
Impact of Cluster Zone Leaf Removal on Grapes cv. Regent Polyphenol Content by the UPLC-PDA/MS Method

Abstract
:1. Introduction
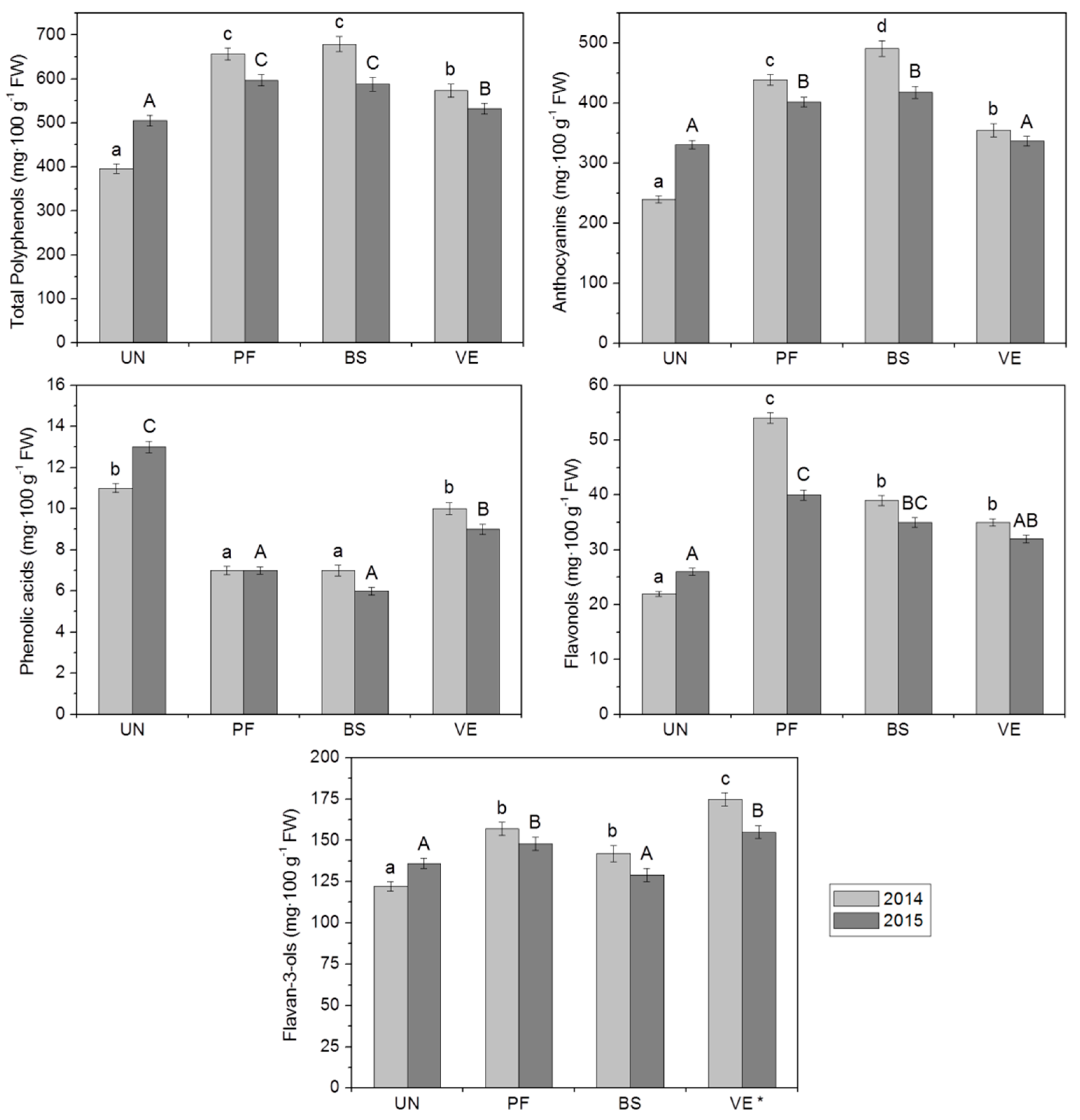
2. Results and Discussion
2.1. Vineyard Weather Data and Treatment Time
2.2. Physico-Chemical Attributes of Berries
2.3. Polyphenol Compounds
2.3.1. Anthocyanins
2.3.2. Phenolic Acids
2.3.3. Flavonols
2.3.4. Flavan-3-ols
3. Materials and Methods
3.1. Characteristics of the Research Area and Experimental Design
3.2. General Grape Parameters
3.3. Polyphenol Content
3.3.1. Reagents and Standards
3.3.2. Extraction Procedure
3.3.3. Identification of Phenolic Compounds with the UPLC-PDA/MS Method
3.4. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of interest
References
- Zhou, K.; Raffoul, J.J. Potential anticancer properties of grape antioxidants. J. Oncol. 2012, 2012, 803294. [Google Scholar] [CrossRef] [PubMed]
- Fahmi, A.I.; El-Shehawi, A.M.; Nagaty, M.A. Antioxadant and antimutagenic activities of Taif grape (Vitis vinifera) cultivars. Am. J. Biochem. Biotechnol. 2013, 9, 102–117. [Google Scholar] [CrossRef]
- Flamini, R.; Mattivi, F.; de Rosso, M.; Arapitsas, P.; Bavaresco, L. Advanced Knowledge of Three Important Classes of Grape Phenolics: Anthocyanins, Stilbenes and Flavonols. Int. J. Mol. Sci. 2013, 14, 19651–19669. [Google Scholar] [CrossRef] [PubMed]
- Xia, E.-Q.; Deng, G.-F.; Guo, Y.-J.; Li, H.-B. Biological Activities of Polyphenols from Grapes. Int. J. Mol. Sci. 2010, 11, 622–646. [Google Scholar] [CrossRef] [PubMed]
- Toaldo, I.M.; van Camp, J.; Gonzales, G.B.; Kamiloglu, S.; Bordignon-Luiz, M.T.; Smagghe, G.; Raes, K.; Capanoglu, E.; Grootaert, C. Resveratrol improves TNF-α-induced endothelial dysfunction in a coculture model of a Caco-2 with an endothelial cell line. J. Nutr. Biochem. 2016, 36, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, J.; Dwyer, J. Flavonoids: Dietary occurrence and biochemical activity. Nutr. Res. 1998, 18, 1995–2018. [Google Scholar] [CrossRef]
- Chedea, V.S.; Braicu, C.; Socaciu, C. Antioxidant/prooxidant activity of a polyphenolic grape seed extract. Food Chem. 2010, 121, 132–139. [Google Scholar] [CrossRef]
- He, J.; Giusti, M.M. Anthocyanins: Natural colorants with health-promoting properties. Annu. Rev. Food Sci. Technol. 2010, 1, 163–187. [Google Scholar] [CrossRef] [PubMed]
- Fenoll, J.; Manso, A.; Hellin, P.; Ruiz, L.; Flores, P. Changes in the aromatic composition of the Vitis vinifera grape Muscat Hamburg during ripening. Food Chem. 2009, 114, 420–428. [Google Scholar] [CrossRef]
