
Capsaicin Inhibited Aggressive Phenotypes through Downregulation of Tumor-Associated NADH Oxidase (tNOX) by POU Domain Transcription Factor POU3F2



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
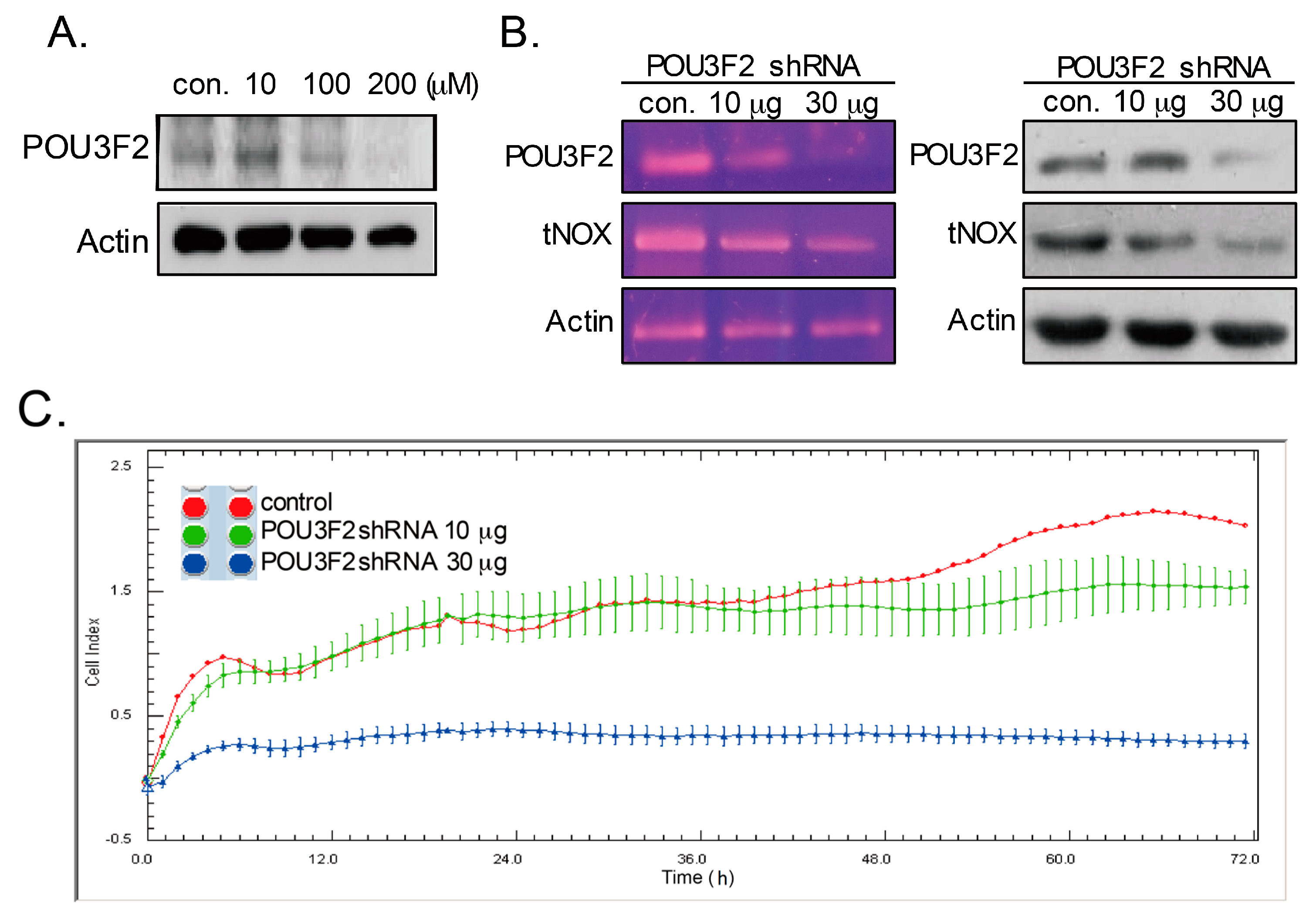
2.1. Capsaicin Suppressed tNOX Expression and Cell Migration
2.2. The Expression of tNOX is Transcriptionally Regulated by POU3F2 in Human Stomach Cancer Cells
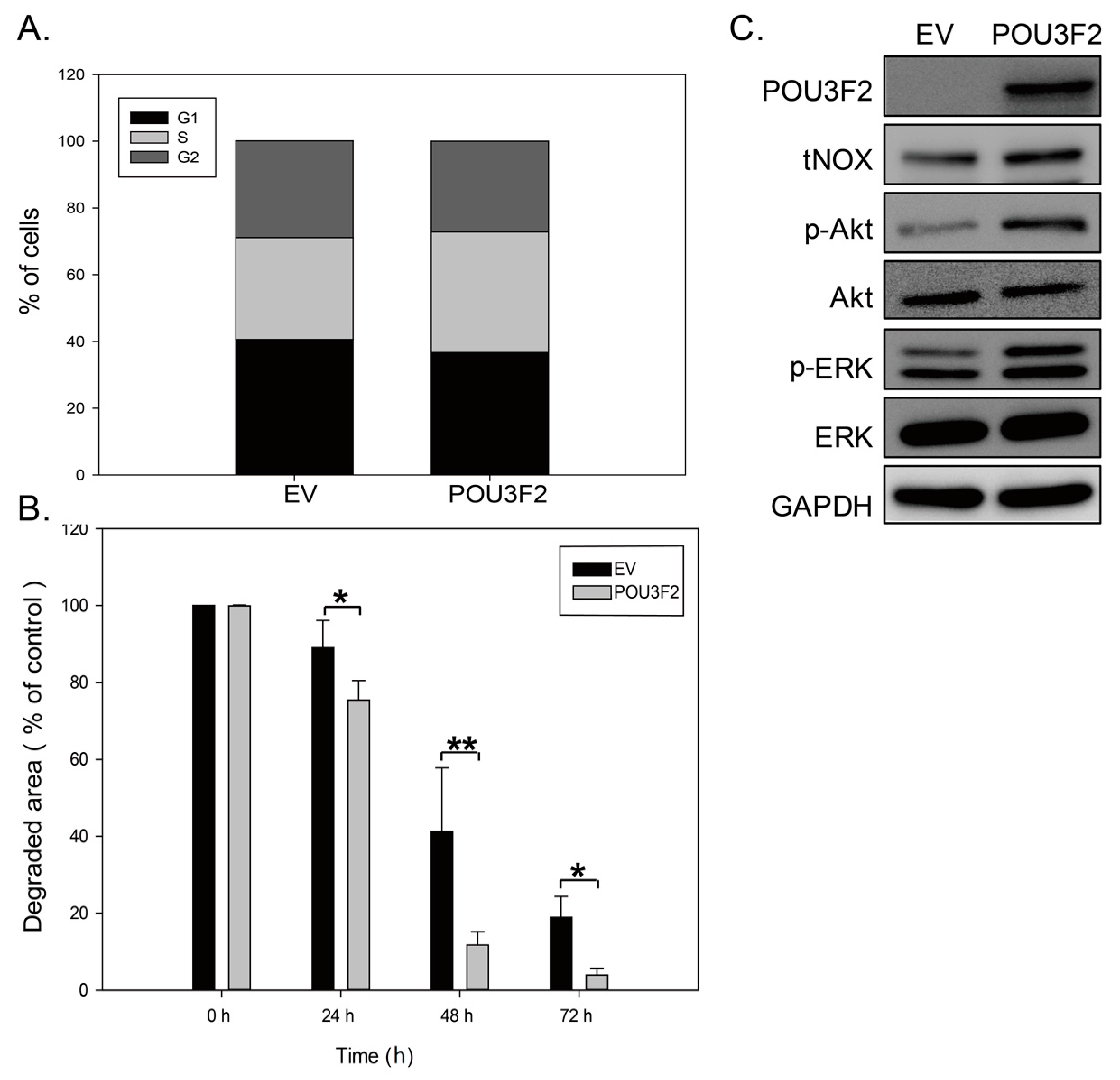
2.3. POU3F2 Overexpression Upregulates tNOX Expression and Increases Cell Proliferation
2.4. POU3F2-Mediated tNOX Upregulation Shortens the Cell Doubling Time
2.5. POU3F2-Mediated tNOX Upregulation Enhances Cell Migration and Invasion
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture and Transfection
4.3. Plasmid Constructs and Luciferase Assay
4.4. Continuous Cell Monitoring with the Cell Impedance Measurement System
4.5. Trypan Blue Exclusion Assay
4.6. Cell Cycle Analysis
4.7. Cell Division Assay
4.8. Boyden Chamber Assay
4.9. Western Blot Analysis
4.10. Reverse Transcriptase-Polymerase Chain Reaction
4.11. Statistics
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Morré, D.J.; Chueh, P.J.; Morré, D.M. Capsaicin inhibits preferentially the NADH oxidase and growth of transformed cells in culture. Proc. Natl. Acad. Sci. USA 1995, 92, 1831–1835. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Richardson, R.L.; Dashwood, R.H.; Baek, S.J. Capsaicin represses transcriptional activity of beta-catenin in human colorectal cancer cells. J. Nutr. Biochem. 2012, 23, 646–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.M.; Chuang, S.M.; Su, Y.C.; Li, Y.H.; Chueh, P.J. Down-regulation of tumor-associated NADH oxidase, tNOX (ENOX2), enhances capsaicin-induced inhibition of gastric cancer cell growth. Cell Biochem. Biophys. 2011, 61, 355–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huh, H.C.; Lee, S.Y.; Lee, S.K.; Park, N.H.; Han, I.S. Capsaicin induces apoptosis of cisplatin-resistant stomach cancer cells by causing degradation of cisplatin-inducible aurora-a protein. Nutr. Cancer 2011, 63, 1095–1103. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.C.; Wang, H.M.; Lin, Y.Y.; Chang, T.K.; Hsin, Y.H.; Chueh, P.J. Stress-induced down-regulation of tumor-associated NADH oxidase during apoptosis in transformed cells. FEBS Lett. 2008, 582, 3445–3450. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.M.; Chueh, P.J.; Chang, S.P.; Yang, C.L.; Shao, K.N. Effect of Ccapsaicin on tNOX (ENOX2) protein expression in stomach cancer cells. Biofactors 2009, 34, 209–217. [Google Scholar] [CrossRef]
