
Efficacy of Acetylshikonin in Preventing Obesity and Hepatic Steatosis in db/db Mice

Abstract
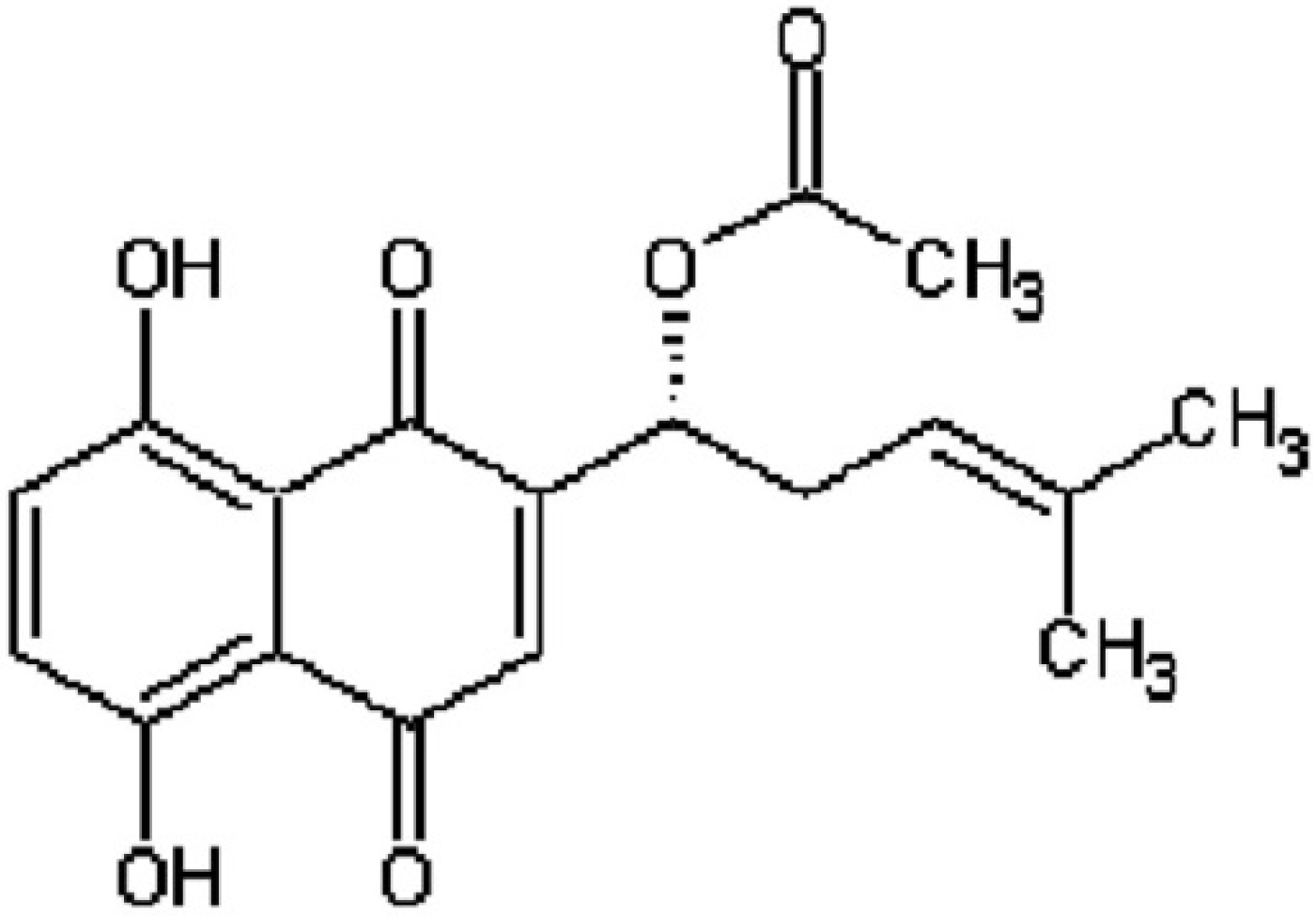