- Lutz, M.; Jorquera, K.; Cancino, B.; Ruby, R.; Henriquez, C. Phenolics and antioxidant capacity of table grape (Vitis vinifera L.) cultivars grown in Chile. J. Food Sci. 2011, 76, 1088–1093. [Google Scholar] [CrossRef] [PubMed]
- Koundouras, S.; Marinos, V.; Gkoulioti, A.; Kotseridis, Y.; van Leeuwen, C. Influence of vineyard location and vine water status on fruit maturation of nonirrigated cv. Agiorgitiko (Vitis vinifera L.), Effects on wine phenolic and aroma components. J. Agric. Food Chem. 2006, 54, 5077–5086. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, N.; Guan, L.; Dai, Z.W.; Wu, B.H.; Lauvergeat, V.; Gomès, E.; Li, S.H.; Godoy, F.; Arce-Johnson, P.; Delrot, S. Berry ripening: Recently heard through the grapevine. J. Exp. Bot. 2013, 65, 4543–4559. [Google Scholar] [CrossRef] [PubMed]
- Rolle, L.; Rio Segade, S.; Torchio, F.; Giacosa, S.; Cagnasso, E.; Marengo, F.; Gerbi, V. Influence of grape density and harvest date on changes in phenolic composition, phenol extractability indices, and instrumental texture properties during ripening. J. Agric. Food Chem. 2011, 59, 8796–8805. [Google Scholar] [CrossRef] [PubMed]
- Jackson, D.I.; Lombard, P.B. Environmental and management practices affecting grape composition and wine quality—A review. Am. J. Enol. Vitic. 1993, 44, 409–430. [Google Scholar]
- Feng, H.; Yuan, F.; Skinkis, P.A.; Qian, M.C. Influence of cluster zone leaf removal on Pinot noir grape chemical and volatile composition. Food Chem. 2015, 173, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Skinkis, P.A. Oregon ‘Pinot noir’ grape anthocyanin enhancement by early leaf removal. Food Chem. 2013, 139, 893–901. [Google Scholar] [CrossRef] [PubMed]
- Palliotti, A.; Gardi, T.; Berrios, J.G.; Civardi, S.; Poni, S. Early source limitation as a tool for yield control and wine quality improvement in a high-yielding red Vitis vinifera L. cultivar. Scientia Hortic. 2012, 145, 10–16. [Google Scholar] [CrossRef]
- Nicolosi, E.; Continella, A.; Gentile, A.; Cicala, A.; Ferlito, F. Influence of early leaf removal on autochthonous and international grapevines in Sicily. Scientia Hortic. 2012, 146, 1–6. [Google Scholar] [CrossRef]
- Nicholas, K.A.; Matthews, M.A.; Lobell, D.B.; Willits, N.H.; Field, C.B. Effect of vineyard-scale climate variability on Pinot noir phenolic composition. Agric. Meteorol. 2011, 151, 1556–1567. [Google Scholar] [CrossRef]