- Liu, N.C.; Hsieh, P.F.; Hsieh, M.K.; Zeng, Z.M.; Cheng, H.L.; Liao, J.W.; Chueh, P.J. Capsaicin-mediated tNOX (ENOX2) up-regulation enhances cell proliferation and migration in vitro and in vivo. J. Agric. Food Chem. 2012, 60, 2758–2765. [Google Scholar] [CrossRef] [PubMed]
- Atanasoski, S.; Toldo, S.S.; Malipiero, U.; Schreiber, E.; Fries, R.; Fontana, A. Isolation of the human genomic brain-2/N-Oct 3 gene (POUF3) and assignment to chromosome 6q16. Genomics 1995, 26, 272–280. [Google Scholar] [CrossRef]
- Smit, D.J.; Smith, A.G.; Parsons, P.G.; Muscat, G.E.O.; Sturm, R.A. Domains of Brn-2 that mediate homodimerization and interaction with general and melanocytic transcription factors. Eur. J. Biochem. 2000, 267, 6413–6422. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.T.; Iwamoto, K.; Kurosaki, T.; Nasu, M.; Ueda, S. The neuronal POU transcription factor Brn-2 interacts with Jab1, a gene involved in the onset of neurodegenerative diseases. Neurosci. Lett. 2005, 382, 175–178. [Google Scholar] [CrossRef] [PubMed]
- Castro, D.S.; Skowronska-Krawczyk, D.; Armant, O.; Donaldson, I.J.; Parras, C.; Hunt, C.; Critchley, J.A.; Nguyen, L.; Gossler, A.; Gottgens, B.; et al. Proneural bHLH and Brn proteins coregulate a neurogenic program through cooperative binding to a conserved DNA motif. Dev. Cell 2006, 11, 831–844. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Liu, L.; Bian, W.; Chen, Y.; Xu, G.; Cheng, L.; Jing, N. Different transcription factors regulate nestin gene expression during p19 cell neural differentiation and central nervous system development. J. Biol. Chem. 2009, 284, 8160–8173. [Google Scholar] [CrossRef] [PubMed]
- McEvilly, R.J.; de Diaz, M.O.; Schonemann, M.D.; Hooshmand, F.; Rosenfeld, M.G. Transcriptional regulation of cortical neuron migration by pou domain factors. Science 2002, 295, 1528–1532. [Google Scholar] [CrossRef] [PubMed]
- Sugitani, M.; Sugai, T.; Onoda, N. Postsynaptic activity of metabotropic glutamate receptors in the piriform cortex. Neuroreport 2002, 13, 1473–1476. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, M.H.; Ayoub, A.E.; Rakic, P. POU-III transcription factors (Brn1, Brn2, and Oct6) influence neurogenesis, molecular identity, and migratory destination of upper-layer cells of the cerebral cortex. Cereb. Cortex 2013, 23, 2632–2643. [Google Scholar] [CrossRef] [PubMed]
