
Clarification of the Antagonistic Effect of the Lipopeptides Produced by Bacillus amyloliquefaciens BPD1 against Pyricularia oryzae via In Situ MALDI-TOF IMS Analysis

 , ,
, , 

Abstract
:1. Introduction
2. Results and Discussion
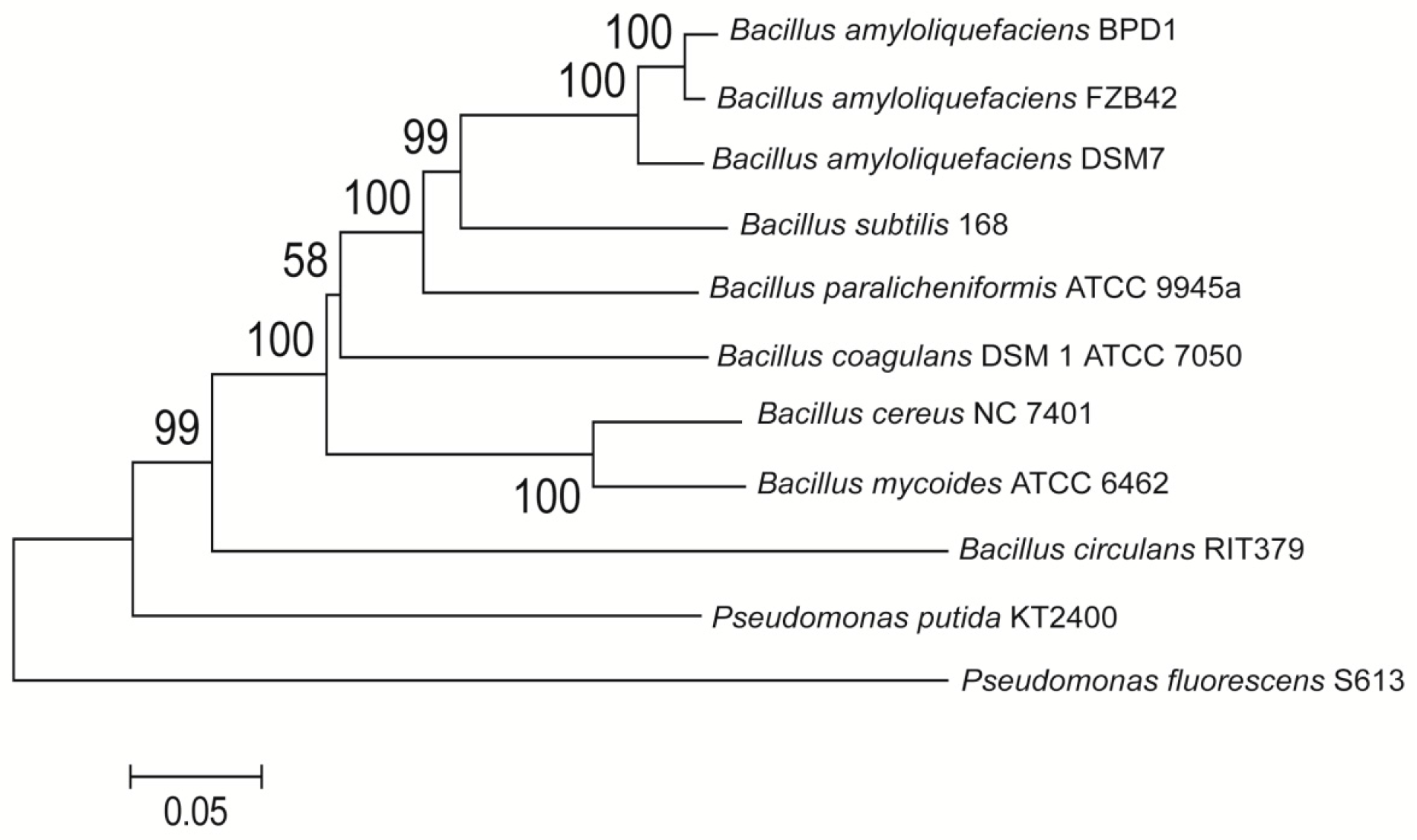
2.1. Identification of the Strain Ba-BPD1
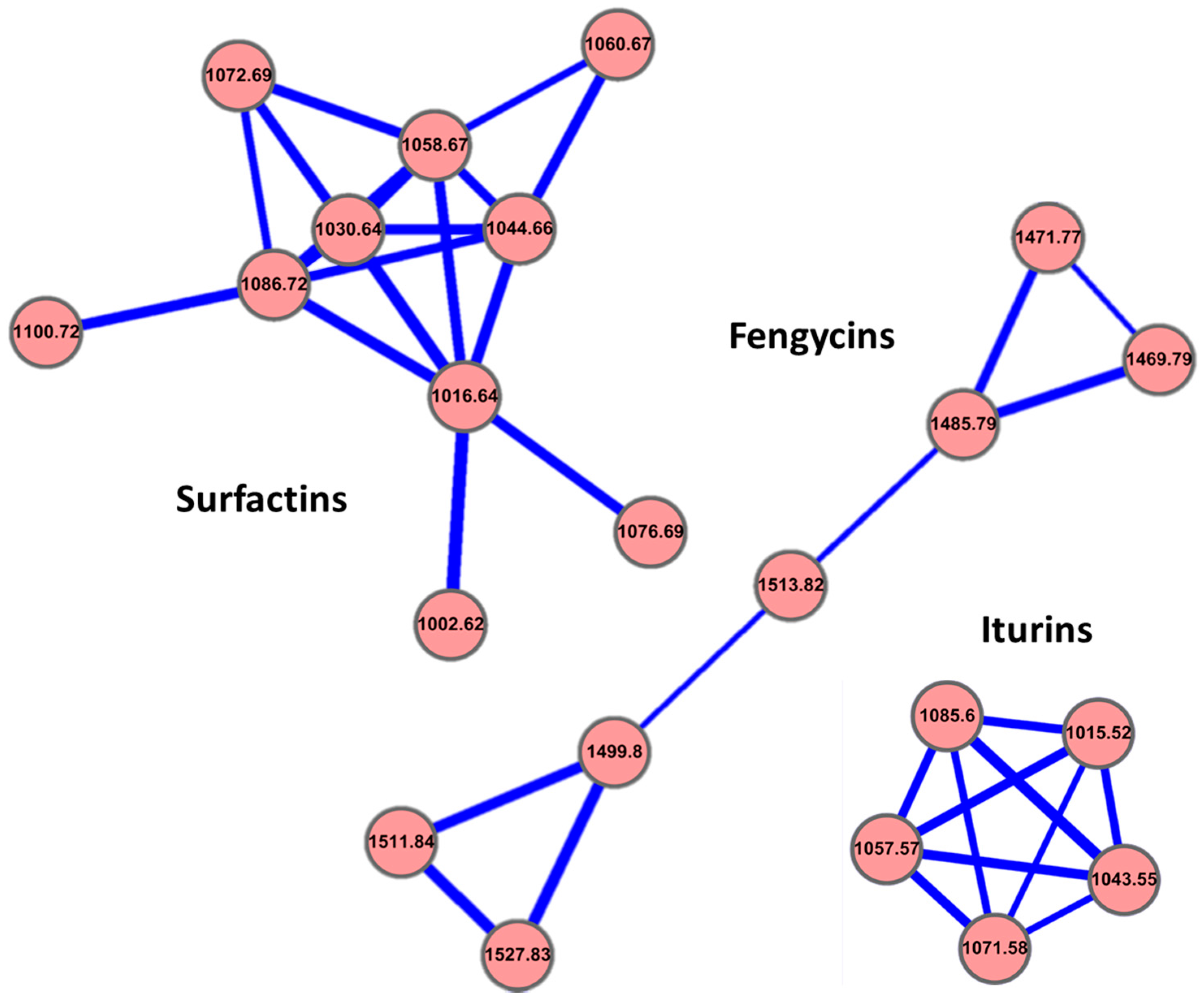
2.2. Identification of the Lipopeptides of Ba-BPD1
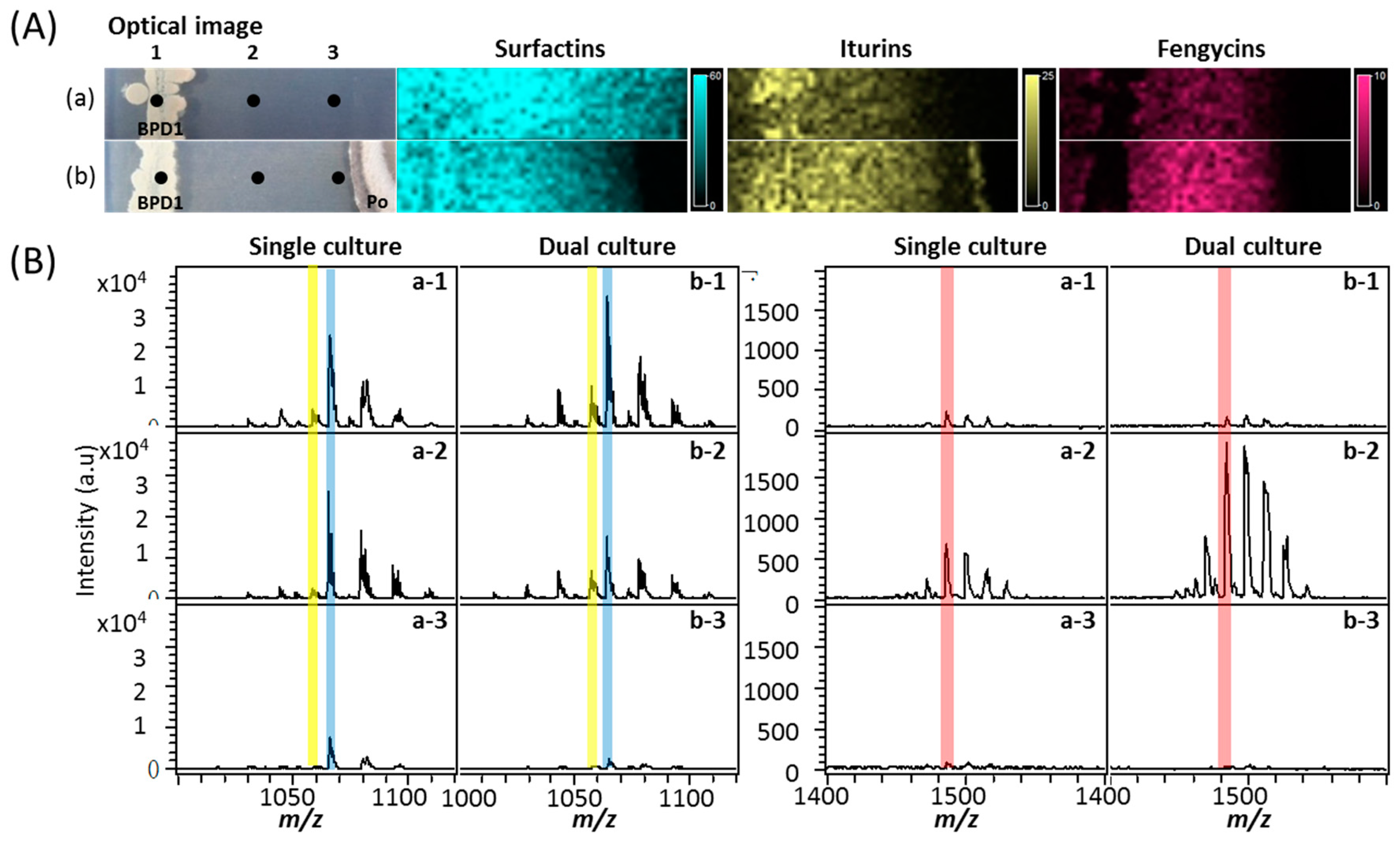
2.3. MALDI-TOF IMS Analysis
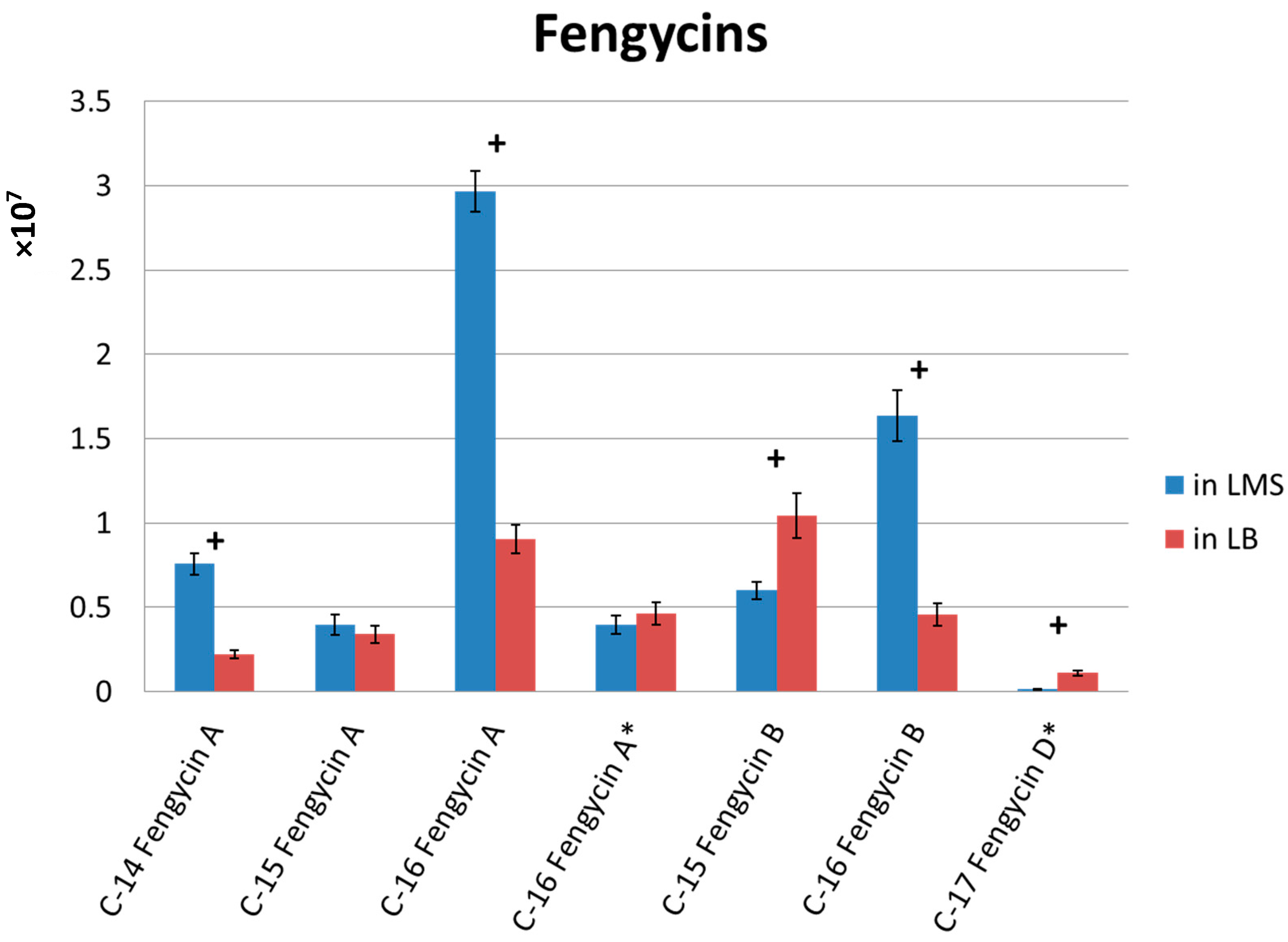
2.4. Fermentation Medium Tested for Fengycin Production
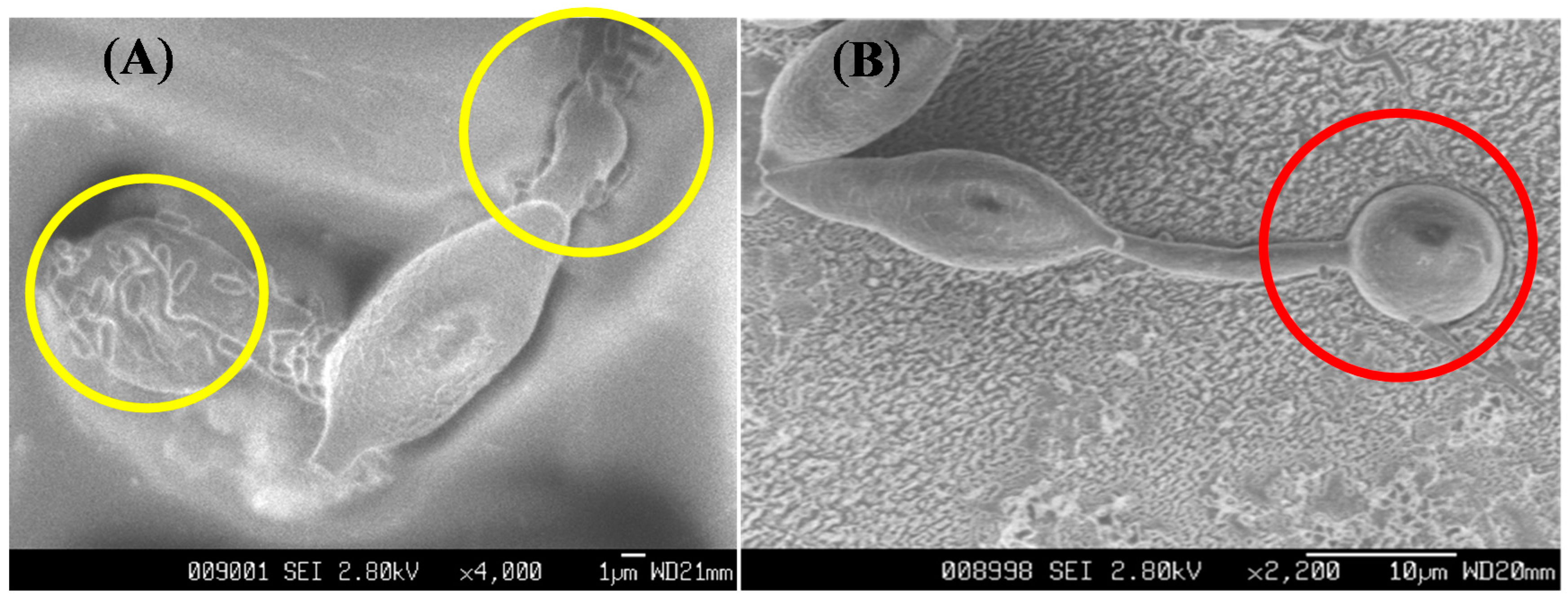
2.5. Antifungal Bioassay and SEM Observations
2.6. The Effect of Ba-BPD1 Broth on the Germination of Conidia of P. oryzae
3. Experimental Section
3.1. Chemicals
3.2. Strain and Culture Conditions
3.3. LC-MS and LC-MS/MS Analysis
3.4. MALDI-TOF IMS Analysis
3.5. Antifungal Bioassay
3.6. Observation of Conidial Morphology
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Luo, Y.; Teng, P.S.; Fabellar, N.G.; TeBeest, D.O. Risk analysis of yield losses caused by rice leaf blast associated with temperature changes above and below for five Asian countries. Agric. Ecosyst. Environ. 1998, 2, 197–205. [Google Scholar] [CrossRef]
- Jia, Y.; McAdams, S.A.; Bryan, G.T.; Hershey, H.P.; Valent, B. Direct interaction of resistance gene and avirulence gene products confers rice blast resistance. Embo J. 2000, 2, 4004–4014. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, S.Y. Estimating losses from rice blast in India. In The Rice Blast Disease; Johns Hopkins Press: Baltimore, MD, USA, 1965; pp. 203–221. [Google Scholar]
