
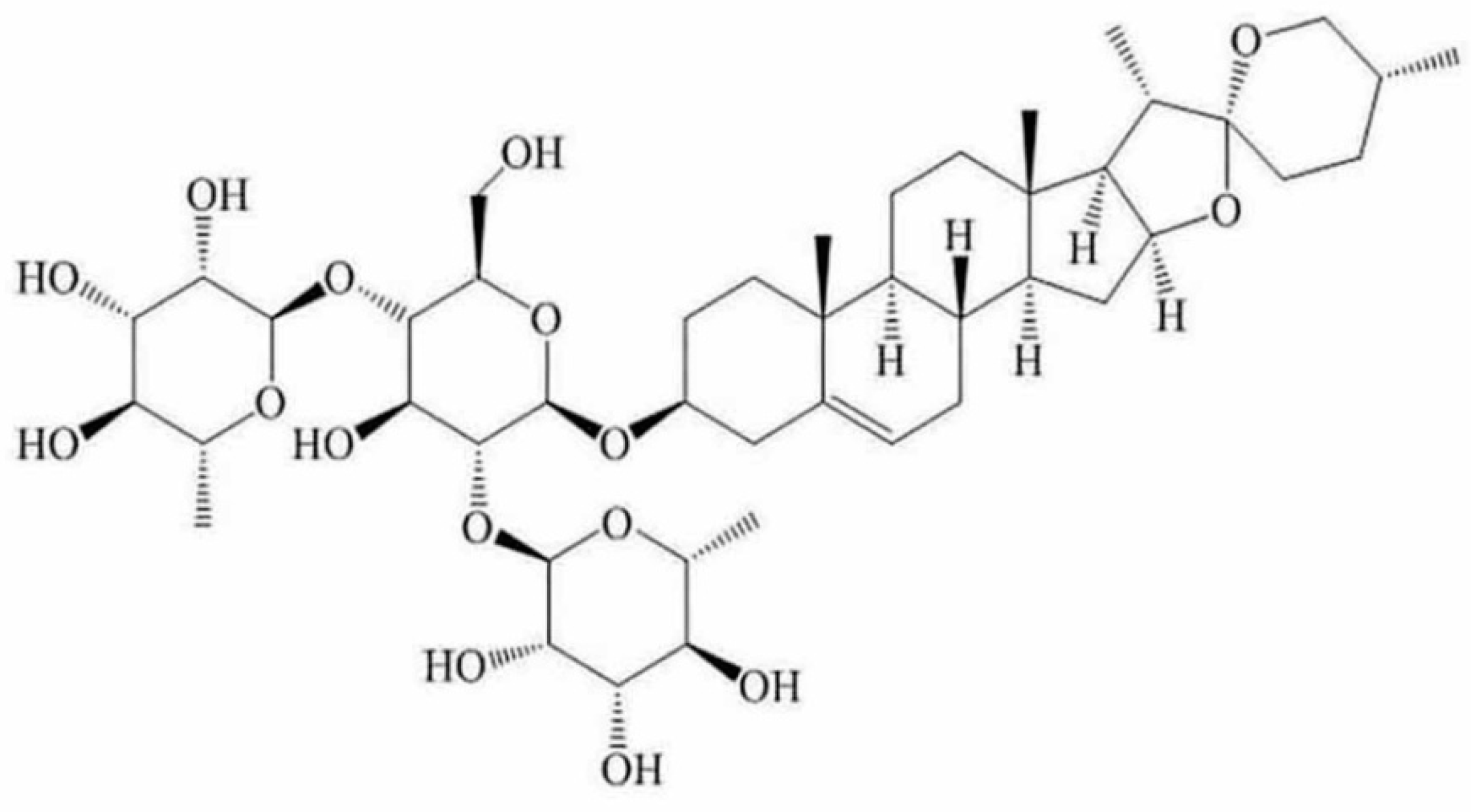
Dioscin Induces Apoptosis in Human Cervical Carcinoma HeLa and SiHa Cells through ROS-Mediated DNA Damage and the Mitochondrial Signaling Pathway


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
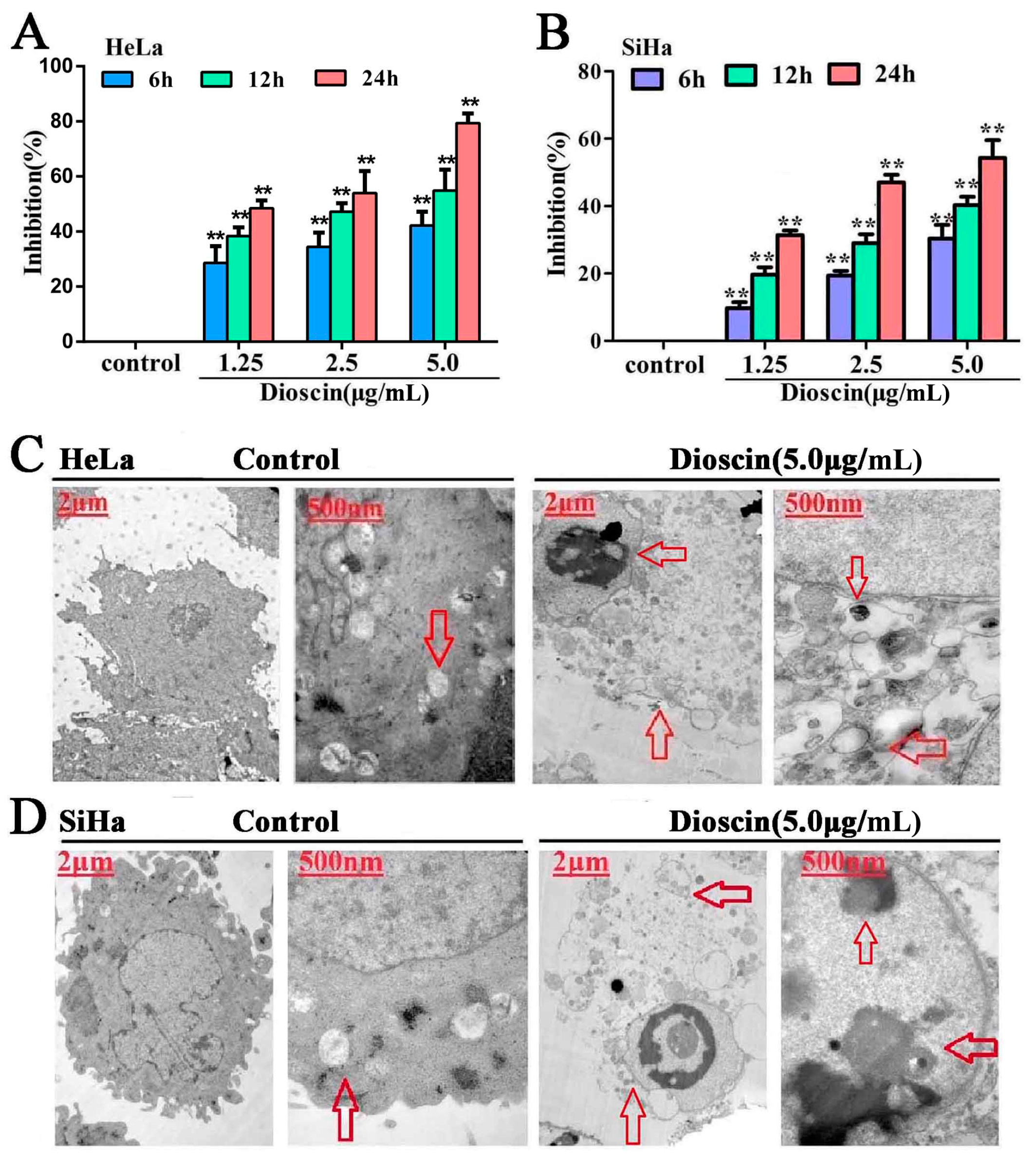
2.1. Dioscin-Inhibited Cell Viability
2.2. Dioscin-Induced Cells Ultrastructure Changes
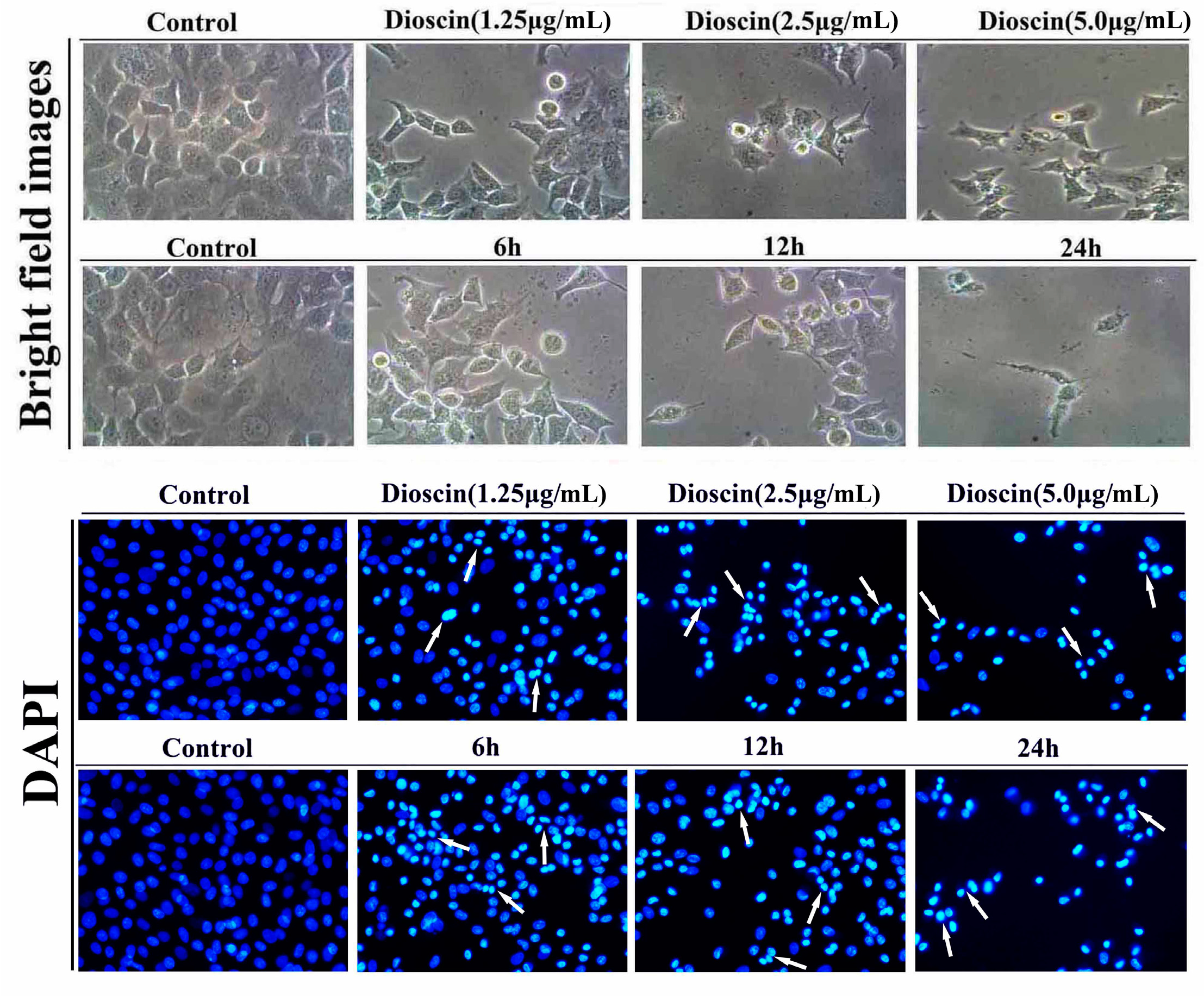
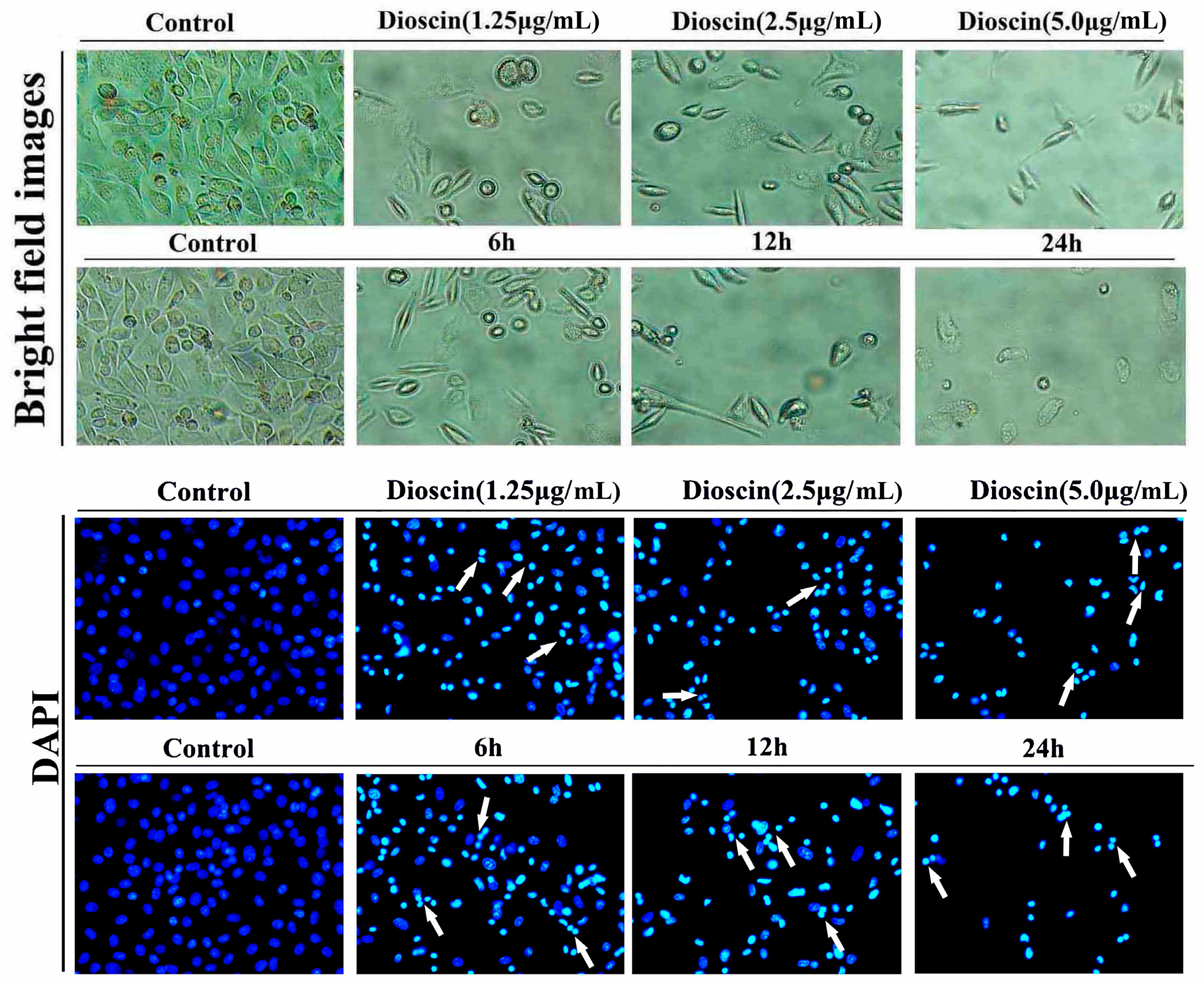
2.3. Dioscin-Induced Morphological Changes
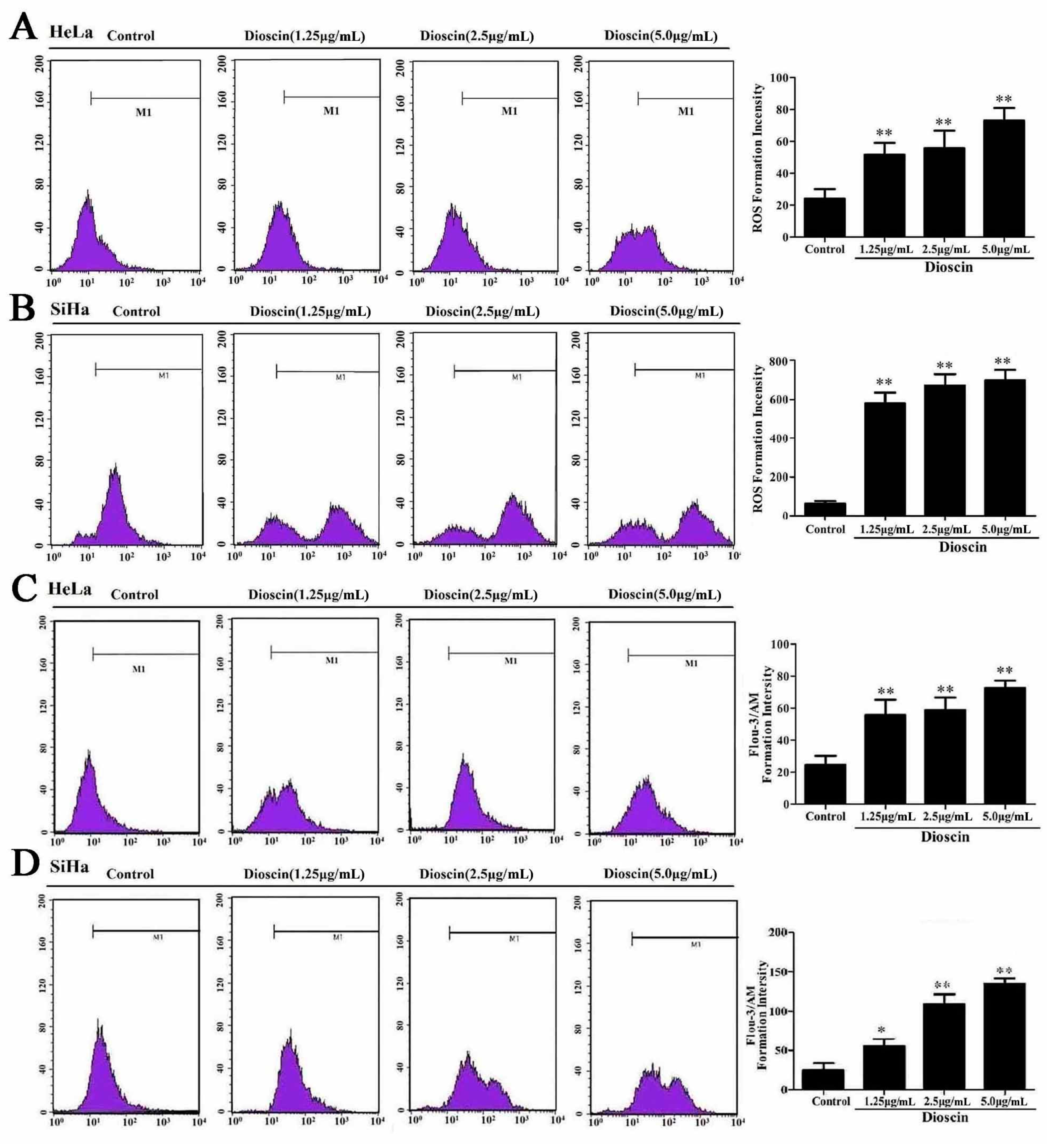
2.4. Effects of Dioscin on ROS Levels
2.5. Dioscin Caused Ca2+ Release in HeLa and SiHa Cells
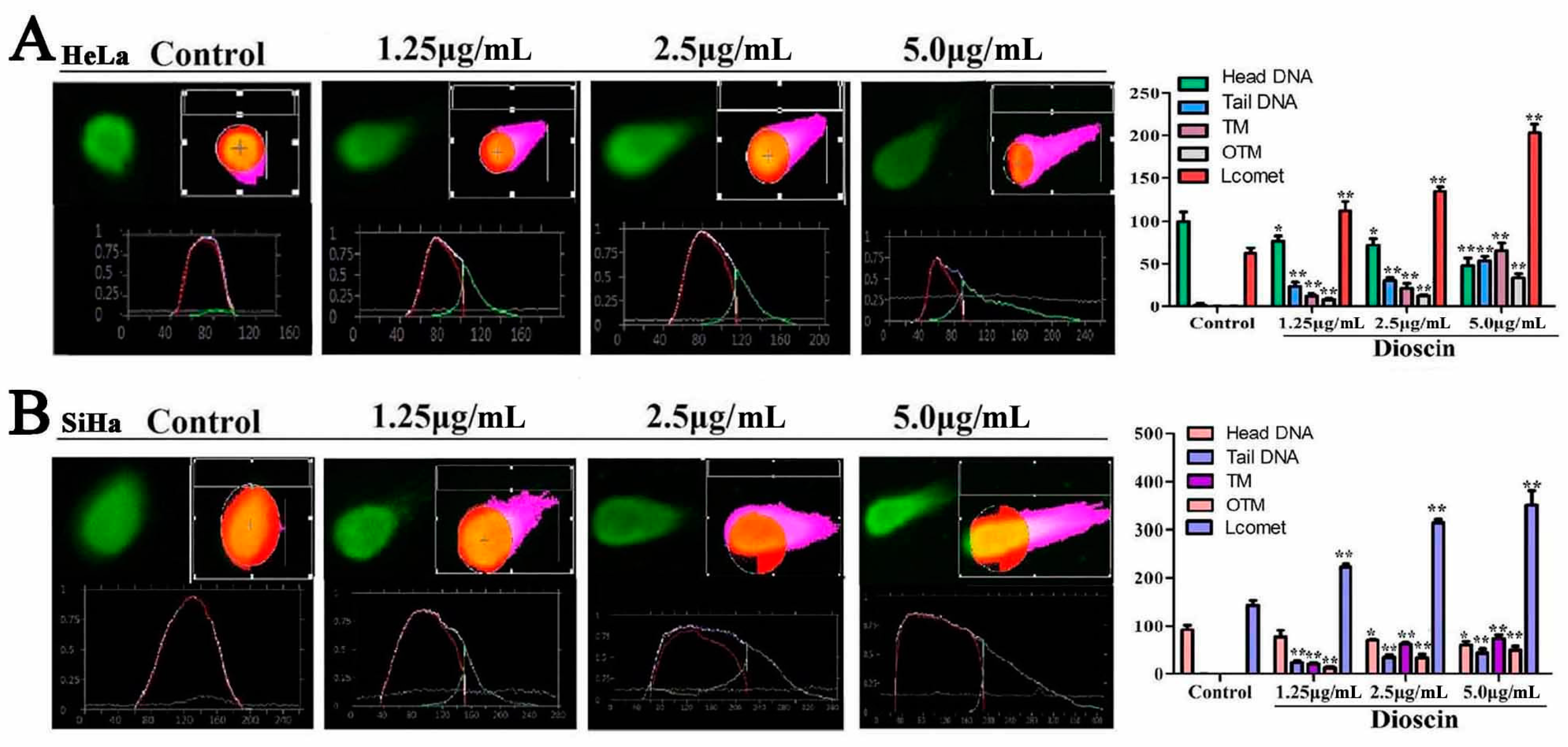
2.6. Dioscin Induced DNA Damage of the HeLa and SaHa Cells
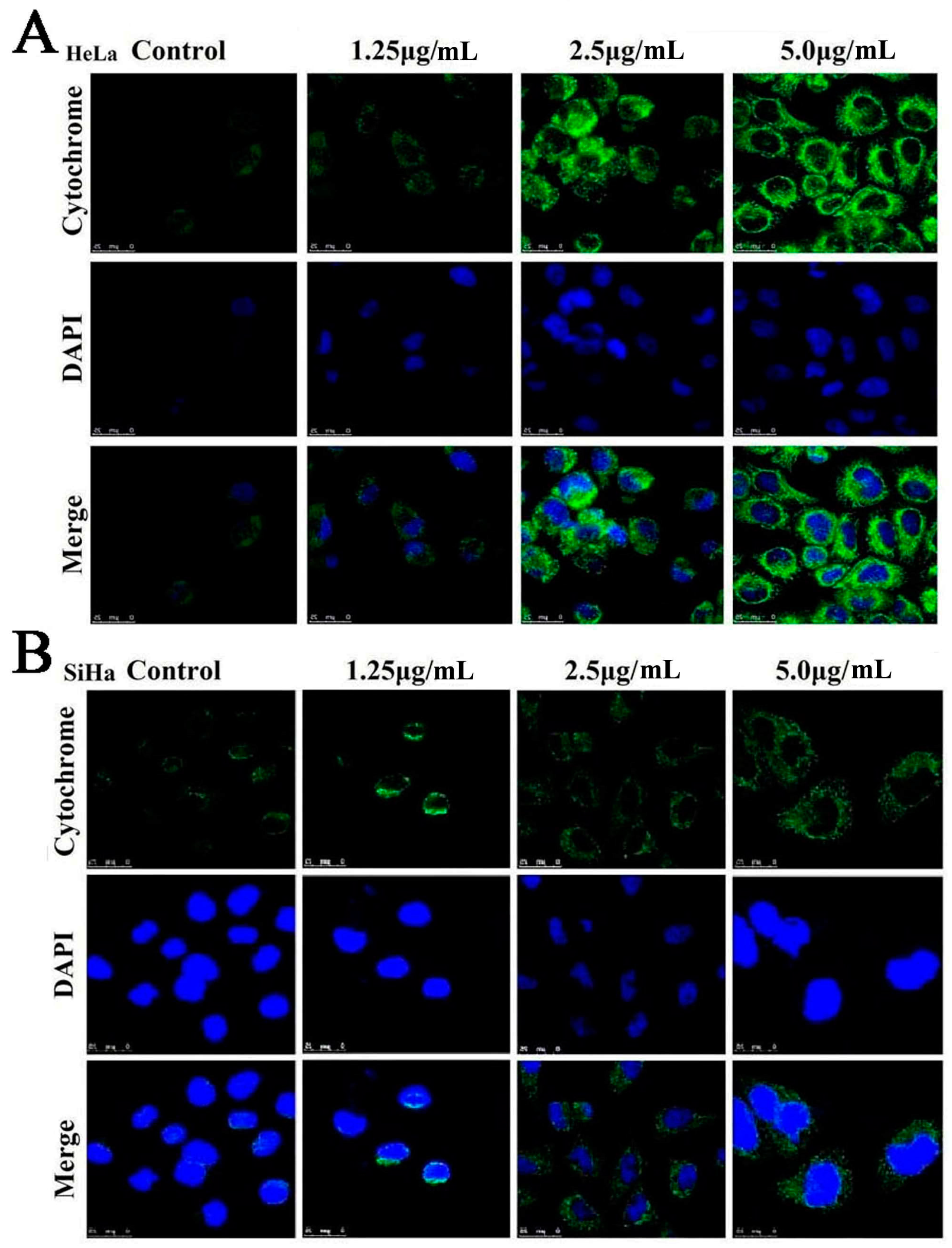
2.7. Dioscin Affects theMitochondrialSignaling Pathway
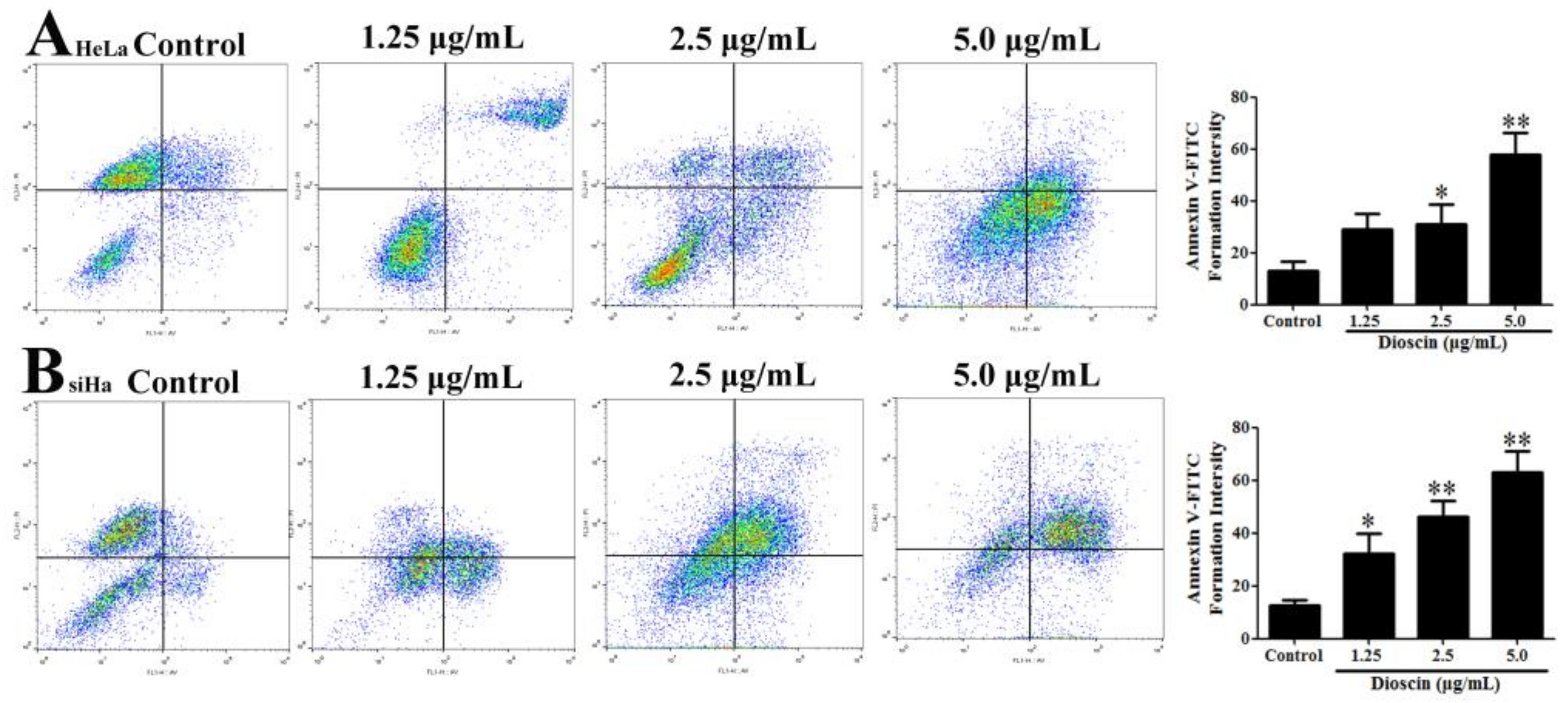
2.8. Dioscin Induced HeLa and SiHa Cell Apoptosis
3. Discussion
4. Materials and Methods
4.1. Materials, Cell Culture and Viability Assay
4.2. Transmission Electron Microscope (TEM) Assay
4.3. DAPI Staining
4.4. Detection of Intracellular ROS Accumulation
4.5. Detection of Cell Apoptosis and Intracellular Ca2+Release
4.6. Single Cell Gel Electrophoresis Assay