- Nakai, S.; Kawano, H.; Yudate, T.; Nishi, M.; Kuno, J.; Nagata, A.; Jishage, K.; Hamada, H.; Fujii, H.; Kawamura, K.; et al. The POU domain transcription factor Brn-2 is required for the determination of specific neuronal lineages in the hypothalamus of the mouse. Genes Dev. 1995, 9, 3109–3121. [Google Scholar] [CrossRef] [PubMed]
- Schonemann, M.D.; Ryan, A.K.; McEvilly, R.J.; O’Connell, S.M.; Arias, C.A.; Kalla, K.A.; Li, P.; Sawchenko, P.E.; Rosenfeld, M.G. Development and survival of the endocrine hypothalamus and posterior pituitary gland requires the neuronal POU domain factor Brn-2. Genes Dev. 1995, 9, 3122–3135. [Google Scholar] [CrossRef] [PubMed]
- Wellbrock, C.; Rana, S.; Paterson, H.; Pickersgill, H.; Brummelkamp, T.; Marais, R. Oncogenic braf regulates melanoma proliferation through the lineage specific factor mitf. PLoS ONE 2008, 3, e2734. [Google Scholar] [CrossRef] [PubMed]
- Ishii, J.; Sato, H.; Yazawa, T.; Shishido-Hara, Y.; Hiramatsu, C.; Nakatani, Y.; Kamma, H. Class III/IV POU transcription factors expressed in small cell lung cancer cells are involved in proneural/neuroendocrine differentiation. Pathol. Int. 2014, 64, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Ishii, J.; Sato, H.; Sakaeda, M.; Shishido-Hara, Y.; Hiramatsu, C.; Kamma, H.; Shimoyamada, H.; Fujiwara, M.; Endo, T.; Aoki, I.; et al. POU domain transcription factor Brn2 is crucial for expression of ASCL1, ND1 and neuroendocrine marker molecules and cell growth in small cell lung cancer. Pathol. Int. 2013, 63, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Li, H.; Zhang, W.; Matkowskyj, K.A.; Liao, J.; Srivastava, S.K.; Yang, G.Y. Inhibition of chronic pancreatitis and pancreatic intraepithelial neoplasia (PanIN) by capsaicin in Lsl-KrasG12D/Pdx1-Cre mice. Carcinogenesis 2011, 32, 1689–1696. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.C.; Yang, J.J.; Shao, K.N.; Chueh, P.J. RNA interference targeting tNOX attenuates cell migration via a mechanism that involves membrane association of Rac. Biochem. Biophys. Res. Commun. 2008, 365, 672–677. [Google Scholar] [CrossRef] [PubMed]
- Ke, N.; Wang, X.; Xu, X.; Abassi, Y.A. The xcelligence system for real-time and label-free monitoring of cell viability. Methods Mol. Biol. 2011, 740, 33–43. [Google Scholar] [PubMed]
- Ungefroren, H.; Sebens, S.; Groth, S.; Gieseler, F.; Fandrich, F. Differential roles of Src in transforming growth factor-ss regulation of growth arrest, epithelial-to-mesenchymal transition and cell migration in pancreatic ductal adenocarcinoma cells. Int. J. Oncol. 2011, 38, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [PubMed]
- Shih, J.Y.; Yang, P.C. The emt regulator slug and lung carcinogenesis. Carcinogenesis 2011, 32, 1299–1304. [Google Scholar] [CrossRef] [PubMed]