- Awodera, V.A.; Esuruoso, O.F. Reduction in grain yield of two rice varieties infected by rice blast disease in Nigeria. Nigerian Agric. J. 1975, 2, 170–173. [Google Scholar]
- Ou, S.H. Rice Diseases; CAB International Mycological Institute: Kew, UK, 1987. [Google Scholar]
- Prabhu, A.S.; Morais, O.P. Blast disease management in upland rice in Brazil. In Progress in Upland Rice Research; International Rice Research Institute (IRRI): Los Baños, Philippines, 1986; pp. 383–392. [Google Scholar]
- Ribot, C.; Hirsch, J.; Balzergue, S.; Tharreau, D.; Notteghem, J.L.; Lebrun, M.H.; Morel, J.B. Susceptibility of rice to the blast fungus Magnaporthe grisea. J. Plant Physiol. 2008, 2, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Manidipa, R.; Dutta, S.G.; Venkata, R.C. Pseudomonads: Potential biocontrol agents of rice diseases. Res. J. Agric. For. Sci. 2013, 2, 19–25. [Google Scholar]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 2, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Shyamala, L.; Sivakumaar, P.K. Integrated control of blast disease of rice using the antagonistic Rhizobacteria Pseudomonas fluorescens and the resistance inducing chemical salicylic acid. Int. J. Res. Pure Appl. Microbiol. 2012, 2, 59–63. [Google Scholar]