- Pogoda.interia.pl. Available online: https://pogoda.interia.pl/archiwum-pogody-01-04-2014,cId,5632 (accessed on 14 May 2016).
- Schernewski, G. Adaptation to Climate Change: Viniculture and Tourism at the Baltic Coast. In Global Change and Baltic Coastal Zones; Schernewski, G., Hofstede, J., Neumann, T., Eds.; Springer Science Business Media B.V.: Dordrecht, Holland; Heidelberg, Germany; London, UK; New York, NY, USA, 2011; Volume 1, pp. 233–247. [Google Scholar]
- Baiano, A.; de Gianni, A.; Previtali, M.A.; del Nobile, M.A.; Novello, V.; de Palma, L. Effects of defoliation on quality attributes of Nero di Troia (Vitis vinifera L.) grape and wine. Food Res. Int. 2015, 75, 260–269. [Google Scholar] [CrossRef]
- Intrigliolo, D.S.; Llacer, E.; Revert, J.; Esteve, M.D.; Climent, M.D.; Palau, D.; Gómez, I. Early defoliation reduces cluster compactness and improves grape composition in Mandó, an autochthonous cultivar of Vitis vinifera from southeastern Spain. Scientia Hortic. 2014, 167, 71–75. [Google Scholar] [CrossRef]
- Kotseridis, Y.; Georgiadou, A.; Tikos, P.; Kallithraka, S.; Koundouras, S. Effects of severity of post-flowering leaf removal on berry growth and composition of three red Vitis vinifera L. cultivars grown under semiarid conditions. J. Agric. Food Chem. 2012, 60, 6000–6010. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, R.F.; Liazid, A.; Palma, M.; Puertas, B.; González-Barrio, R.; Gil-Izquierdo, Á.; García-Barroso, C.; Cantos-Villar, E. Phenolic characterisation of red grapes autochthonous to Andalusia. Food Chem. 2009, 112, 949–955. [Google Scholar] [CrossRef]
- Koundouras, S.; Tsialtas, I.T.; Zioziou, E.; Nikolaou, N. Rootstock effects on the adaptive strategies of grapevine (Vitis vinifera L. cv. Cabernet-Sauvignon) under contrasting water status: Leaf physiological and structural responses. Agric. Ecosyst. Environ. 2008, 128, 86–96. [Google Scholar] [CrossRef]
- Yamane, T.; Jeong, S.T.; Goto-Yamamoto, N.; Koshita, Y.; Kobayashi, S. Effects of temperature on anthocyanin biosynthesis in grape berry skins. Am. J. Enol. Vitic. 2006, 57, 54–59. [Google Scholar]
- Mori, K.; Goto-Yamamoto, N.; Kitayama, M.; Hashizume, K. Loss of anthocyanins in red-wine grape under high temperature. J. Exp. Bot. 2007, 58, 1935–1945. [Google Scholar] [CrossRef] [PubMed]
- Goto-Yamamoto, N.; Mori, K.; Numata, M.; Koyama, K.; Kitayama, M. Effects of temperature and water regimes on flavonoid contents and composition in the skin of red-wine grapes. J. Int. Sci. Vigne Vin 2009, 43, 75–80. [Google Scholar]