- Goodall, J.; Wellbrock, C.; Dexter, T.J.; Roberts, K.; Marais, R.; Goding, C.R. The Brn-2 transcription factor links activated BRAF to melanoma proliferation. Mol. Cell. Biol. 2004, 24, 2923–2931. [Google Scholar] [CrossRef] [PubMed]
- Goodall, J.; Martinozzi, S.; Dexter, T.J.; Champeval, D.; Carreira, S.; Larue, L.; Goding, C.R. Brn-2 expression controls melanoma proliferation and is directly regulated by beta-catenin. Mol. Cell. Biol. 2004, 24, 2915–2922. [Google Scholar] [CrossRef] [PubMed]
- Thomson, J.A.; Murphy, K.; Baker, E.; Sutherland, G.R.; Parsons, P.G.; Sturm, R.A.; Thomson, F. The Brn-2 gene regulates the melanocytic phenotype and tumorigenic potential of human melanoma cells. Oncogene 1995, 11, 691–700. [Google Scholar] [PubMed]
- Suva, M.L.; Rheinbay, E.; Gillespie, S.M.; Patel, A.P.; Wakimoto, H.; Rabkin, S.D.; Riggi, N.; Chi, A.S.; Cahill, D.P.; Nahed, B.V.; et al. Reconstructing and reprogramming the tumor-propagating potential of glioblastoma stem-like cells. Cell 2014, 157, 580–594. [Google Scholar] [CrossRef] [PubMed]
- Cook, A.L.; Sturm, R.A. POU domain transcription factors: Brn2 as a regulator of melanocytic growth and tumourigenesis. Pigment Cell Melanoma Res. 2008, 21, 611–626. [Google Scholar] [CrossRef] [PubMed]
- Ellmann, L.; Joshi, M.B.; Resink, T.J.; Bosserhoff, A.K.; Kuphal, S. BRN2 is a transcriptional repressor of CDH13(T-cadherin) in melanoma cells. Lab. Investig. 2012, 92, 1788–1800. [Google Scholar] [CrossRef] [PubMed]
- Chueh, P.J. Cell membrane redox systems and transformation. Antioxid. Redox Signal. 2000, 2, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Morré, D.J.; Caldwell, S.; Mayorga, A.; Wu, L.Y.; Morré, D.M. NADH oxidase activity from sera altered by capsaicin is widely distributed among cancer patients. Arch. Biochem. Biophys. 1997, 342, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Morré, D.J.; Reust, T. A circulating form of NADH oxidase activity responsive to the antitumor sulfonylurea N-4-(methylphenylsulfonyl)-N′-(4-chlorophenyl)urea (LY181984) specific to sera from cancer patients. J. Bioenerg. Biomembr. 1997, 29, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Chueh, P.J.; Morre, D.J.; Wilkinson, F.E.; Gibson, J.; Morré, D.M. A 33.5-kDa heat- and protease-resistant NADH oxidase inhibited by capsaicin from sera of cancer patients. Arch. Biochem. Biophys. 1997, 342, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Chueh, P.J.; Wu, L.Y.; Morré, D.M.; Morré, D.J. tNOX is both necessary and sufficient as a cellular target for the anticancer actions of capsaicin and the green tea catechin (−)-epigallocatechin-3-gallate. Biofactors 2004, 20, 235–249. [Google Scholar] [PubMed]
