
Transport of Twelve Coumarins from Angelicae Pubescentis Radix across a MDCK-pHaMDR Cell Monolayer—An in Vitro Model for Blood-Brain Barrier Permeability

Abstract
:1. Introduction

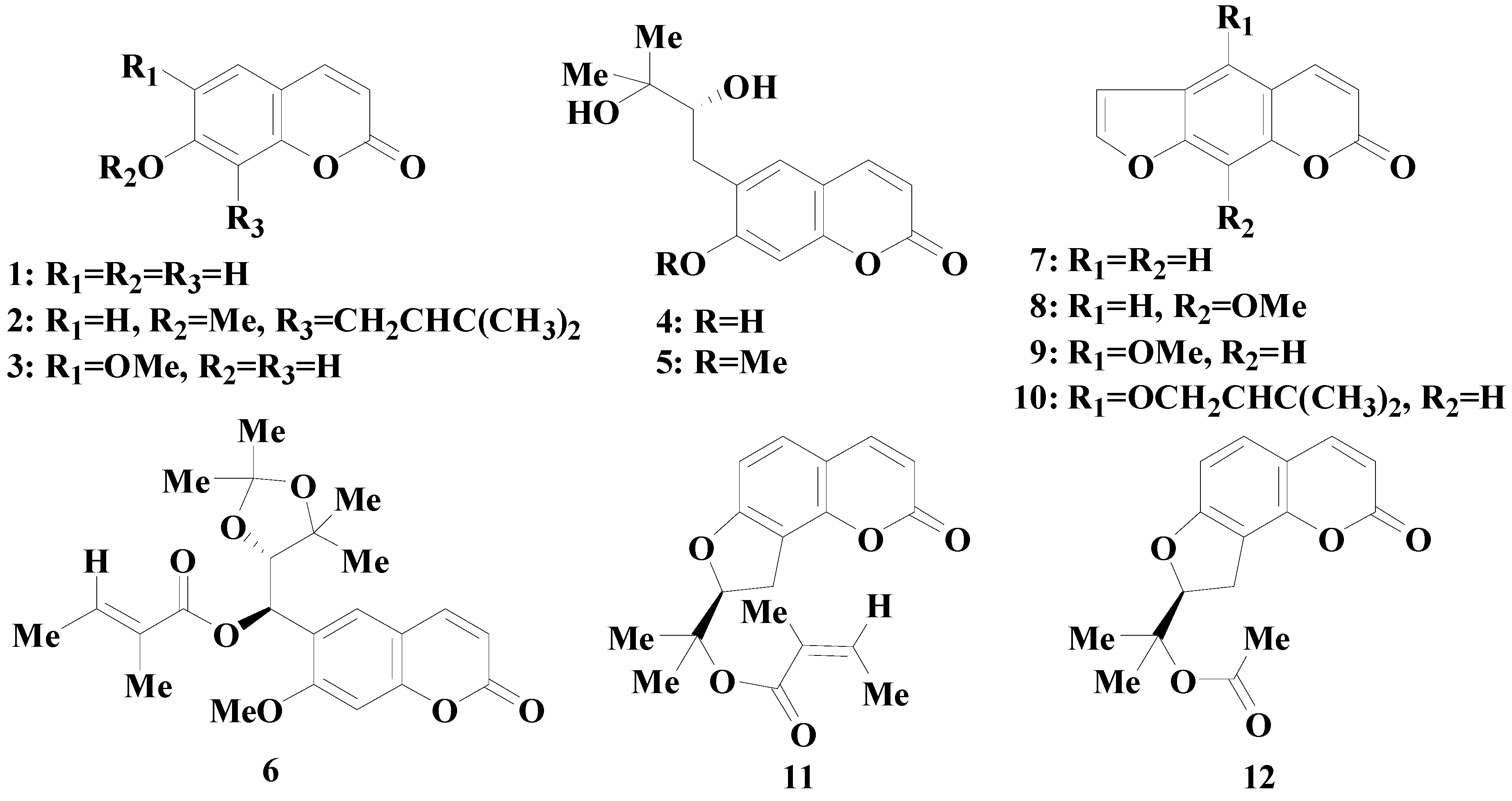
2. Results and Discussion
2.1. Validation of the MDCK-pHaMDR Cell System

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Papp AP-BL (×10−6 cm/s) | Papp BL-AP (×10−6 cm/s) | Papp BL-AP/Papp AP-BL | LogD (pH 7.4) | TEER (Ω·cm2) |
|---|---|---|---|---|---|
| 1 | 35.86 ± 2.28 | 32.79 ± 1.25 | 0.91 | 1.44 | 1321 ± 129 |
| 2 | 23.79 ± 1.44 | 20.37 ± 3.04 | 0.86 | 4.08 | 1548 ± 104 |
| 3 | 37.10 ± 1.97 | 27.38 ± 3.51 | 0.96 | 1.65 | 1586 ± 194 |
| 4 | 9.91 ± 0.21 | 18.97 ± 0.38 | 1.91 | 0.63 | 1595 ± 93 |
| 5 | 22.36 ± 4.02 | 26.66 ± 2.94 | 1.19 | 1.09 | 1664 ± 190 |
| 6 | 20.85 ± 1.32 | 18.73 ± 1.02 | 0.90 | 3.40 | 1347 ± 110 |
| 7 | 27.34 ± 2.35 | 26.75 ± 2.75 | 0.98 | 2.08 | 1549 ± 82 |
| 8 | 26.21 ± 3.54 | 23.34 ± 1.67 | 0.89 | 2.17 | 1298 ± 161 |
| 9 | 43.62 ± 2.13 | 37.38 ± 5.81 | 0.86 | 2.17 | 1595 ± 173 |
| 10 | 7.29 ± 0.17 | 3.73 ± 0.33 | 0.51 | 3.7 | 1615 ± 136 |
| 11 | 10.42 ± 0.95 | 7.12 ± 0.12 | 0.68 | 4.28 | 1429 ± 152 |
| 12 | 38.34 ± 1.72 | 33.74 ± 6.35 | 0.88 | 2.54 | 1463 ± 119 |
| caffeine (10 µM) | 43.58 ± 4.86 | 1635 ± 53 | |||
| Atenolol (200 µM) | 0.76 ± 0.16 | 1573 ± 91 |
| Compound | Papp (×10−7 cm/s) | Papp BL-AP/Papp AP-BL | Intracellular Accumulation Fluorescence Intensity | |
|---|---|---|---|---|