- Sternad Lemut, M.; Trost, K.; Sivilotti, P.; Arapitsas, P.; Vrhovsek, U. Early versus late leaf removal strategies for Pinot Noir (Vitis vinifera L.): Effect on colour-related phenolics in young wines following alcoholic fermentation. J. Sci. Food Agric. 2013, 93, 3670–3681. [Google Scholar] [CrossRef] [PubMed]
- Balík, J.; Kumšta, M.; Rop, O. Comparison of anthocyanins present in grapes of Vitis vinifera L. varieties and interspecific hybrids grown in the Czech Republic. Chem. Pap. 2013, 67, 1285–1292. [Google Scholar] [CrossRef]
- Lemut, M.S.; Trost, K.; Sivilotti, P.; Vrhovsek, U. Pinot Noir grape colour related phenolics as affected by leaf removal treatments in the Vipava Valley. J. Food Compos. Anal. 2011, 24, 777–784. [Google Scholar] [CrossRef]
- Nicoletti, I.; Bellincontro, A.; de Rossi, A.; de Sanctis, F.; Tiberi, D.; Pietromarchi, P.; Botondi, R.; Corradini, D.; Mencarelli, F. Postharvest dehydration of Nebbiolo grapes grown at altitude is affected by time of defoliation. Aust. J. Grape Wine Res. 2013, 19, 358–368. [Google Scholar] [CrossRef]
- Bubola, M.; Peršurić, Đ.; Kovačević Ganić, K.; Karoglan, M.; Kozina, B. Effects of fruit zone leaf removal on the concentrations of phenolic and organic acids in Istrian Malvasia grape juice and wine. Food Technol. Biotech. 2012, 50, 159–166. [Google Scholar]
- Ehrhardt, C.; Arapitsas, P.; Stefanini, M.; Flick, G.; Mattivi, F. Analysis of the phenolic composition of fungus-resistant grape varieties cultivated in Italy and Germany using UHPLC-MS/MS. J. Mass Spectrom. 2014, 49, 860–869. [Google Scholar] [CrossRef] [PubMed]
- Mattivi, F.; Guzzon, R.; Vrhovsek, U.; Stefanini, M.; Velasco, R. Metabolite Profiling of Grape: Flavonols and Anthocyanins. J. Agric. Food Chem. 2006, 54, 7692–7702. [Google Scholar] [CrossRef] [PubMed]
- Spayd, S.E.; Tarara, J.M.; Mee, D.L.; Ferguson, J.C. Separation of sunlight and temperature effects on the composition of Vitis vinifera cv. Merlot berries. Am. J. Enol. Vitic. 2002, 53, 171–182. [Google Scholar]
- Lisek, J. Yielding and healthiness of selected grape cultivars for processing in central Poland. J. Fruit Ornam. Plant Res. 2010, 18, 265–272. [Google Scholar]
- Oszmiański, J.; Kolniak-Ostek, J.; Wojdyło, A. Characterization and content of flavonol derivatives of Allium ursinum L. plant. J. Agric. Food Chem. 2013, 61, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not Available.


