Leaves. Plant Cell Physiol. 2011, 52, 181–192. [Google Scholar] [CrossRef]
- Huang, A.X.; She, X.P.; Cao, B.; Zhang, B.; Mu, J.; Zhang, S.J. Nitric oxide, actin reorganization and vacuoles change are involved in PEG 6000-induced stomatal closure in Vicia faba. Physiol. Plant. 2009, 136, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Zheng, L.P.; Wu, J.Y.; Tan, R.X. Involvement of nitric oxide in oxidative burst, phenylalanine ammonia-lyase activation and Taxol production induced by low-energy ultrasound in Taxus yunnanensis cell suspension cultures. Nitric Oxide 2006, 15, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Neill, S.J.; Desikan, R.; Clarke, A.; Hancock, J.T. Nitric Oxide Is a Novel Component of Abscisic Acid Signaling in Stomatal Guard Cells. Plant Physiol. 2002, 128, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.M.; Zuo, Z.; Chow, M.S.S. Danshen: An overview of its chemistry, pharmacology, pharmacokinetics, and clinical use. J. Clin. Pharmacol. 2005, 45, 1345–1359. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Du, X.; Liang, X.; Han, R.; Liang, Z.; Liu, Y.; Liu, F.; Zhao, J. Different Roles of the Mevalonate and Methylerythritol Phosphate Pathways in Cell Growth and Tanshinone Production of Salvia miltiorrhiza Hairy Roots. PLoS ONE 2012, 7, e46797. [Google Scholar] [CrossRef] [PubMed]
- Bach, T.J.; Lichtenthaler, H.K. Mevinolin: A highly specific inhibitor of microsomal 3-hydroxy-3-methylglutaryl-coenzyme A reductase of radish plants. Z. Naturforschung 1982, 37, 46–50. [Google Scholar]
- Kai, G.; Xu, H.; Zhou, C.; Liao, P.; Xiao, J.; Luo, X.; You, L.; Zhang, L. Metabolic engineering tanshinone biosynthetic pathway in Salvia miltiorrhiza hairy root cultures. Metab. Eng. 2011, 13, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.Y.; Wang, X.D.; Wang, D.H.; Zou, Z.R.; Liang, Z.S. Effect of drought stress on growth and accumulation of active constituents in Salvia miltiorrhiza Bunge. Ind. Crops Prod. 2011, 33, 84–88. [Google Scholar] [CrossRef]
- Yang, D.F.; Ma, P.D.; Liang, X.; Wei, Z.; Liang, Z.S.; Liu, Y.; Liu, F.H. PEG and ABA trigger methyl jasmonate accumulation to induce the MEP pathway and increase tanshinone production in Salvia miltiorrhiza hairy roots. Physiol. Plant. 2012, 146, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.S.; Yang, D.F.; Liang, X.; Zhang, Y.J.; Liu, Y.; Liu, F.H. Roles of reactive oxygen species in methyl jasmonate and nitric oxide-induced tanshinone production in Salvia miltiorrhiza hairy roots. Plant. Cell Rep. 2012, 31, 873–883. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.B.; Cui, G.H.; Zhou, S.F.; Zhang, X.A.; Huang, L.Q. Cloning and characterization of a novel 3-hydroxy-3-methylglutaryl coenzyme A reductase gene from Salvia miltiorrhiza involved in diterpenoid tanshinone accumulation. J. Plant. Physiol. 2011, 168, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Zhang, L.; Wang, J.; Liao, P.; Zhang, Y.; Zhang, R.; Kai, G. Molecular characterization and expression of 1-deoxy-d-xylulose 5-phosphate reductoisomerase (DXR) gene from Salvia miltiorrhiza. Acta Physiol. Plant. 2009, 31, 1015–1022. [Google Scholar] [CrossRef]
- Wu, S.J.; Shi, M.; Wu, J.Y. Cloning and characterization of the 1-deoxy-d-xylulose 5-phosphate reductoisomerase gene for diterpenoid tanshinone biosynthesis in Salvia miltiorrhiza (Chinese sage) hairy roots. Biotechnol. Appl. Biochem. 2009, 52, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.G.; Tian, Q.T.; Zhang, W.H. Nitric oxide synthase-dependent nitric oxide production is associated with salt tolerance in Arabidopsis. Plant. Physiol. 2007, 144, 206–217. [Google Scholar] [CrossRef] [PubMed]
- Foissner, I.; Wendehenne, D.; Langebartels, C.; Durner, J. In vivo imaging of an elicitor-induced nitric oxide burst in tobacco. Plant. J. 2000, 23, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Wu, J.Y. Nitric oxide is involved in methyl jasmonate-induced defense responses and secondary metabolism activities of Taxus cells. Plant. Cell Physiol. 2005, 46, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.J.; Dong, J.F. Nitric oxide stimulates indole alkaloid production in Catharanthus roseus cell suspension cultures through a protein kinase-dependent signal pathway. Enzym. Microb. Technol. 2005, 37, 49–53. [Google Scholar] [CrossRef]
- Zhao, Z.G.; Chen, G.C.; Zhang, C.L. Interaction between reactive oxygen species and nitric oxide in drought-induced abscisic acid synthesis in root tips of wheat seedlings. Aust. J. Plant. Physiol. 2001, 28, 1055–1061. [Google Scholar]
- Yang, D.; Sheng, D.; Duan, Q.; Liang, X.; Liang, Z.; Liu, Y. PEG and ABA Trigger the Burst of Reactive Oxygen Species to Increase Tanshinone Production in Salvia miltiorrhiza Hairy Roots. J. Plant. Growth Regul. 2012, 31, 579–587. [Google Scholar] [CrossRef]
- Hagen, C.; Grunewald, K. Fosmidomycin as an inhibitor of the non-mevalonate terpenoid pathway depresses synthesis of secondary carotenoids in flagellates of the green alga Haematococcus pluvialis. J. Appl. Bot. 2000, 74, 137–140. [Google Scholar]
- Samples Availability: Samples of the compounds are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, X.; Zhang, C.; Guo, W.; Jin, W.; Liang, Z.; Yan, X.; Guo, Z.; Liu, Y.; Yang, D. Nitric Oxide Plays a Central Role in Water Stress-Induced Tanshinone Production in Salvia miltiorrhiza Hairy Roots. Molecules 2015, 20, 7574-7585. https://doi.org/10.3390/molecules20057574
Du X, Zhang C, Guo W, Jin W, Liang Z, Yan X, Guo Z, Liu Y, Yang D. Nitric Oxide Plays a Central Role in Water Stress-Induced Tanshinone Production in Salvia miltiorrhiza Hairy Roots. Molecules. 2015; 20(5):7574-7585. https://doi.org/10.3390/molecules20057574
Chicago/Turabian StyleDu, Xuhong, Chenlu Zhang, Wanli Guo, Weibo Jin, Zongsuo Liang, Xijun Yan, Zhixin Guo, Yan Liu, and Dongfeng Yang. 2015. "Nitric Oxide Plays a Central Role in Water Stress-Induced Tanshinone Production in Salvia miltiorrhiza Hairy Roots" Molecules 20, no. 5: 7574-7585. https://doi.org/10.3390/molecules20057574
APA StyleDu, X., Zhang, C., Guo, W., Jin, W., Liang, Z., Yan, X., Guo, Z., Liu, Y., & Yang, D. (2015). Nitric Oxide Plays a Central Role in Water Stress-Induced Tanshinone Production in Salvia miltiorrhiza Hairy Roots. Molecules, 20(5), 7574-7585. https://doi.org/10.3390/molecules20057574