
Proteomic Analysis of Tung Tree (Vernicia fordii) Oilseeds during the Developmental Stages

Abstract
:1. Introduction
2. Results
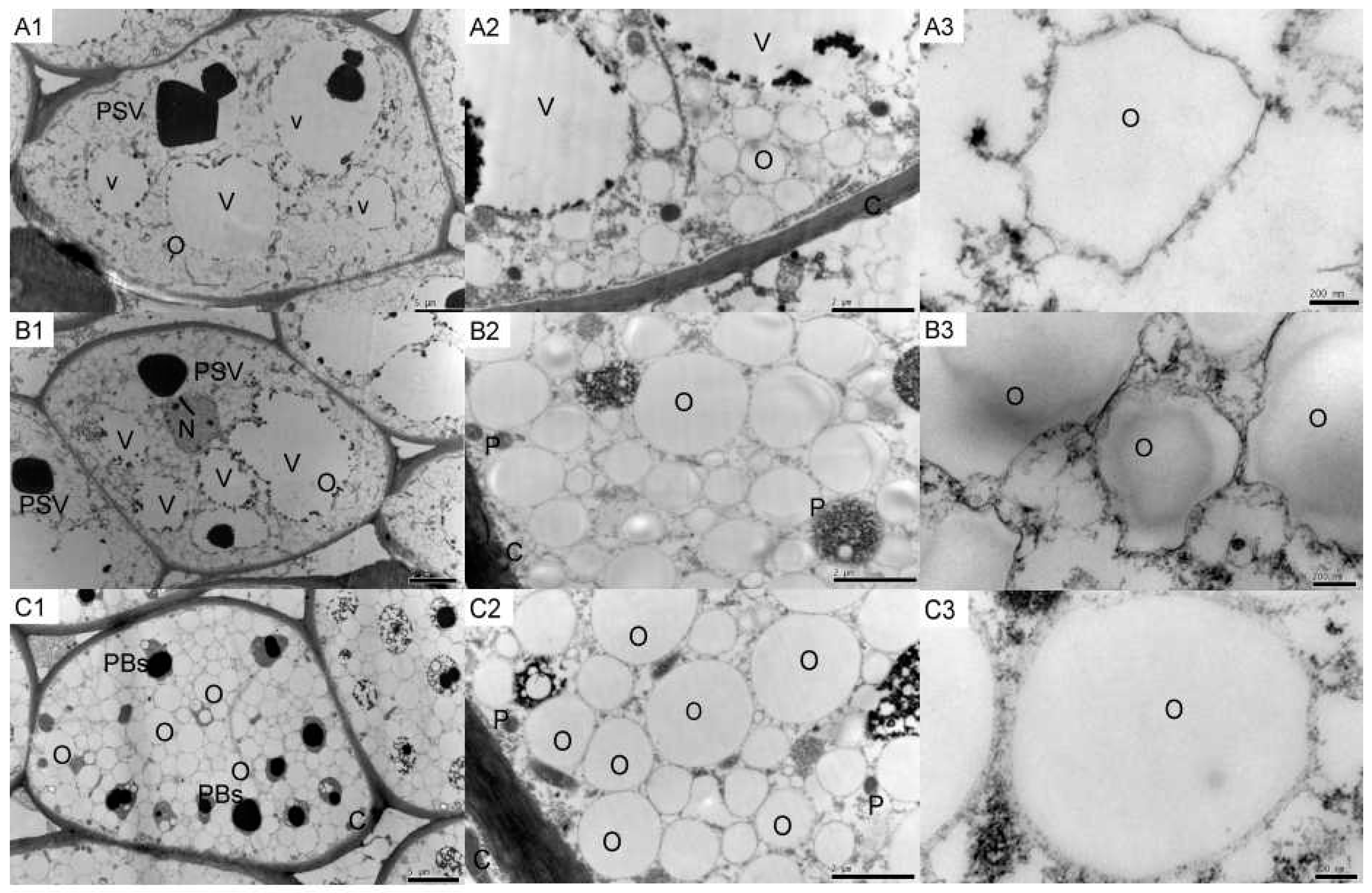
2.1. Tung Endosperm Cell Ultrastructure
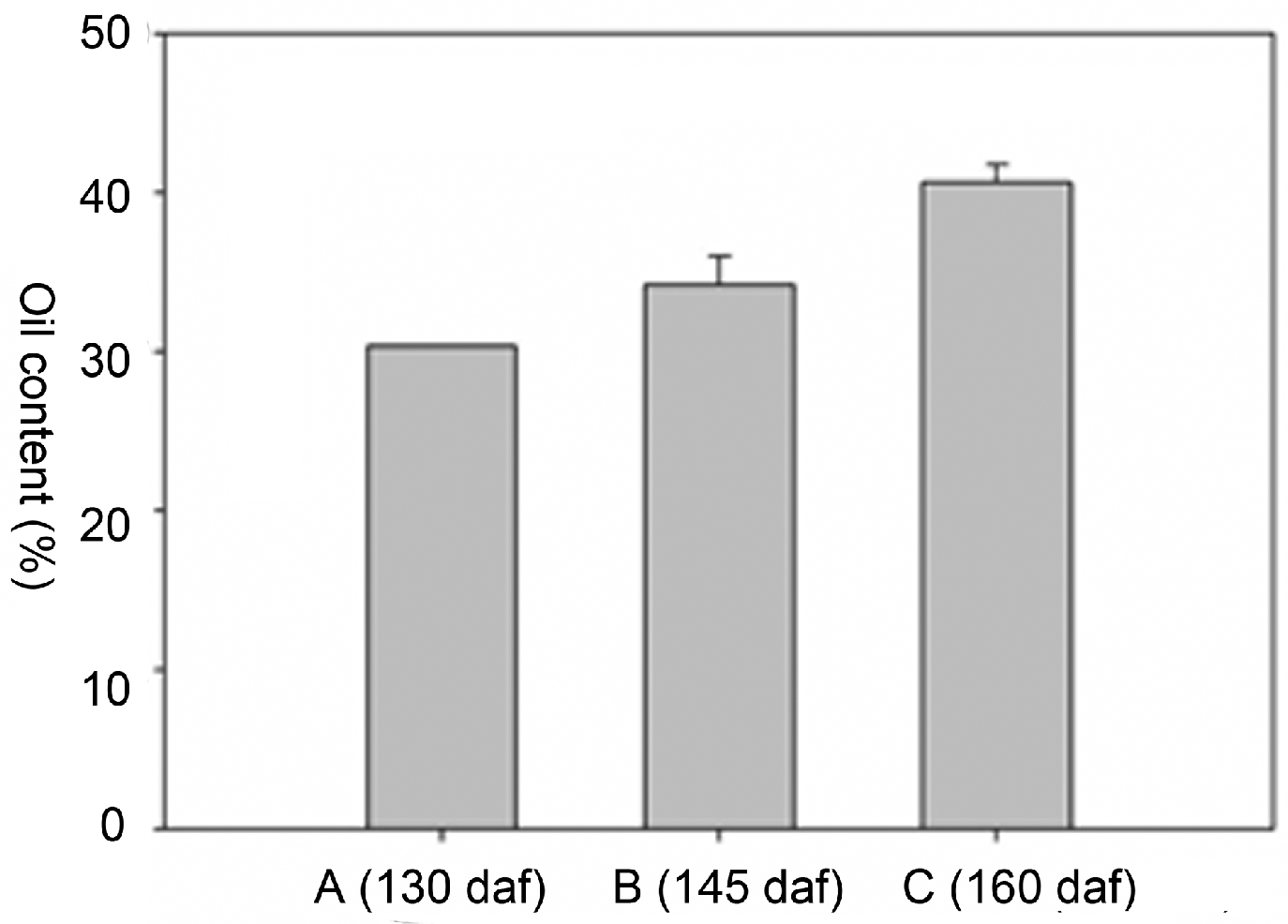
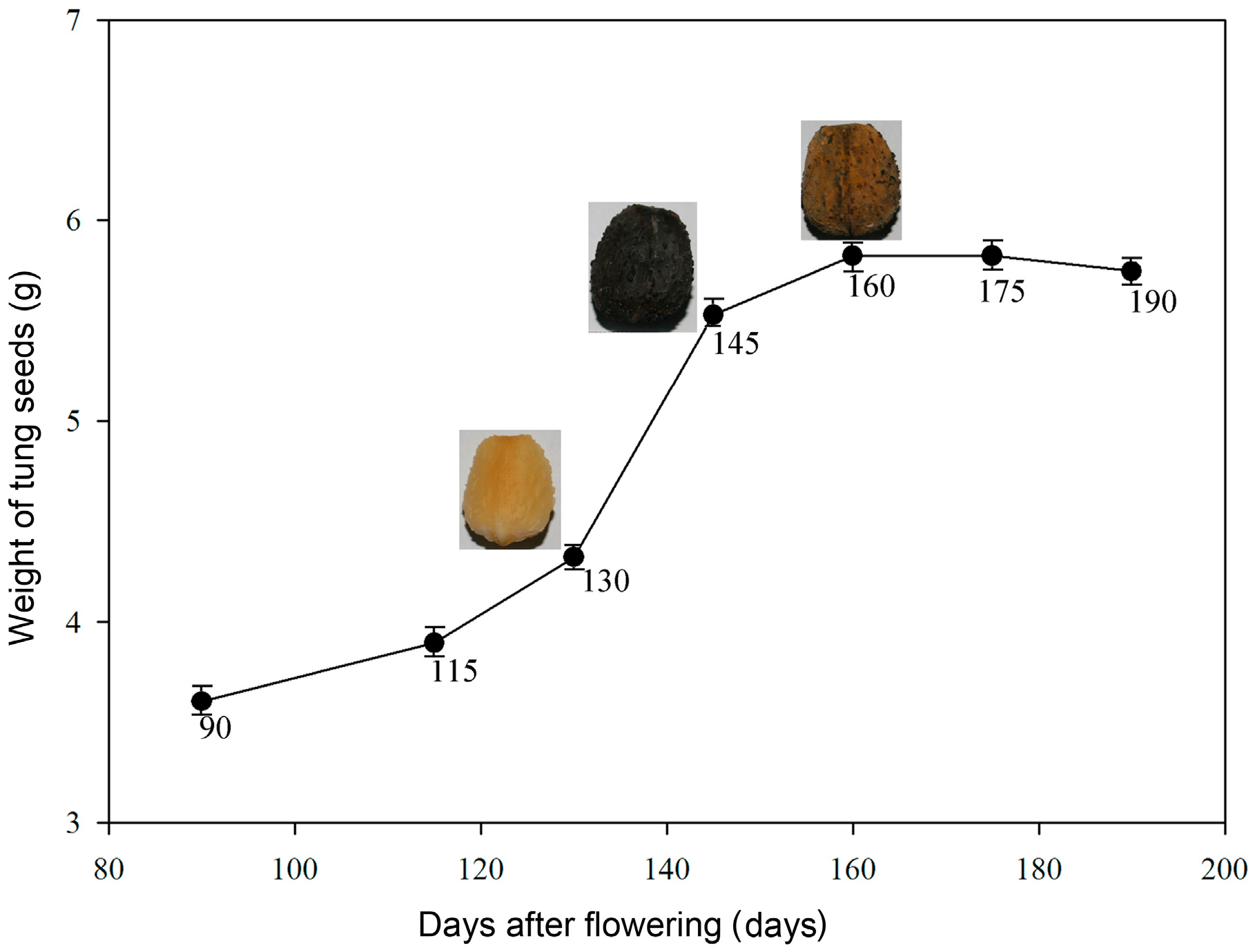
2.2. Oil Content and Fatty Acid (FA) Composition
2.3. Comparison of the Different Protein Extraction Methods
2.4. Identification of Differentially Expressed Proteins in Different Functional Categories
2.4.1. Energy Metabolism
2.4.2. Fatty Acid Metabolism
2.4.3. Defense-Related
3. Discussion
3.1. Efficient Extraction Methods for Tung Tree Seed Proteins
3.2. The Relationship between Oil Bodies and Oil Content in Seeds During Developmental Stages
3.3. Seed Reserve Synthesis during Development
3.4. Some Enzymes Were Undetectable during Seed Development
4. Materials and methods
4.1. Materials
4.2. Tissue Preparation for Transmission Electron Microscopy
4.3. Oil Content and Fatty Acid Composition Analysis
4.4. Protein Extraction, Two-Dimensional Gel Electrophoresis and Data Analysis
4.5. In-Gel Protein Digestion and MALDI-TOF/MS Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sghaier-Hammami, B.; Valledor, L.; Drira, N.; Jorrin-Novo, J.V. Proteomic analysis of the development and germination of date palm (Phoenix dactylifera L.) zygotic embryos. Proteomics 2009, 9, 2543–2554. [Google Scholar] [CrossRef] [PubMed]
- Miernyk, J.A.; Hajduch, M. Seed proteomics. Proteomics 2011, 74, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Møller, I.M.; Song, S.Q. Proteomics of desiccation tolerance during development and germination of maize embryos. Proteomics 2012, 75, 1247–1262. [Google Scholar] [CrossRef] [PubMed]
- Schiltz, S.; Gallardo, K.; Huart, M.; Negroni, L.; Sommerer, N.; Burstin, J. Proteome reference maps of vegetative tissues in Pea. An investigation of nitrogen mobilization from leaves during seed filling. Plant Physiol. 2004, 135, 2241–2260. [Google Scholar] [CrossRef] [PubMed]
- Chibani, K.; Ali-Rachedi, S.; Job, C.; Job, D.; Jullien, M.; Grappin, P. Proteomic analysis of seed dormancy in Arabidopsis. Plant Physiol. 2006, 142, 1493–1510. [Google Scholar] [CrossRef] [PubMed]