- Arguelles-Arias, A.; Ongena, M.; Halimi, B.; Lara, Y.; Brans, A.; Joris, B.; Fickers, P. Bacillus amyloliquefaciens GA1 as a source of potent antibiotics and other secondary metabolites for biocontrol of plant pathogens. Microb. Cell Fact. 2009, 2, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Stein, T. Bacillus subtilis antibiotics: Structures, syntheses and specific functions. Mol. Microbiol. 2005, 2, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Liu, X.; Zhou, H.; Wang, X.; Chen, Z. Identification of four NRPS gene clusters in Bacillus subtilis 916 for four families of lipopeptides biosynthesis and evaluation of their intricate functions to the typical phenotypic features. Appl. Environ. Microbiol. 2014, 2, 422–443. [Google Scholar]
- Il, K.P.; Ryu, J.; Kim, Y.H.; ChI, Y.T. Production of biosurfactant lipopeptides iturin A, fengycin, and surfactin A from Bacillus subtilis CMB32 for control of Colletotrichum gloeosporioides. J. Microbiol. Biotechnol. 2010, 2, 138–145. [Google Scholar]
- Liu, J.; Zhou, T.; He, D.; Li, X.; Wu, H.; Liu, W.; Gao, X. Functions of lipopeptides bacillomycin D and fengycin in antagonism of Bacillus amyloliquefaciens C06 towards Monilinia fructicola. J. Mol. Microb. Biotechnol. 