- Su, Y.C.; Lin, Y.H.; Zeng, Z.M.; Shao, K.N.; Chueh, P.J. Chemotherapeutic agents enhance cell migration and epithelial-to-mesenchymal transition through transient up-regulation of tNOX (ENOX2) protein. Biochim. Biophys. Acta 2012, 1820, 1744–1752. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.F.; Huang, S.; Liu, S.C.; Chueh, P.J. Effect of polyclonal antisera to recombinant tNOX protein on the growth of transformed cells. Biofactors 2006, 28, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Chueh, P.J.; Lin, Y.W.; Shih, T.S.; Chuang, S.M. Disturbed mitotic progression and genome segregation are involved in cell transformation mediated by nano-TiO2 long-term exposure. Toxicol. Appl. Pharmacol. 2009, 241, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Chuang, S.M.; Lee, Y.H.; Liang, R.Y.; Roam, G.D.; Zeng, Z.M.; Tu, H.F.; Wang, S.K.; Chueh, P.J. Extensive evaluations of the cytotoxic effects of gold nanoparticles. Biochim. Biophys. Acta 2013, 1830, 4960–4973. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the tNOX promoter reporter constructs are available from the authors.








© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, H.Y.; Lee, Y.H.; Chen, H.Y.; Yeh, C.A.; Chueh, P.J.; Lin, Y.-M.J. Capsaicin Inhibited Aggressive Phenotypes through Downregulation of Tumor-Associated NADH Oxidase (tNOX) by POU Domain Transcription Factor POU3F2. Molecules 2016, 21, 733. https://doi.org/10.3390/molecules21060733
Chen HY, Lee YH, Chen HY, Yeh CA, Chueh PJ, Lin Y-MJ. Capsaicin Inhibited Aggressive Phenotypes through Downregulation of Tumor-Associated NADH Oxidase (tNOX) by POU Domain Transcription Factor POU3F2. Molecules. 2016; 21(6):733. https://doi.org/10.3390/molecules21060733
Chicago/Turabian StyleChen, Hung Yen, Yi Hui Lee, Huei Yu Chen, Chia An Yeh, Pin Ju Chueh, and Yi-Mei J. Lin. 2016. "Capsaicin Inhibited Aggressive Phenotypes through Downregulation of Tumor-Associated NADH Oxidase (tNOX) by POU Domain Transcription Factor POU3F2" Molecules 21, no. 6: 733. https://doi.org/10.3390/molecules21060733
APA StyleChen, H. Y., Lee, Y. H., Chen, H. Y., Yeh, C. A., Chueh, P. J., & Lin, Y. -M. J. (2016). Capsaicin Inhibited Aggressive Phenotypes through Downregulation of Tumor-Associated NADH Oxidase (tNOX) by POU Domain Transcription Factor POU3F2. Molecules, 21(6), 733. https://doi.org/10.3390/molecules21060733