{kind=link}
| Year | Month | |||||||
|---|---|---|---|---|---|---|---|---|
| April | May | June | July | August | September | October | ||
| Average Temperature (°C) | Mean | |||||||
| 2014 | 10.8 | 13.4 | 16.3 | 21.3 | 17.5 | 15.4 | 11.8 | 15.2 |
| 2015 | 8.7 | 12.5 | 15.6 | 18.6 | 21.1 | 14.1 | 13.7 | 14.9 |
| 1951–2012 | 8.0 | 13.0 | 16.4 | 18.2 | 17.6 | 13.8 | 9.2 | 13.7 |
| Rainfall (mm) | Total | |||||||
| 2014 | 47.5 | 85.3 | 26.5 | 70.8 | 104.6 | 80.9 | 32.8 | 448 |
| 2015 | 29.0 | 48.0 | 32.8 | 62.0 | 14.7 | 34.4 | 22.1 | 242 |
| 1951–2012 | 39.7 | 62.9 | 48.2 | 69.6 | 74.2 | 58.7 | 37.3 | 391 |
| Number of Days with Temperature Over 30 °C | Total | |||||||
| 2014 | - | - | 1 | 8 | - | 1 | - | 10 |
| 2015 | - | - | - | 3 | 11 | - | - | 14 |
| Sun Hours | Total | |||||||
| 2014 | 210 | 213 | 189 | 224 | 123 | 133 | 99 | 1191 |
| 2015 | 136 | 161 | 159 | 197 | 278 | 126 | 131 | 1188 |
| Cloudiness (%) | Mean | |||||||
| 2014 | 29 | 35 | 32 | 21 | 31 | 32 | 37 | 31 |
| 2015 | 34 | 31 | 37 | 31 | 17 | 29 | 36 | 31 |
| Year | Leaf Removal Treatments * | ||||
|---|---|---|---|---|---|
| UN | PF | BS | VE | Mean | |
| Cluster Weight (g) | |||||
| 2014 | 95 ± 17 a | 106 ± 20 ab | 87 ± 14 a | 127 ± 19 bc | 104 A |
| 2015 | 125 ± 21 bc | 133 ± 23 bc | 146 ± 18 c | 151 ± 24 c | 139 B |
| mean | 110 A | 120 A | 116 A | 139 A | |
| Weight of 100 Berries (g) | |||||
| 2014 | 177 ± 9 bcd | 200 ± 11 de | 193 ± 11 cde | 215 ± 13 e | 196 B |
| 2015 | 161 ± 8 ab | 148 ± 7 a | 165 ± 10 abc | 158 ± 8 ab | 158 A |
| mean | 169 A | 174 A | 179 A | 186 A | |
| Total Soluble solid (°Brix) | |||||
| 2014 | 19.5 ± 0.24 a | 20.4 ± 0.22 b | 19.4 ± 0.21 a | 20.2 ± 0.17 a | 19.9 A |
| 2015 | 22.3 ±0.27 c | 21.9 ± 0.19 c | 22.4 ± 0.21 c | 20.7 ± 0.25 b | 21.8 B |
| mean | 20.9 AB | 21.2 B | 20.9 AB | 20.5 A | |
| pH | |||||
| 2014 | 3.88 ± 0.07 c | 3.82 ± 0.06 bc | 3.77 ± 0.09 bc | 3.77 ± 0.05 bc | 3.80 B |
| 2015 | 3.24 ± 0.04 a | 3.23 ± 0.05 a | 3.29 ± 0.04 a | 3.23 ± 0.03 a | 3.26 A |
| mean | 3.56 A | 3.53 A | 3.53 A | 3.50 A | |
| Titratable acidity (g/L) | |||||
| 2014 | 7.55 ± 0.08 b | 7.36 ± 0.08 ab | 7.30 ± 0.09 a | 7.42 ± 0.07 ab | 7.41 A |