| AP-BL | BL-AP | |||
| Rh123 | 11.81 ± 0.56 | 52.74 ± 2.09 | 4.47 ± 0.24 | 34.51 ± 1.18 |
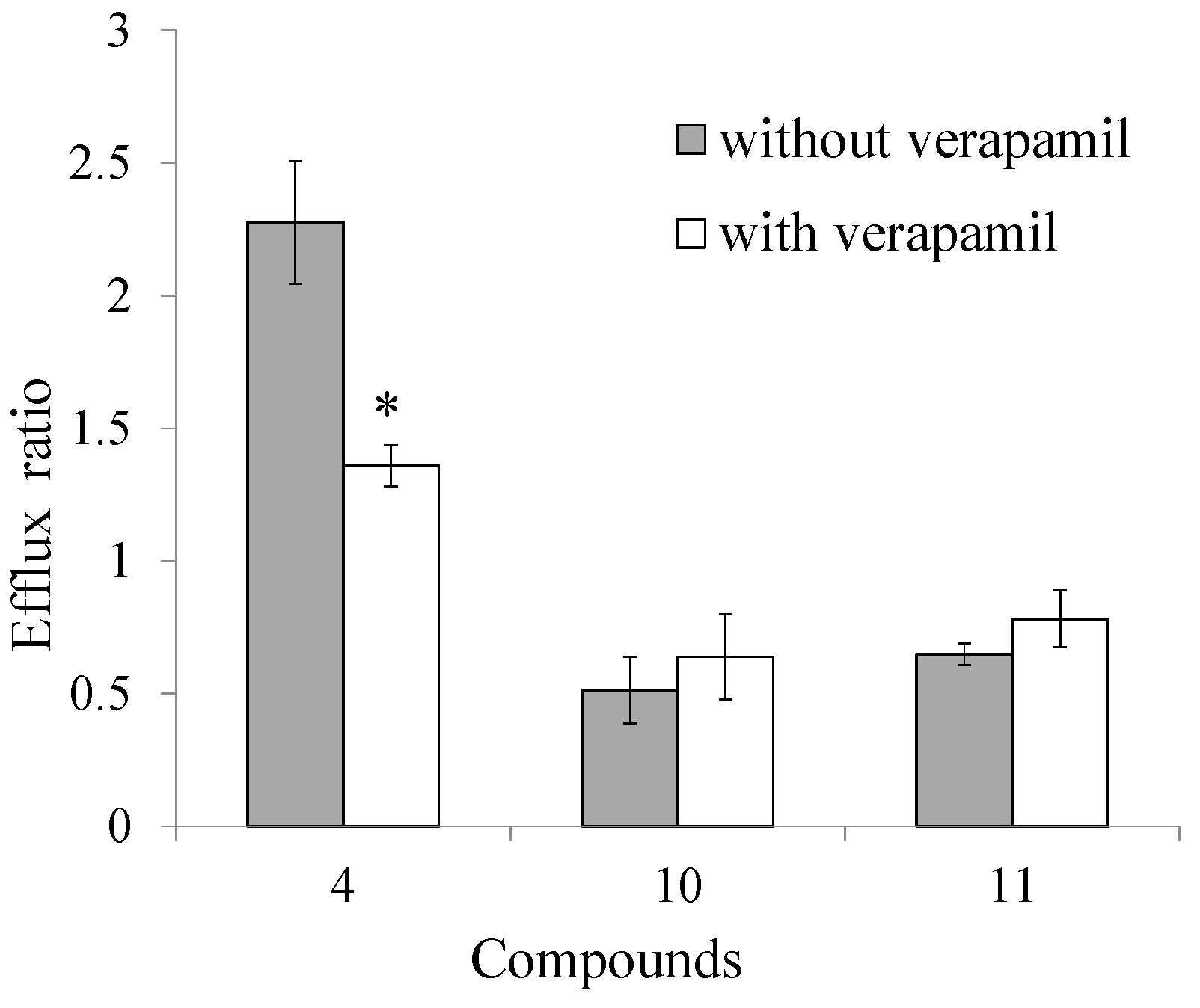
| Rh123 + 100 µM verapamil | 5.36 ± 0.41 | 8.51 ± 1.43 | 1.58 ± 0.208 * | 153.64 ± 3.28 * |
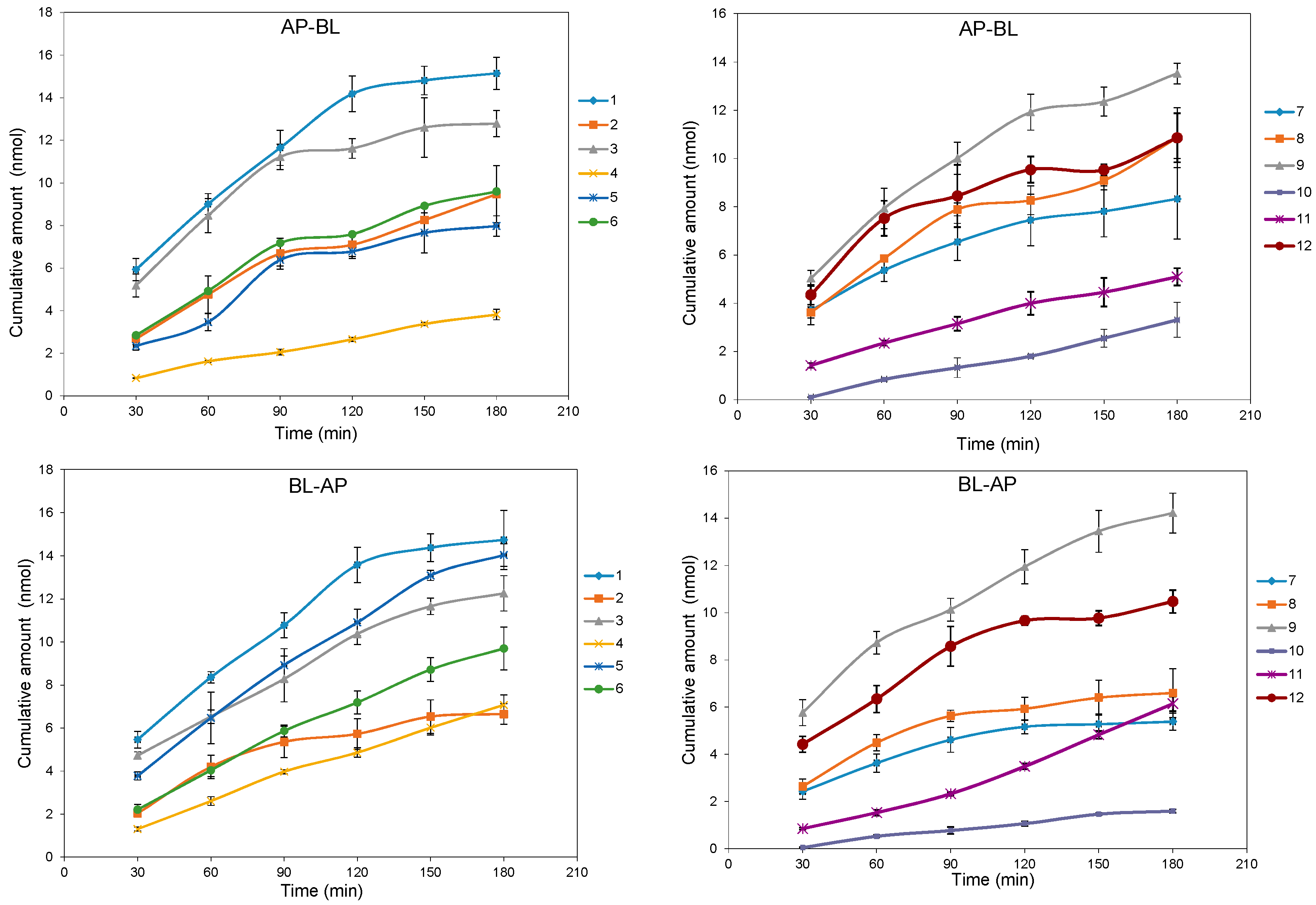
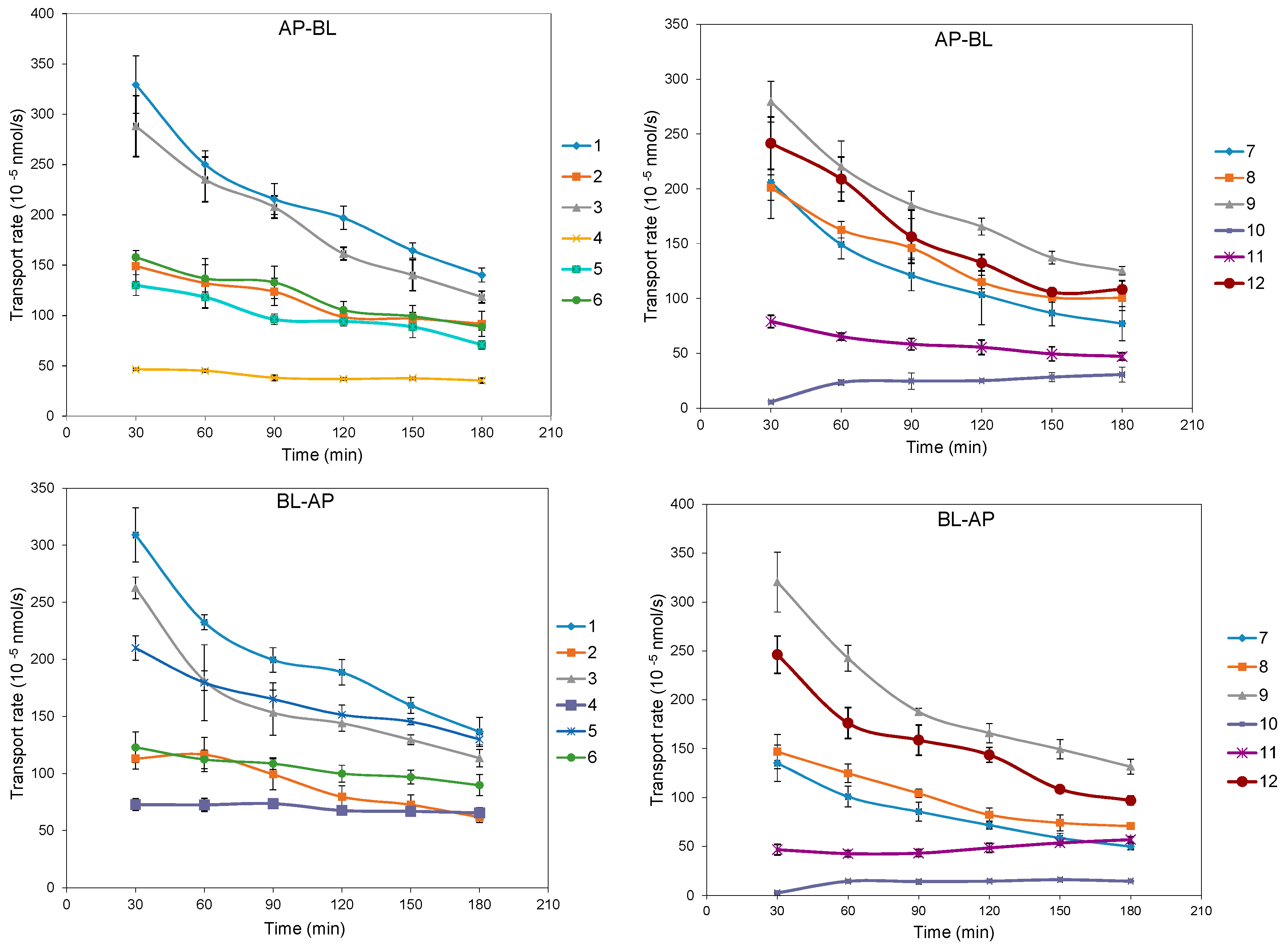
2.2. Bidirectional Transport Determination
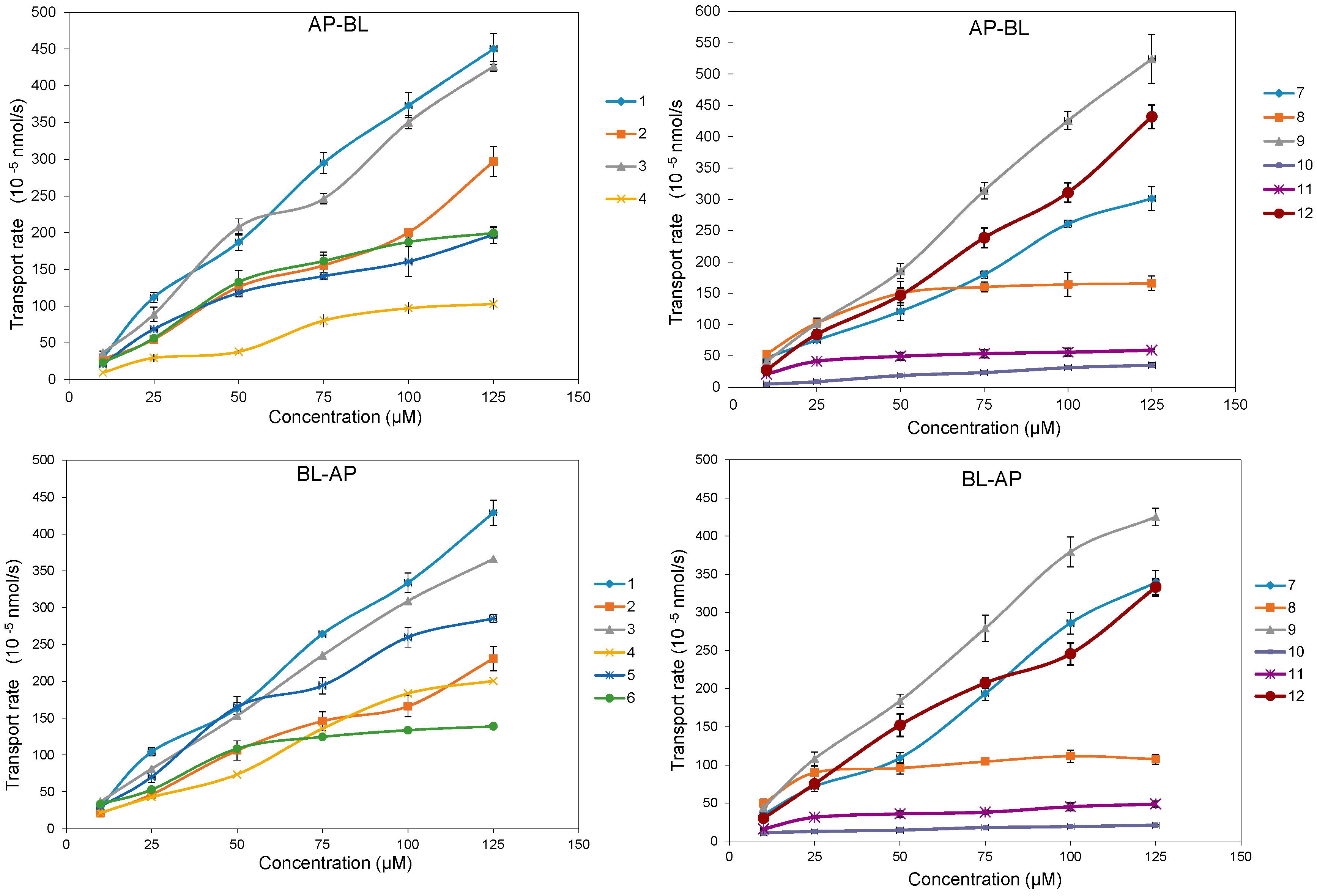
2.3. Time- and Concentration-Dependent Permeation



2.4. Transport Inhibition by Verapamil

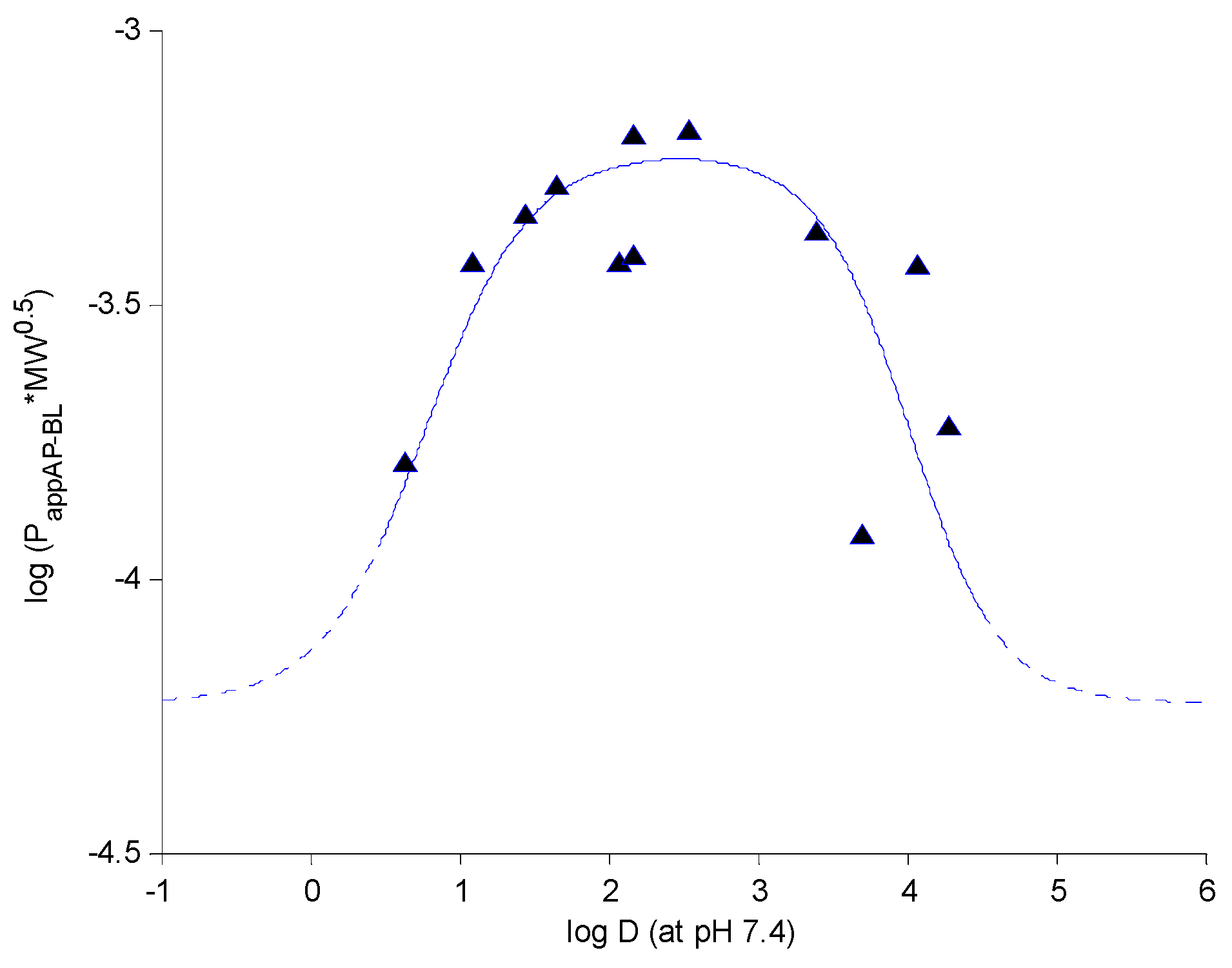
2.5. Structure-Permeability Relationship Analysis

3. Experimental Section
3.1. Chemicals and Reagents
3.2. Cell Culture
3.3. Transport Experiments
3.4. Standard Conditions of MDCK-pHaMDR Cell Monolayer
3.5. Time- and Concentration-Dependent Transport Experiments
3.6. Verapamil Inhibition of Transport
3.7. HPLC Analysis
3.8. Data Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chen, Y.F.; Tsai, H.Y.; Wu, T.S. Anti-inflammatory and analgesic activities from roots of Angelica pubescens. Planta Med. 1995, 61, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, J.; Gao, L. Anti-inflammatory and analgesic activity of R.A.P. (Radix Angelicae Pubescentis) ethanol extracts. Afr. J. Tradit. Complement. Altern. Med. 2013, 10, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Liu, B.R.; Hu, W.J.; Yu, L.X.; Qian, X.P. In vitro anticancer activity of aqueous extracts and ethanol extracts of fifteen traditional Chinese medicines on human digestive tumor cell lines. Phytother. Res. 2007, 21, 1102–1104. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.H.; Xu, S.X.; Yao, X.S.; Kobayashi, H. Angelol-type coumarins from Angelica pubescens f. biserrata, and their inhibitory effect on platelet aggregation. Phytochemistry 1995, 39, 1099–1101. [Google Scholar]
- Liao, J.F.; Jan, Y.M.; Huang, S.Y.; Wang, H.H.; Yu, L.L.; Chen, C.F. Evaluation with receptor binding assay on the water extracts of ten CNS-active Chinese herbal drugs. Proc. Natl. Sci. Counc. Repub. China B 1995, 19, 151–158. [Google Scholar] [PubMed]
- Pei, Y.; Li, D.X.; Sun, S.H. Experimental study on the effect of Radix Angelicae Pubescentis and its alcohol extract on apoptosis of brain cells in nature aging mice. Chin. J. Gerontol. 2005, 25, 959. [Google Scholar]
- Hu, Y.; Zhao, D.; Zhang, X.D.; Sun, D.; Hao, H.G.; Yang, J.X. Different extracts of Angelica pubescens inhibit H2O2-induced SH-SY5Y cells injury. Chin. J. Exp. Tradit. Med. Form. 2013, 19, 184–188. [Google Scholar]
- Zhu, M.D.; Cui, J.P.; Wang, C.X. The experimental study of the intervention effect of Duhuo on immune damage of model rats suffering from Alzheimer. Liaoning J. Tradit. Chin. Med. 2011, 38, 2085–2086. [Google Scholar]
- Liu, J.H.; Xu, S.X.; Yao, X.S. Chemical Constituents and Pharmacology Research Review of Angelicae Pubescentis Radix. J. Shenyang Coll. Pharm. 1994, 11, 143–150. [Google Scholar]