4.7. Detection of Cytochrome C Release
4.8. Detection of Cell Apoptosis
4.9. Western Blotting Assay
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Xu, L.; Yin, L.; Qi, Y.; Li, H.; Xu, Y.; Han, X.; Peng, J.; Wan, X. Cytotoxicity of dioscin in human gastric carcinoma cells through death receptor and mitochondrial pathways. J. Appl. Toxicol. 2013, 33, 712–722. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Yuan, Y.; Okumura, Y.; Shinkai, N.; Yamauchi, H. Camptothecin disrupts androgen receptor signaling and suppresses prostate cancer cell growth. Biochem. Biophys. Res. Commun. 2010, 394, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Pathak, N.; Khandelwal, S. Role of oxidative stress and apoptosis in cadmium induced thymic atrophy and splenomegaly in mice. Toxicol. Lett. 2007, 169, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Parent, N.; Winstall, E.; Beauchemin, M.; Paquet, C.; Poirier, G.G.; Bertrand, R. Proteomic analysis of enriched lysosomes at early phase of camptothecin-induced apoptosis in human U-937 cells. J. Proteom. 2009, 72, 960–973. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Fan, J.; Wang, Q.; Ju, D.; Feng, M.; Li, J.; Guan, Z.B.; An, D.; Wang, X.; Ye, L. Diosgenin induces ROS-dependent autophagy and cytotoxicity via mTOR signaling pathway in chronic myeloid leukemia cells. Phytomedicine 2016, 23, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Li, X.; He, X.; Qiu, X.; Jin, X.; Zhao, X.Y.; Xu, R.Z. Polyphyllin I induces cell cycle arrest and apoptosis in human myeloma cells via modulating β-catenin signalling pathway. Eur. J. Haematol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Jia, X.; Bao, J.; Chen, S.; Wang, K.; Zhang, Y.; Li, P.; Wan, J.B.; Su, H.; Wang, Y.; et al. Polyphyllin VII induces apoptosis in HepG2 cells through ROS-mediated mitochondrial dysfunction and MAPK pathways. BMC Complement. Altern. Med. 2016, 16, 58. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Jin, X.; Qian, C.; Ruan, Y.; Jiang, H. Signaling network of OSW-1-induced apoptosis and necroptosis in hepatocellular carcinoma. Mol. Med. Rep. 2013, 7, 1646–1650. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Han, X.; Yu, B. Synthesis of monomethylated dioscin derivatives and their antitumor activities. Carbohydr. Res. 2003, 338, 117–121. [Google Scholar] [CrossRef]