| 2015 | 8.65 ± 0.11 d | 8.24 ± 0.09 c | 8.40 ± 0.07 c | 8.63 ± 0.07 cd | 8.48 B |
| mean | 8.10 B | 7.80 A | 7.85 AB | 8.03 B | |
| Compounds | Rt (min) | λmax (nm) | (M − H)+ (m/z) | (MS/MS) (m/z) |
|---|---|---|---|---|
| Gallic acid | 1.08 | 276 | 169 | 125 |
| cis-Caftaric acid | 2.25 | 328 | 311 | 179; 149 |
| trans-Caftaric acid | 2.58 | 328 | 311 | 179; 149 |
| cis-Coutaric acid | 2.63 | 310 | 295 | 163; 149 |
| Procyanidin B1 | 2.76 | 278 | 577 | 289 |
| trans-Coutaric acid | 3.04 | 310 | 295 | 163; 149 |
| Procyanidin B3 | 3.12 | 278 | 577 | 289 |
| (+)-Catechin | 3.19 | 276 | 289 | 245; 205 |
| cis-Fertaric acid | 3.57 | 328 | 325 | 193 |
| Procyanidin B2 | 3.98 | 278 | 577 | 289 |
| trans-Fertaric acid | 4.00 | 328 | 325 | 193 |
| Delphinidin-3-O-glucoside | 4.15 | 526 | 465+ | 303 |
| (−)-Epicatechin | 4.69 | 276 | 289 | 245; 205 |
| Cyanidin-3-O-glucoside | 4.85 | 516 | 449+ | 287 |
| Petunidin-3-O-glucoside | 5.34 | 527 | 479+ | 317 |
| Peonidin-3-O-glucoside | 6.06 | 516 | 463+ | 301 |
| Malvidin-3-O-glucoside | 6.42 | 527 | 493+ | 331 |
| Myricetin-3-O-glucoside | 6.51 | 343 | 479 | 317 |
| Delphinidin-3-O-acethyl-glucoside | 6.90 | 529 | 507+ | 303 |
| Quercetin-3-O-rutinoside | 7.11 | 352 | 609 | 463; 301 |
| Quercetin-3-O-glucoside | 7.26 | 352 | 463 | 301 |
| Quercetin-3-O-glucuronide | 7.37 | 352 | 477 | 301 |
| Cyanidin-3-O-acethyl-glucoside | 7.70 | 517 | 491+ | 287 |
| Petunidin-3-O-acethyl-glucoside | 8.03 | 529 | 521+ | 317 |
| Kaempferol-3-O-glucoside | 8.04 | 350 | 447 | 285 |
| Peonidin-3-O-acethyl-glucoside | 8.45 | 517 | 505+ | 301 |
| Isorhamnetin-3-O-glucoside | 8.51 | 354 | 477 | 315 |
| Malvidin-3-O-acethyl-glucoside | 8.89 | 529 | 535+ | 331 |
| Delphinidin-3-O-(6-p-coumaroyl)-glucoside | 9.09 | 527 | 611+ | 303 |
| Cyanidin-3-O-(6-p-coumaroyl)-glucoside | 9.66 | 520 | 595+ | 287 |
| Petunidin-3-O-(6-p-coumaroyl)-glucoside | 9.89 | 527 | 625+ | 317 |
| Peonidin-3-O-(6-p-coumaroyl)-glucoside | 10.70 | 525 | 609+ | 301 |
| Malvidin-3-O-(6-p-coumaroyl)-glucoside | 10.84 | 530 | 639+ | 331 |
| Compounds | Leaf Removal Treatments * | |||
|---|---|---|---|---|
| UN | PF | BS | VE | |
| Cyanidin-3-O-glucoside | 45.32 ± 2.91 a | 78.97 ± 4.57 b | 72.99 ± 5.02 b | 54.78 ± 3.14 a |