- Hornick, A.; Lieb, A.; Vo, N.P.; Rollinger, J.M.; Stuppner, H.; Prast, H. The coumarin scopoletin potentiates acetylcholine release from synaptosomes, amplifies hippocampal long-term potentiation and ameliorates anticholinergic- and age-impaired memory. Neuroscience 2011, 197, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Witaicenis, A.; Seito, L.N.; da Silveira Chagas, A.; de Almeida, L.D.; Luchini, A.C.; Rodrigues-Orsi, P.; Cestari, S.H.; Di Stasi, L.C. Antioxidant and intestinal anti-inflammatory effects of plant-derived coumarin derivatives. Phytomedicine 2014, 21, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Wen, Q.; Liang, W.; Kang, T.; Ren, L.; Zhang, N.; Zhao, D.; Sun, D.; Yang, J. Osthole reverses beta-amyloid peptide cytotoxicity on neural cells by enhancing cyclic AMP response element-binding protein phosphorylation. Biol. Pharm. Bull. 2013, 36, 1950–1958. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Qu, S.; Wang, J.; He, X.; Lin, W.; Zhen, H.; Zhang, X. Neuroprotective effects of osthole pretreatment against traumatic brain injury in rats. Brain Res. 2012, 1433, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.K.; Lim, J.P.; Yang, J.H.; Eom, D.O.; Eun, J.S.; Leem, K.H. Acetylcholinesterase inhibitors from the roots of Angelica dahurica. Arch. Pharm. Res. 2002, 25, 856–859. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.D.; Tan, R.X.; Woo, A.Y.; Cheng, C.H. Inhibition of rat brain monoamine oxidase activities by psoralen and isopsoralen: Implications for the treatment of affective disorders. Pharmacol. Toxicol. 