- Sheoran, I.S.; Olson, D.J.H.; Ross, A.R.S.; Sawhney, V.K. Proteome analysis of mbryo and endosperm from germination tomato seesds. Proteomics 2005, 5, 3752–3764. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Li, X.; Wang, X.; Chen, H.; Chen, F.; Shen, S. Proteomic analysis of rice (Oryza sativa) seeds during germination. Proteomics 2007, 7, 3358–3368. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.F.; Liu, Y.J.; Liu, Y.; Chen, H.; Chen, F.; Shen, S.H. Proteomic analysis of oil mobilization in seed germination and postgermination development of Jatropha curcas. Proteome Res. 2009, 8, 1441–1451. [Google Scholar] [CrossRef] [PubMed]
- Wong, P.F.; Abubakar, S. Post-germination changes in Hevea brasiliensis seeds proteome. Plant Sci. 2005, 169, 303–311. [Google Scholar] [CrossRef]
- Finnie, C.; Melchior, S.; Roepstorff, P.; Svensson, B. Proteome analysis of grain filling and seed maturation in barly. Plant Physiol. 2002, 129, 1308–1319. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Yang, Z.; Yang, M.; Shen, S. The differential proteome of endosperm and embryo from mature seed of Jatropha curcas. Plant Sci. 2011, 181, 660–666. [Google Scholar] [CrossRef] [PubMed]
- Nouri, M.Z.; Komatsu, S. Comparative analysis of soybean plasma membrane proteins under osmotic stress using gel-based and LC MS/MS-based proteomics approaches. Proteomics 2010, 10, 1930–1945. [Google Scholar] [CrossRef] [PubMed]
- Sheoran, I.S.; Pedersen, E.J.; Ross, A.R.S.; Sawhney, V.K. Dynamics of protein expression during pollen germination in canola (Brassica napus). Planta 2009, 230, 779–793. [Google Scholar] [CrossRef] [PubMed]
- Hajduch, M.; Casteel, J.E.; Hurrelmeyer, K.E.; Song, Z.; Agrawal, G.K.; Thelen, J.J. Proteomic analysis of seed filling in Brassica napus. Developmental characterization of metabolic isozymes using high-resolution two-dimensional gel electrophoresis. Plant Physiol. 2006, 141, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Potter, G. The domestic tung industry. I. Production and improvement of the tung tree. Econ. Bot. 1959, 13, 328–342. [Google Scholar] [CrossRef]