:
1. Introduction
2. Results
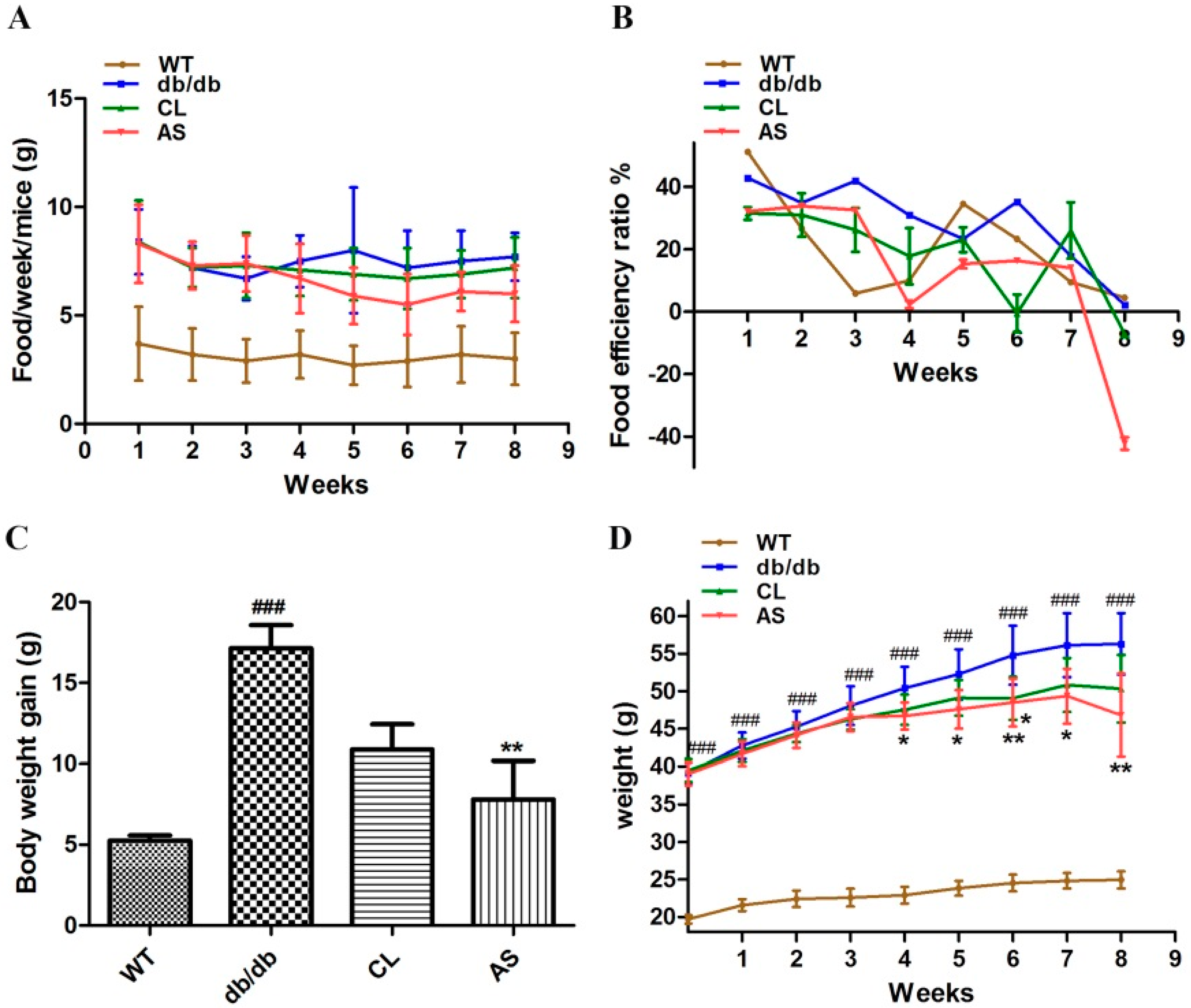
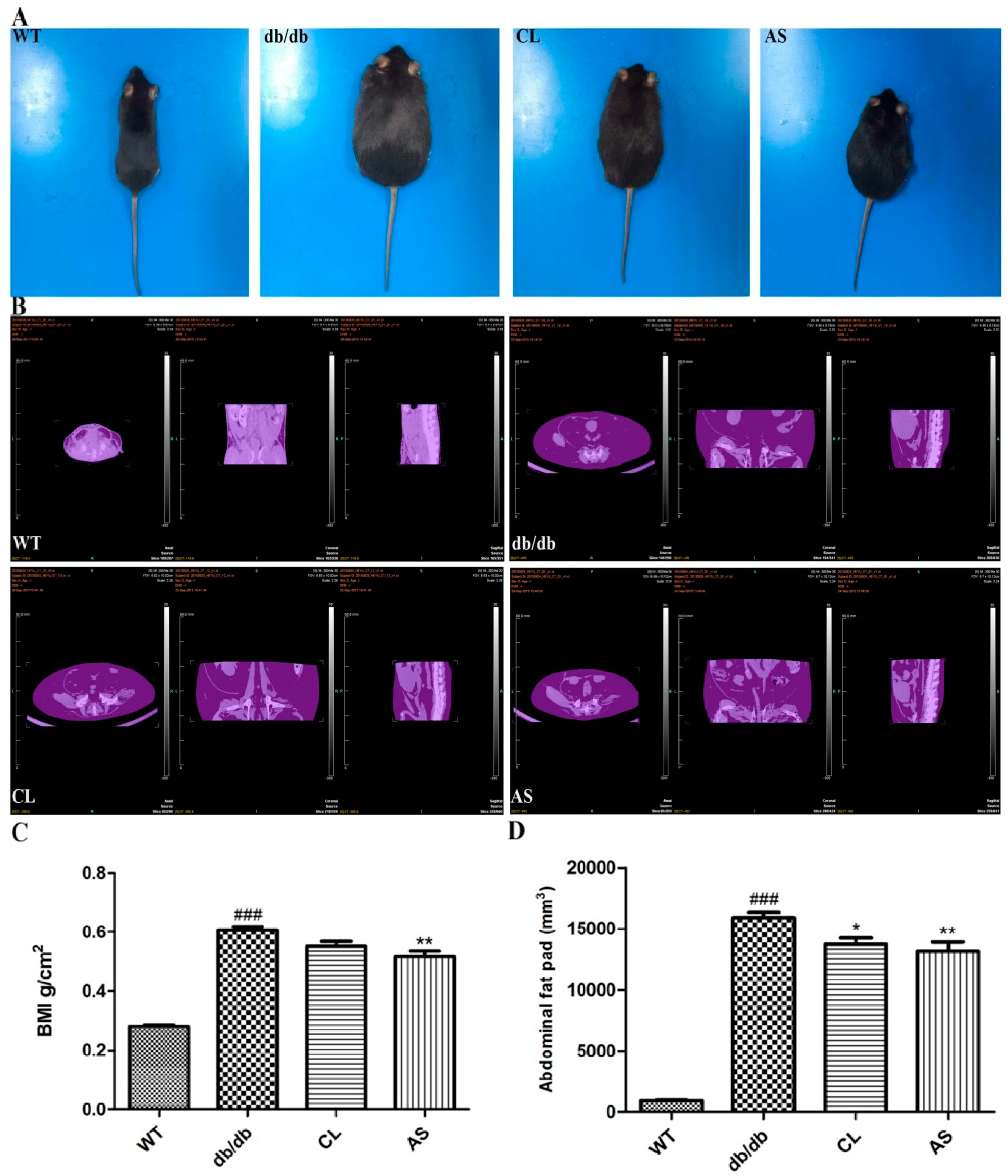
2.1. AS Suppressed Obesity in db/db Mice