2011, 2, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.G.; Dong, W.X.; Li, S.Z.; Lu, X.Y.; Wang, P.P.; Zhang, X.Y.; Wang, Y.; Ma, P. Fengycin produced by Bacillus subtilis NCD-2 plays a major role in biocontrol of cotton seedling damping-off disease. Microbiol. Res. 2014, 2, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Romero, D.; de Vicente, A.; Rakotoaly, R.H.; Dufour, S.E.; Veening, J.W.; Arrebola, E.; et al. The iturin and fengycin families of lipopeptides are key factors in antagonism of Bacillus subtilis toward Podosphaera fusca. Mol. Plant Microb. Interact. 2007, 2, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Maget-Dana, R.; Thimon, L.; Fau-Peypoux, F.; Peypoux, F.; Fau-Ptak, M.; Ptak, M. Surfactin/iturin A interactions may explain the synergistic effect of surfactin on the biological properties of iturin A. Biochimie 1992, 2, 1047–1051. [Google Scholar] [CrossRef]
- Tao, Y.; Bie, X.M.; Lv, F.X.; Zhao, H.Z.; Lu, Z.X. Antifungal activity and mechanism of fengycin in the presence and absence of commercial surfactin against Rhizopus stolonifer. J. Microbiol. 2011, 2, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Arrebola, E.; Jacobs, R.; Korsten, L. Iturin A is the principal inhibitor in the biocontrol activity of Bacillus amyloliquefaciens PPCB004 against postharvest fungal pathogens. J. Appl. Microbiol. 2010, 2, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Pathak, K.V.; Keharia, H.; Gupta, K.; Thakur, S.S.; Balaram, P. Lipopeptides from the banyan endophyte, Bacillus subtilis K1: Mass spectrometric characterization of a library of fengycins. J. Am. Soc. Mass Spectrom. 