- Ikeda, T.; Ando, J.; Miyazono, A.; Zhu, X.H.; Tsumagari, H.; Nohara, T.; Yokomizo, K.; Uyeda, M. Anti-herpes virus activity of Solanum steroidal glycosides. Biol. Pharm. Bull. 2000, 23, 363–364. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Yin, L.; Xu, L.; Peng, J. Application of proteomic and bioinformatic techniques for studying the hepatoprotective effect of dioscin against CCl(4)-induced liver damage in mice. Planta Med. 2011, 77, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Tao, X.; Wan, X.; Xu, Y.; Xu, L.; Qi, Y.; Yin, L.; Han, X.; Lin, Y.; Peng, J. Dioscin attenuates hepatic ischemia-reperfusion injury in rats through inhibition of oxidative-nitrative stress, inflammation and apoptosis. Transplantation 2014, 98, 604–611. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Xu, Y.; Han, X.; Qi, Y.; Xu, L.; Xu, Y.; Yin, L.; Sun, H.; Liu, K.; Peng, J. Anti-cancer effects of dioscin on three kinds of human lung cancer cell lines through inducing DNA damage and activating mitochondrial signal pathway. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2013, 59, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Liu, M.; Wang, Z.; Ju, Y. Apoptosis induced by dioscin in Hela cells. Biol. Pharm. Bull. 2002, 25, 193–196. [Google Scholar] [CrossRef] [PubMed]
- Franco, E.L. Epidemiology of anogenital warts and cancer. Obstet. Gynecol. Clin. N. Am. 1996, 23, 597–623. [Google Scholar]