- Sonntag, N. Composition and characteristics of individual fats and oils. Bailey’s Ind. Oil Fat Prod. 1979, 1, 289–477. [Google Scholar]
- Park, J.; Kim, D.; Wang, Z. Production and characterization of biodiesel from tung oil. Appl. Biochem. Biotechnol. 2008, 148, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.; Jiang, W.; Lu, H.; Liang, B. Properties of tung oil biodiesel and its blends with 0# diesel. Bioresour. Technol. 2010, 101, 826–828. [Google Scholar] [PubMed]
- Chen, Y.; Chen, J.; Chang, C.; Chang, C. Biodiesel production from tung (Vernicia montana) oil and its blending properties in different fatty acid compositions. Bioresour. Technol. 2010, 101, 9521–9526. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Chapital, D.C.; Shockey, J.M.; Klasson, K.T. Expression of tung tree diacylglycerol acyltransferase 1 in E. coli. BMC Biotechnol. 2011, 11, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Dyer, J.M.; Chapital, D.C.; Kuan, J.C.W.; Mullen, R.T.; Turner, C.; McKeon, T.A.; Pepperman, A.B. Molecular analysis of a bifunctional fatty acid conjugase/desaturase from tung. Implications for the evolution of plant fatty acid diversity. Plant Physiol. 2002, 130, 2027–2038. [Google Scholar] [CrossRef] [PubMed]
- Shockey, J.M.; Gidda, S.K.; Chapital, D.C.; Kuan, J.C.; Dhanoa, P.K.; Bland, J.M.; Rothstein, S.J.; Mullen, R.T.; Dyer, J.M. Tung tree DGAT1 and DGAT2 have nonredundant functions in triacylglycerol biosynthesis and are localized to different subdomains of the endoplasmic reticulum. Plant Cell 2006, 18, 2294–2313. [Google Scholar] [CrossRef] [PubMed]
- Turchetto-Zolet, A.C.; Maraschin, F.S.; de Morais, G.L.; Cagliari, A.; Andrade, C.M.B.; Margis-Pinheiro, M.; Margis, R. Evolutionary view of acyl-CoA diacylglycerol acyltransferase (DGAT), a key enzyme in neutral lipid biosynthesis. BMC Evol. Biol. 2011, 11, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Li, X.J.; Yang, M.F.; Chen, H.; Qu, L.Q.; Chen, F.; Shen, S.H. Abscisic acid pretreatment enhances salt tolerance of rice seedlings: Proteomic evidence. Biochim. Biophys. Acta 2010, 1804, 929–940. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Kim, M.D.; Yang, K.S.; Kwon, S.Y.; Kim, S.H.; Kim, J.S.; Yun, D.J.; Kwak, S.S.; Lee, H.S. Enhanced tolerance of transgenic potato plants overexpressing nucleoside diphosphate kinase 2 against multiple environment stresses. Transgenic Res. 2008, 17, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Park, S.J.; Jang, B.; Jung, C.H.; Ahn, S.J.; Goh, C.H.; Cho, K.; Han, O.; Kang, H. Functional characterization of a glycine-rich RNA-binding protein 2 in Arabidopsis thaliana under abiotic stress conditions. Plant J. 2007, 50, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Chen, H.; Li, X.J.; Yang, M.F.; Liu, G.S.; Shen, S.H. A comparative proteomic analysis of rice seedlings under various high-temperature stresses. Biochim. Biophys. Acta 2009, 1794, 1625–1634. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.I.; An, C.S. Cloning and expression of mitochondrial MnSOD from the small radish (Raphanus sativus L.). Mol. Cells 2003, 16, 194–200. [Google Scholar] [PubMed]
- Isaacson, T.; Damasceno, C.M.B.; Saravanan, R.S.; He, Y.H.; Catalá, C.; Saladié, M.; Rose, J.K.C. Sample extraction techniques for enhanced proteomic analysis of plant tissues. Nat. Protocol. 2006, 1, 769–774. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, R.S.; Rose, J.K.C. A critical evaluation of sample extraction techniques for enhanced proteomic analysis of recalcitrant plant tissues. Proteomics 2004, 4, 2522–2532. [Google Scholar] [CrossRef] [PubMed]
- Hurkman, W.J.; Tanaka, C.K. Solubilisation of plant membrane proteins for analysis by two-dimensional gel electrophoresis. Plant Physiol. 1986, 81, 802–806. [Google Scholar] [CrossRef] [PubMed]
- Sheoran, I.; Ross, A.; Olson, D.; Sawhney, V. Compatibility of plant protein extraction methods with mass spectrometry for proteome analysis. Plant Sci. 2009, 176, 99–104. [Google Scholar] [CrossRef]
- De la Fuente, M.; Borrajo, A.; Bermúdez, J.; Lores, M.; Alonso, J.; López, M.; Santalla, M.; de Ron, A.M.; Zapata, C.; Alvarez, G. 2-DE-based proteomic analysis of common bean (Phaseolus vulgaris L.) seeds. Proteomics 2011, 74, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Vignani, R.; Scali, M.; Cresti, M. A universal and rapid protocol for protein extraction from recalcitrant plant tissues for proteomic analysis. Electrophoresis 2006, 27, 2782–2786. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.Y.; Wang, X.F.; Zhang, G.M.; Liu, G.H.; Hua, W.; Wang, H.Z. Unusually large oilbodies are highly correlated with lower oil content in Brassica napus. Plant Cell Rep. 2009, 28, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Ting, J.T.L.; Lee, K.Y.; Ratnayake, C.; Platt, K.A.; Balsamo, R.A.; Huang, A.H.C. Oleosin genes in maize kernels having diverse oil contents are constitutively expressed independent of oil contents. Size and shape of intracellular oil-bodies are determined by the oleosins/oils ratio. Planta 1996, 199, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Mantese, A.L.; Medan, D.; Hall, A.J. Achene structure development and lipid accumulation in sunflower cultivars differing in oil content at maturity. Ann. Bot. 2006, 97, 999–1010. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.M.; Baudet, M.; Cuine, S.; Adriano, J.; Barthe, D.; Billon, E.; Bruley, C.; Beisson, F.; Peltier, G.; Ferro, M.; et al. Proteomic profiling of oil bodies isolated from the unicellular green microalga Ch-lamydomonas reinhardtii: With focus on proteins involved in lipid metabolism. Proteomics 2011, 11, 4266–4273. [Google Scholar] [CrossRef] [PubMed]
- Murphy, D.J. Structure, function and biogenesis of storage lipid bodies and oleosins in plants. Prog. Lipid Res. 1993, 2, 247–280. [Google Scholar] [CrossRef]
- Huang, A.H.C. Oil bodies and oleosins in seeds. Annu. Rev. Plant Physiol. 1992, 43, 177–200. [Google Scholar] [CrossRef]
- Tnani, H.; López, I.; Jouenne, T.; Vicient, C.M. Protein composition analysis of oil bodies from maize embryos during germination. Plant Physiol. 2011, 168, 510–513. [Google Scholar] [CrossRef] [PubMed]
- Poxleitner, M.; Rogers, S.W.; Samuels, A.L.; Browse, J.; Rogers, J.C. A role for caleosin in degradation of oil-body storage lipid during seed germination. Plant J. 2006, 47, 917–933. [Google Scholar] [CrossRef] [PubMed]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef] [PubMed]