2001, 88, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Huong, D.T.; Choi, H.C.; Rho, T.C.; Lee, H.S.; Lee, M.K.; Kim, Y.H. Inhibitory activity of monoamine oxidase by coumarins from Peucedanum japonicum. Arch. Pharm. Res. 1999, 22, 324–326. [Google Scholar] [CrossRef] [PubMed]
- Sigurdsson, S.; Gudbjarnason, S. Inhibition of acetylcholinesterase by extracts and constituents from Angelica archangelica and Geranium sylvaticum. Z. Naturforsch. C 2007, 62, 689–693. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.W.; Guo, Q.M.; Wang, Y. Absorption and transport of six coumarins isolated from the roots of Angelica pubescens f. biserrata in human Caco-2 cell monolayer model. J. Chin. Integr. Med. 2008, 6, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Long, J.; Li, S.N.; Yang, X.W. Absorption characteristics of four linear furocoumarins across human intestinal epithelial in a model of Caco-2 cell monolayer. Chin. J. New Drugs 2009, 18, 1865–1871. [Google Scholar]
- Li, S.N.; Yang, X.W. Absorption and transportation characteristic of six linear furocoumarins in a model of Caco-2 cell monolayer in human intestine. Chin. Tradit. Herb Drugs 2011, 42, 96–102. [Google Scholar]
- Cecchelli, R.; Berezowski, V.; Lundquist, S.; Culot, M.; Renftel, M.; Dehouck, M.P.; Fenart, L. Modelling of the blood-brain barrier in drug discovery and development. Nat. Rev. Drug Discov. 2007, 6, 650–661. [Google Scholar] [CrossRef] [PubMed]