2012, 2, 1716–1728. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.L.; Xu, Y.; Straight, P.; Dorrestein, P.C. Translating metabolic exchange with imaging mass spectrometry. Nat. Chem. Biol. 2009, 2, 885–887. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.T.; Yang, Y.L.; Xu, Y.; Lamsa, A.; Haste, N.M.; Yang, J.Y.; Ng, J.; Gonzalez, D.; Ellermeier, C.D.; Straight, P.D.; et al. Imaging mass spectrometry of intraspecies metabolic exchange revealed the cannibalistic factors of Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2010, 2, 16286–16290. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, D.J.; Xu, Y.; Yang, Y.L.; Esquenazi, E.; Liu, W.T.; Edlund, A.; Duong, T.; Du, L.; Molnár, I.; Gerwick, W.H.; et al. Observing the invisible through imaging mass spectrometry, a window into the metabolic exchange patterns of microbes. J. Proteom. 2012, 2, 5069–5076. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Sanchez, L.M.; Rath, C.M.; Liu, X.; Boudreau, P.D.; Bruns, N.; Glukhov, E.; Wodtke, A.; de Felicio, R.; Fenner, A.; et al. Molecular networking as a dereplication Strategy. J. Nat. Prod. 2013, 2, 1686–1699. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, H.; Raoudha, J.; Imen, L.; Ons, M.; Mohamed, D.; Moncef, N. GC/MS and LC/MS analysis, and antioxidant and antimicrobial activities of various solvent extracts from Mirabilis jalapa tubers. Proc. Biochem. 2010, 2, 1486–1493. [Google Scholar]