- Walboomers, J.M.; Jacobs, M.V.; Manos, M.M.; Bosch, F.X.; Kummer, J.A.; Shah, K.V. Human papillomavirus is a necessary cause of invasive cervical cancer worldwide. J. Pathol. 1999, 189, 12–19. [Google Scholar] [CrossRef]
- Wright, J.D.; Herzog, T.J. Human papillomavirus: emerging trends in detection and management. Curr. Womens Health Rep. 2002, 2, 259–265. [Google Scholar] [PubMed]
- Kim, E.A.; Jang, J.H.; Lee, Y.H.; Sung, E.G.; Song, I.H.; Kim, J.Y.; Kim, S.; Sohn, H.Y.; Lee, T.J. Dioscin induces caspase-independent apoptosis through activation of apoptosis-inducing factor in breast cancer cells. Apoptosis 2014, 19, 1165–1175. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, H.M.; Zhang, X.N.; Xiong, C.M.; Ruan, J.L. Dioscin-induced apoptosis of human LNCaP prostate carcinoma cells through activation of caspase-3 and modulation of Bcl-2 protein family. J. Huazhong Univ. Sci. Technol. Med. Sci. 2014, 34, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Xu, L.; Zheng, L.; Yin, L.; Qi, Y.; Han, X.; Xu, Y.; Peng, J. Potent effects of dioscin against gastric cancer in vitro and in vivo. Phytomedicine 2016, 23, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Kundu, S.; Bhattacharyya, A. Mechanism of cadmium induced apoptosis in the immunocyte. Toxicol. Lett. 2008, 177, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Martin, K.R.; Barrett, J.C. Reactive oxygen species as double-edged swords in cellular processes: Low-dose cell signaling versus high-dose toxicity. Hum. Exp. Toxicol. 2002, 21, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Green, D.R.; Reed, J.C. Mitochondria and apoptosis. Science (New York, N. Y.) 1998, 281, 1309–1312. [Google Scholar] [CrossRef]
- Mallis, R.J.; Buss, J.E.; Thomas, J.A. Oxidative modification of H-ras: S-thiolation and S-nitrosylation of reactive cysteines. Biochem. J. 2001, 355, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Simon, H.U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis Int. J. Programm. Cell Death 2000, 5, 415–418. [Google Scholar] [CrossRef]
- Schumacker, P.T. Reactive oxygen species in cancer cells: live by the sword, die by the sword. Cancer Cell 2006, 10, 175–176. [Google Scholar] [CrossRef] [PubMed]
- Borner, C. The Bcl-2 protein family: Sensors and checkpoints for life-or-death decisions. Mol. Immunol. 2003, 39, 615–647. [Google Scholar] [CrossRef]
- Brenner, C.; Cadiou, H.; Vieira, H.L.; Zamzami, N.; Marzo, I.; Xie, Z.; Leber, B.; Andrews, D.; Duclohier, H.; Reed, J.C.; et al. Bcl-2 and Bax regulate the channel activity of the mitochondrial adenine nucleotide translocator. Oncogene 2000, 19, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Vyssokikh, M.Y.; Zorova, L.; Zorov, D.; HeimLich, G.; Jurgensmeier, J.J.; Brdiczka, D. Bax releases cytochrome c preferentially from a complex between porin and adenine nucleotide translocator. Hexokinase activity suppresses this effect. Mol. Biol. Rep. 2002, 29, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Budihardjo, I.; Zou, H.; Slaughter, C.; Wang, X. Bid, a Bcl2 interacting protein, mediates cytochrome c release from mitochondria in response to activation of cell surface death receptors. Cell 1998, 94, 481–490. [Google Scholar] [CrossRef]
- Huttemann, M.; Pecina, P.; Rainbolt, M.; Sanderson, T.H.; Kagan, V.E.; Samavati, L.; Doan, J.W.; Lee, I. The multiple functions of cytochrome c and their regulation in life and death decisions of the mammalian cell: From respiration to apoptosis. Mitochondrion 2011, 11, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds like HeLa and SiHa cells and dioscin are available from the authors.









© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Tao, X.; Xu, L.; Yin, L.; Qi, Y.; Xu, Y.; Han, X.; Peng, J. Dioscin Induces Apoptosis in Human Cervical Carcinoma HeLa and SiHa Cells through ROS-Mediated DNA Damage and the Mitochondrial Signaling Pathway. Molecules 2016, 21, 730. https://doi.org/10.3390/molecules21060730
Zhao X, Tao X, Xu L, Yin L, Qi Y, Xu Y, Han X, Peng J. Dioscin Induces Apoptosis in Human Cervical Carcinoma HeLa and SiHa Cells through ROS-Mediated DNA Damage and the Mitochondrial Signaling Pathway. Molecules. 2016; 21(6):730. https://doi.org/10.3390/molecules21060730
Chicago/Turabian StyleZhao, Xinwei, Xufeng Tao, Lina Xu, Lianhong Yin, Yan Qi, Youwei Xu, Xu Han, and Jinyong Peng. 2016. "Dioscin Induces Apoptosis in Human Cervical Carcinoma HeLa and SiHa Cells through ROS-Mediated DNA Damage and the Mitochondrial Signaling Pathway" Molecules 21, no. 6: 730. https://doi.org/10.3390/molecules21060730
APA StyleZhao, X., Tao, X., Xu, L., Yin, L., Qi, Y., Xu, Y., Han, X., & Peng, J. (2016). Dioscin Induces Apoptosis in Human Cervical Carcinoma HeLa and SiHa Cells through ROS-Mediated DNA Damage and the Mitochondrial Signaling Pathway. Molecules, 21(6), 730. https://doi.org/10.3390/molecules21060730