| Delphinidin-3-O-glucoside | 65.85 ± 2.06 a | 83.82 ± 3.24 bc | 93.49 ± 3.11 c | 74.49 ± 2.48 ab |
| Malvidin-3-O-glucoside | 51.99 ± 1.79 a | 75.71 ± 2.35 b | 91.05 ± 2.52 c | 61.52 ± 1.90 a |
| Peonidin-3-O-glucoside | 42.31 ± 1.33 a | 68.46 ± 2.15 bc | 75.03 ± 1.98 c | 61.18 ± 1.76 b |
| Petunidin-3-O-glucoside | 47.01 ± 1.87 a | 81.52 ± 1.54 c | 89.56 ± 1.72 c | 64.54 ± 1.90 b |
| Cyanidin-3-O-acethyl-glucoside | 2.56 ± 0.19 b | 1.01 ± 0.13 a | 1.11 ± 0.11 a | 0.82 ± 0.10 a |
| Delphinidin-3-O-acethyl-glucoside | 4.56 ± 0.18 b | 1.16 ± 0.12 a | 1.16 ± 0.10 a | 1.04 ± 0.13 a |
| Malvidin-3-O-acethyl-glucoside | 2.70 ± 0.0.9 a | 4.05 ± 0.10 c | 3.89 ± 0.09 bc | 3.48 ± 0.08 b |
| Peonidin-3-O-acethyl-glucoside | 0.22 ± 0.01 a | 0.67 ± 0.02 b | 0.75 ± 0.02 b | 0.59 ± 0.02 b |
| Petunidin-3-O-acethyl-glucoside | 3.51 ± 0.21 c | 1.10 ± 0.04 b | 0.89 ± 0.03 a | 1.17 ± 0.03 b |
| Cyanidin-3-O-(6-p-coumaroyl)-glucoside | 3.09 ± 0.10 a | 5.46 ± 0.10 c | 5.77 ± 0.11 c | 4.90 ± 0.08 b |
| Delphinidin-3-O-(6-p-coumaroyl)-glucoside | 6.27 ± 0.28 b | 1.24 ± 0.09 a | 1.49 ± 0.11 a | 1.34 ± 0.09 a |
| Malvidin-3-O-(6-p-coumaroyl)-glucoside | 4.43 ± 0.13 a | 6.94 ± 0.18 b | 7.06 ±0.17 b | 6.67 ± 0.15 b |
| Peonidin-3-O-(6-p-coumaroyl)-glucoside | 2.38 ± 0.07 a | 4.47 ± 0.12 c | 4.36 ± 0.10 bc | 4.11 ± 0.11 b |
| Petunidin-3-O-(6-p-coumaroyl)-glucoside | 3.08 ± 0.09 a | 5.67 ± 0.08 b | 5.96 ± 0.08 b | 5.21 ± 0.10 b |
| Anthocyanins (%) | 285.28 A (63) | 420.25 C (67) | 454.56 C (72) | 345.84 B (62) |
| cis-Caftaric acid | 1.61 ± 0.07 c | 0.45 ± 0.02 a | 0.97 ± 0.03 b | 1.42 ± 0.03 c |
| trans-Caftaric acid | 5.25 ± 0.16 d | 1.66 ± 0.07 b | 0.94 ± 0.04 a | 2.30 ± 0.08 c |
| cis-Coutaric acid | 1.07 ± 0.05 a | 2.21 ± 0.08 b | 2.01 ± 0.13 b | 2.46 ± 0.10 b |
| trans-Coutaric acid | 0.51 ± 0.02 a | 0.55 ± 0.01 a | 0.61 ± 0.02 a | 0.50 ± 0.02 a |
| cis-Fertaric acid | 0.97 ± 0.05 c | 0.34 ± 0.02 a | 0.25 ± 0.01 a | 0.73 ± 0.03 b |
| trans-Fertaric acid | 0.39 ± 0.02 a | 0.55 ± 0.01 b | 0.54 ± 0.03 b | 0.45 ± 0.02 ab |
| Gallic acid | 2.19 ± 0.09 b | 1.18 ± 0.06 a | 1.03 ± 0.04 a | 1.26 ± 0.05 a |
| Phenolic acids (%) | 11.98 C (3) | 6.94 A (1) | 6.35 A (1) | 9.12 B (2) |