- Hoefler, B.C.; Gorzelnik, K.V.; Yang, J.Y.; Hendricks, N.; Dorrestein, P.C.; Straight, P.D. Enzymatic resistance to the lipopeptide surfactin as identified through imaging mass spectrometry of bacterial competition. Proc. Natl. Acad. Sci. USA 2012, 2, 13082–13087. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.Y.; Bie, X.M.; Lu, Z.X.; Lv, F.X.; Tao, Y.; Qu, X.X. Effects of fengycin from Bacillus subtilis fmbJ on apoptosis and necrosis in Rhizopus stolonifer. J. Microbiol. 2014, 2, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, J.T.; Richard, A.W. Under Pressure: Investigating the biology of plant infection by Magnaporthe oryza. Nat. Rev. Microbiol. 2009, 2, 185–195. [Google Scholar]
- Sample Availability: Not available.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Molecular Formula | tR (min) | [M + H]+ | [M + Na]+ | Observed [M + H]+ | Error (ppm) |
|---|---|---|---|---|---|---|
| Surfactin group | ||||||
| C-12 Surfactin # | C49H85N7O13 | 5.91 | 980.6284 | 1002.6104 | 980.6276 | −0.82 |
| C-13 Surfactin # | C50H87N7O13 | 6.22 | 994.6440 | 1016.6260 | 994.6445 | 0.50 |
| C-13 Surfactin # | C51H89N7O13 | 6.39 | 1008.6597 | 1030.6417 | 1008.6606 | 0.89 |
| C-14 Surfactin # | C52H91N7O13 | 6.65 | 1022.6753 | 1044.6573 | 1022.6745 | −0.78 |
| C-15 Surfactin # | C53H93N7O13 | 7.11 | 1036.6912 | 1058.6732 | 1036.6905 | −0.68 |
| C-14 Surfactin *,# | C52H91N7O14 | 5.66 | 1038.6702 | 1060.6522 | 1038.6667 | −3.37 |