- Abbott, N.J.; Patabendige, A.A.; Dolman, D.E.; Yusof, S.R.; Begley, D.J. Structure and function of the blood-brain barrier. Neurobiol. Dis. 2010, 37, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Rager, J.D.; Weinstein, K.; Kardos, P.S.; Dobson, G.L.; Li, J.; Hidalgo, I.J. Evaluation of the MDR-MDCK cell line as a permeability screen for the blood-brain barrier. Int. J. Pharm. 2005, 288, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Hellinger, E.; Veszelka, S.; Tóth, A.E.; Walter, F.; Kittel, A.; Bakk, M.L.; Tihanyi, K.; Háda, V.; Nakagawa, S.; Duy, T.D.H.; et al. Comparison of brain capillary endothelial cell-based and epithelial (MDCK-pHaMDR, Caco-2, and VB-Caco-2) cell-based surrogate blood-brain barrier penetration models. Eur. J. Pharm. Biopharm. 2012, 82, 340–351. [Google Scholar] [CrossRef] [PubMed]
- Madgula, V.L.M.; Avula, B.; Yu, Y.B.; Wang, Y.H.; Tchantchou, F.; Fisher, S.; Luo, Y.; Khan, I.A.; Khan, S.I. Intestinal and blood-brain barrier permeability of ginkgolides and bilobalide: In vitro and in vivo approaches. Planta Med. 2010, 76, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Madgula, V.L.M.; Avula, B.; Reddy, N.V.L.; Khan, I.A.; Khan, S.I. Transport of decursin and decursinol angelate across Caco-2 and MDR-MDCK cell monolayers: In vitro models for intestinal and blood-brain barrier permeability. Planta Med. 2007, 73, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Pastan, I.; Gottesman, M.M.; Ueda, K.; Lovelace, E.; Rutherford, A.V.; Willingham, M.C. A retrovirus carrying an MDR1 cDNA confers multidrug resistance and polarized expression of P-glycoprotein in MDCK cells. Proc. Natl. Acad. Sci. USA 1988, 85, 4486–4490. [Google Scholar] [CrossRef] [PubMed]
- Camenisch, G.; Folkers, G.; van de Waterbeemd, H. Review of theoretical passive drug absorption models: Historical background, recent developments and limitations. Pharm. Acta. Helv. 1996, 71, 309–327. [Google Scholar] [CrossRef]
- Wils, P.; Warnery, A.; Phung-Ba, V.; Legrain, S.; Scherman, D. High lipophilicity decreases drug transport across intestinal epithelial cells. J. Pharmacol. Exp. Ther. 1994, 269, 654–658. [Google Scholar] [PubMed]
- Yang, X.W.; Huang, X.; Ma, L.; Wu, Q.; Xu, W. The intestinal permeability of neolignans from the seeds of Myristica fragrans in the Caco-2 cell monolayer model. Planta Med. 2010, 76, 1587–1591. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.Y.; Zhang, B.G.; Yang, X.W. Studies on the chemical constituents of the root of Angelica pubescens f. biserrata. Pharm. J. Chin. P.L.A. 2007, 23, 241–245. [Google Scholar]
- Yang, X.W.; Zhang, C.Y.; Zhang, B.G.; Lu, Y.; Luan, J.W.; Zheng, Q.T. Novel coumarin and furan from the roots of Angelica pubescens f. biserrata. J. Asian Nat. Prod. Res. 2009, 11, 698–703. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.Y.; Hu, H.H.; Wang, X.J.; Yu, L.S.; Jiang, H.D.; Chen, J.Z.; Lou, Y.; Zeng, S. Inhibitory Effects of Neochamaejasmin B on P-Glycoprotein in MDCK-hMDR1 Cells and Molecular Docking of NCB Binding in P-Glycoprotein. Molecules 2015, 20, 2931–2948. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples are not available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.-F.; Xu, W.; Song, W.; Ye, M.; Yang, X.-W. Transport of Twelve Coumarins from Angelicae Pubescentis Radix across a MDCK-pHaMDR Cell Monolayer—An in Vitro Model for Blood-Brain Barrier Permeability. Molecules 2015, 20, 11719-11732. https://doi.org/10.3390/molecules200711719
Yang Y-F, Xu W, Song W, Ye M, Yang X-W. Transport of Twelve Coumarins from Angelicae Pubescentis Radix across a MDCK-pHaMDR Cell Monolayer—An in Vitro Model for Blood-Brain Barrier Permeability. Molecules. 2015; 20(7):11719-11732. https://doi.org/10.3390/molecules200711719
Chicago/Turabian StyleYang, Yan-Fang, Wei Xu, Wei Song, Min Ye, and Xiu-Wei Yang. 2015. "Transport of Twelve Coumarins from Angelicae Pubescentis Radix across a MDCK-pHaMDR Cell Monolayer—An in Vitro Model for Blood-Brain Barrier Permeability" Molecules 20, no. 7: 11719-11732. https://doi.org/10.3390/molecules200711719
APA StyleYang, Y. -F., Xu, W., Song, W., Ye, M., & Yang, X. -W. (2015). Transport of Twelve Coumarins from Angelicae Pubescentis Radix across a MDCK-pHaMDR Cell Monolayer—An in Vitro Model for Blood-Brain Barrier Permeability. Molecules, 20(7), 11719-11732. https://doi.org/10.3390/molecules200711719