| Myricetin-3-O-glucoside | 3.50 ± 0.15 a | 4.54 ± 0.21 b | 6.23 ± 0.30 c | 5.43 ± 0.26 bc |
| Quercetin-3-O-rutinoside | 3.39 ± 0.17 ab | 5.71 ± 0.19 c | 3.73 ± 0.22 b | 3.07 ± 0.14 a |
| Quercetin-3-O-glucoside | 6.95 ± 0.28 a | 15.09 ± 0.55 c | 11.79 ± 0.37 b | 11.31 ± 0.34 b |
| Quercetin-3-O-glucuronide | 8.44 ± 0.29 a | 16.74 ± 0.41 c | 12.05 ± 0.34 b | 11.02 ± 0.28 b |
| Kaempferol-3-O-glucoside | 0.45 ± 0.03 ab | 0.97 ± 0.04 c | 0.56 ± 0.04 b | 0.44 ± 0.02 a |
| Isorhamnetin-3-O-glucoside | 1.25 ± 0.09 a | 3.85 ± 0.12 c | 2.58 ± 0.11 b | 2.53 ± 0.10 b |
| Flavonols (%) | 23.98A (5) | 46.90 C (7) | 36.94 B (6) | 33.80 B (6) |
| Procyanidin B1 | 10.87 ± 0.48 a | 13.21 ± 0.53 b | 10.70 ± 0.44 a | 17.47 ± 0.58 c |
| Procyanidin B2 | 31.19 ± 1.44 b | 23.76 ± 1.31 a | 24.16 ± 1.35 a | 26.10 ± 1.48 a |
| Procyanidin B3 | 5.64 ± 0.21 b | 4.45 ± 0.14 a | 5.81 ± 0.17 b | 9.15 ± 0.24 c |
| (+)-Catechin | 39.17 ± 1.56 a | 77.42 ± 2.93 c | 64.83 ± 2.38 b | 71.32 ± 3.02 bc |
| (−)-Epicatechin | 42.40 ± 2.11 c | 33.85 ± 1.76 a | 30.20 ± 1.95 a | 40.82 ± 1.48 b |
| Flavan-3-ols (%) | 129.27 A (29) | 152.68 B (24) | 135.70 A (21) | 164.86 B (30) |
| Total polyphenols | 450.51 A | 626.77 C | 633.55 C | 553.62 B |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mijowska, K.; Ochmian, I.; Oszmiański, J. Impact of Cluster Zone Leaf Removal on Grapes cv. Regent Polyphenol Content by the UPLC-PDA/MS Method. Molecules 2016, 21, 1688. https://doi.org/10.3390/molecules21121688
Mijowska K, Ochmian I, Oszmiański J. Impact of Cluster Zone Leaf Removal on Grapes cv. Regent Polyphenol Content by the UPLC-PDA/MS Method. Molecules. 2016; 21(12):1688. https://doi.org/10.3390/molecules21121688
Chicago/Turabian StyleMijowska, Kamila, Ireneusz Ochmian, and Jan Oszmiański. 2016. "Impact of Cluster Zone Leaf Removal on Grapes cv. Regent Polyphenol Content by the UPLC-PDA/MS Method" Molecules 21, no. 12: 1688. https://doi.org/10.3390/molecules21121688
APA StyleMijowska, K., Ochmian, I., & Oszmiański, J. (2016). Impact of Cluster Zone Leaf Removal on Grapes cv. Regent Polyphenol Content by the UPLC-PDA/MS Method. Molecules, 21(12), 1688. https://doi.org/10.3390/molecules21121688