| C-16 Surfactin # | C54H95N7O13 | 7.16 | 1050.7059 | 1072.6879 | 1050.7073 | 1.33 |
| C-15 Surfactin *,# | C53H95N7O14 | 5.41 | 1054.7029 | 1076.6849 | 1054.7013 | −1.52 |
| C-17 Surfactin | C55H97N7O13 | 7.32 | 1064.7223 | 1086.7043 | 1064.7214 | −0.85 |
| C-19 Surfactin | C57H101N7O13 | 7.51 | 1078.7379 | 1100.7199 | 1078.7374 | −0.46 |
| Iturin group | ||||||
| C-11 Iturin # | C45H70N12O14 | 2.92 | 1015.5221 | 1037.5041 | 1015.5179 | −4.14 |
| C-13 Iturin A1 # | C48H74N12O14 | 3.43 | 1043.5526 | 1065.5346 | 1043.5489 | −3.55 |
| C-14 Iturin A2 # | C49H76N12O14 | 3.62 | 1057.5682 | 1079.5502 | 1057.5686 | 0.38 |
| C-16 Iturin A6 # | C50H78N12O14 | 3.96 | 1071.5839 | 1093.5659 | 1071.5805 | −3.17 |
| C-17 Iturin A8 # | C51H80N12O14 | 4.14 | 1085.5995 | 1107.5815 | 1085.5958 | −3.41 |
| Fengycin group | ||||||
| C-14 Fengycin A # | C70H106N12O20 | 4.01 | 1435.7725 | 1457.7545 | 1435.7688 | −2.58 |
| C-16 Fengycin A *,# | C72H110N12O19 | 4.72 | 1447.8088 | 1469.7908 | 1447.8038 | −3.45 |
| C-15 Fengycin A # | C71H108N12O20 | 4.29 | 1449.7881 | 1471.7701 | 1449.7848 | −2.28 |
| C-16 Fengycin A # | C72H110N12O20 | 4.36 | 1463.8038 | 1485.7858 | 1463.8012 | −1.78 |
| C-15 Fengycin B # | C72H110N12O20 | 4.49 | 1477.8194 | 1499.8014 | 1477.8159 | −2.37 |
| C-17 Fengycin D *,# | C73H112N12O19 | 5.05 | 1489.8576 | 1511.8396 | 1489.8540 | −2.42 |
| C-16 Fengycin B # | C74H114N12O20 | 4.52 | 1491.8351 | 1513.8171 | 1491.8308 | −2.88 |