- Granier, F. Extraction of plant proteins for two-dimensional electrophoresis. Electrophoresis 1988, 11, 712–718. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Scali, M.; Vignani, R.; Spadafora, A.; Sensi, E.; Mazzuca, S.; Cresti, M. Protein extraction for two-dimensional electrophoresis from olive leaf, a plant tissue containing high levels of interfering compounds. Electrophoresis 2003, 14, 2369–2375. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Tai, F.; Chen, S. Optimizing protein extraction from plant tissues for enhanced proteomics analysis. J. Sep. Sci. 2008, 31, 2032–2039. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Z.Y.; Wang, Y.D.; Chen, Y.C.; Han, X.J.; Cui, Q.Q. Comparison of three protein extraction methods for tung trees (Vernicia fordii) seed. Bull. Bot. Res. 2013, 33, 91–97. [Google Scholar]
- Sample Availability: Samples of the compounds in the article are available from the authors.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | Fatty Acid | |||||
|---|---|---|---|---|---|---|
| Palmitic Acid | Stearic Acid | Oleic Acid | Linoleic Acid | Linolenic Acid | Eleostearic Acid | |
| 8.25 (A) | 3.74 ± 0.02 | 2.75 ± 0.03 | 7.42 ± 0.08 | 13.82 ± 0.12 | 0.21 ± 0.15 | 72.28 ± 0.06 |
| 9.9 (B) | 2.50 ± 0.05 | 1.95 ± 0.01 | 7.99 ± 0.06 | 9.11 ± 0.01 | - | 77.47 ± 0.05 |
| 9.26 (C) | 2.35 ± 0.06 | 2.17 ± 0.02 | 8.03 ± 0.08 | 7.85 ± 0.05 | - | 77.49 ± 0.02 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhan, Z.; Chen, Y.; Shockey, J.; Han, X.; Wang, Y. Proteomic Analysis of Tung Tree (Vernicia fordii) Oilseeds during the Developmental Stages. Molecules 2016, 21, 1486. https://doi.org/10.3390/molecules21111486
Zhan Z, Chen Y, Shockey J, Han X, Wang Y. Proteomic Analysis of Tung Tree (Vernicia fordii) Oilseeds during the Developmental Stages. Molecules. 2016; 21(11):1486. https://doi.org/10.3390/molecules21111486
Chicago/Turabian StyleZhan, Zhiyong, Yicun Chen, Jay Shockey, Xiaojiao Han, and Yangdong Wang. 2016. "Proteomic Analysis of Tung Tree (Vernicia fordii) Oilseeds during the Developmental Stages" Molecules 21, no. 11: 1486. https://doi.org/10.3390/molecules21111486
APA StyleZhan, Z., Chen, Y., Shockey, J., Han, X., & Wang, Y. (2016). Proteomic Analysis of Tung Tree (Vernicia fordii) Oilseeds during the Developmental Stages. Molecules, 21(11), 1486. https://doi.org/10.3390/molecules21111486