| C-17 Fengycin B # | C75H116N12O20 | 4.67 | 1505.8510 | 1527.8490 | 1505.8460 | −3.12 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, J.-H.; Chen, P.-Y.; Yang, Y.-L.; Kan, S.-C.; Hsieh, F.-C.; Liu, Y.-C. Clarification of the Antagonistic Effect of the Lipopeptides Produced by Bacillus amyloliquefaciens BPD1 against Pyricularia oryzae via In Situ MALDI-TOF IMS Analysis. Molecules 2016, 21, 1670. https://doi.org/10.3390/molecules21121670
Liao J-H, Chen P-Y, Yang Y-L, Kan S-C, Hsieh F-C, Liu Y-C. Clarification of the Antagonistic Effect of the Lipopeptides Produced by Bacillus amyloliquefaciens BPD1 against Pyricularia oryzae via In Situ MALDI-TOF IMS Analysis. Molecules. 2016; 21(12):1670. https://doi.org/10.3390/molecules21121670
Chicago/Turabian StyleLiao, Jen-Hung, Pi-Yu Chen, Yu-Liang Yang, Shu-Chen Kan, Feng-Chia Hsieh, and Yung-Chang Liu. 2016. "Clarification of the Antagonistic Effect of the Lipopeptides Produced by Bacillus amyloliquefaciens BPD1 against Pyricularia oryzae via In Situ MALDI-TOF IMS Analysis" Molecules 21, no. 12: 1670. https://doi.org/10.3390/molecules21121670
APA StyleLiao, J. -H., Chen, P. -Y., Yang, Y. -L., Kan, S. -C., Hsieh, F. -C., & Liu, Y. -C. (2016). Clarification of the Antagonistic Effect of the Lipopeptides Produced by Bacillus amyloliquefaciens BPD1 against Pyricularia oryzae via In Situ MALDI-TOF IMS Analysis. Molecules, 21(12), 1670. https://doi.org/10.3390/molecules21121670