2.2. Effect of AS on Abdominal Adiposity in db/db Mice
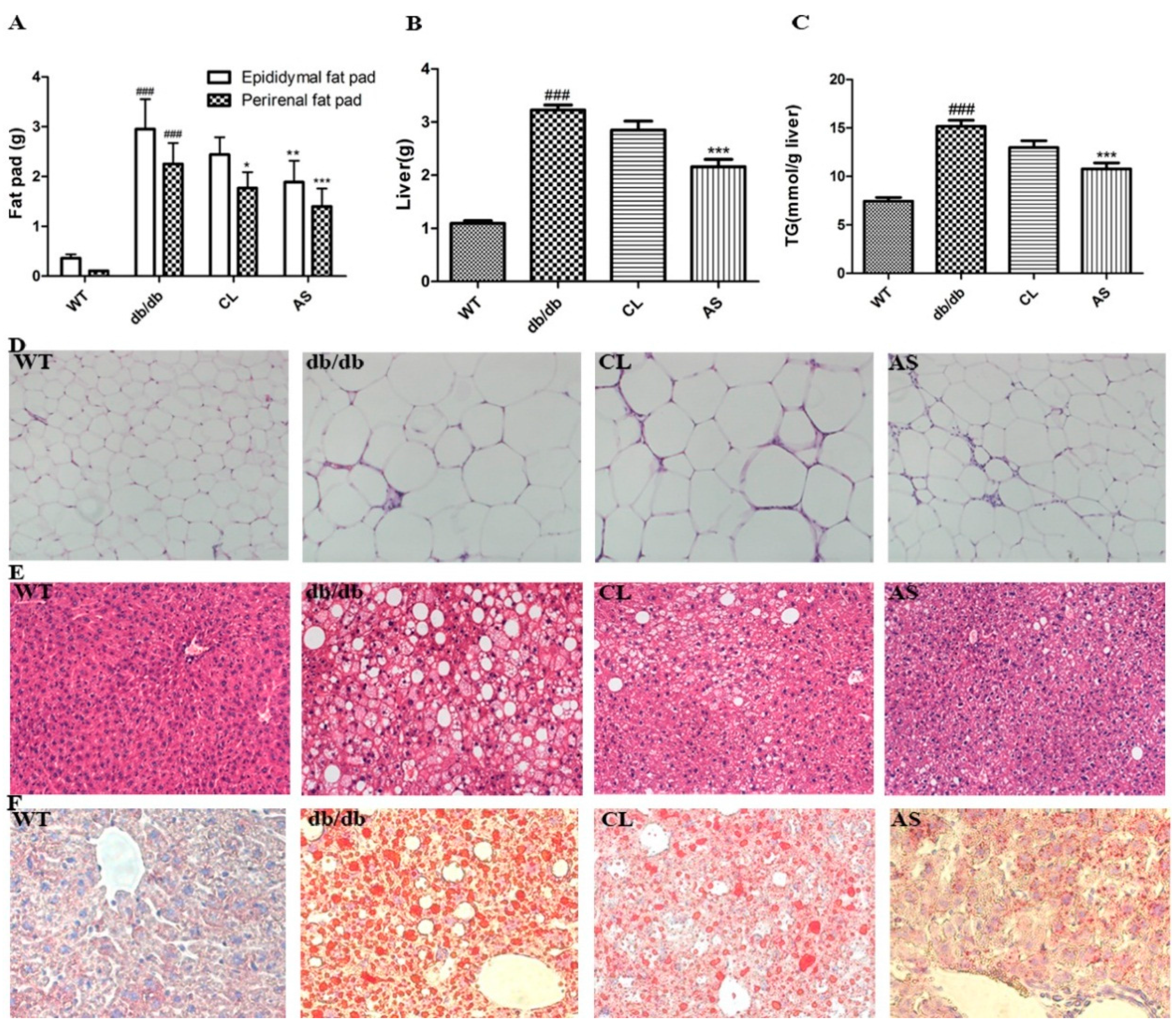
2.3. AS Reduced Adipose Tissue, Liver Weight and Hepatic Lipid Accumulation in db/db Mice
2.4. AS Decreased the Blood Lipids and Hepatic Enzyme Levels in db/db Mice
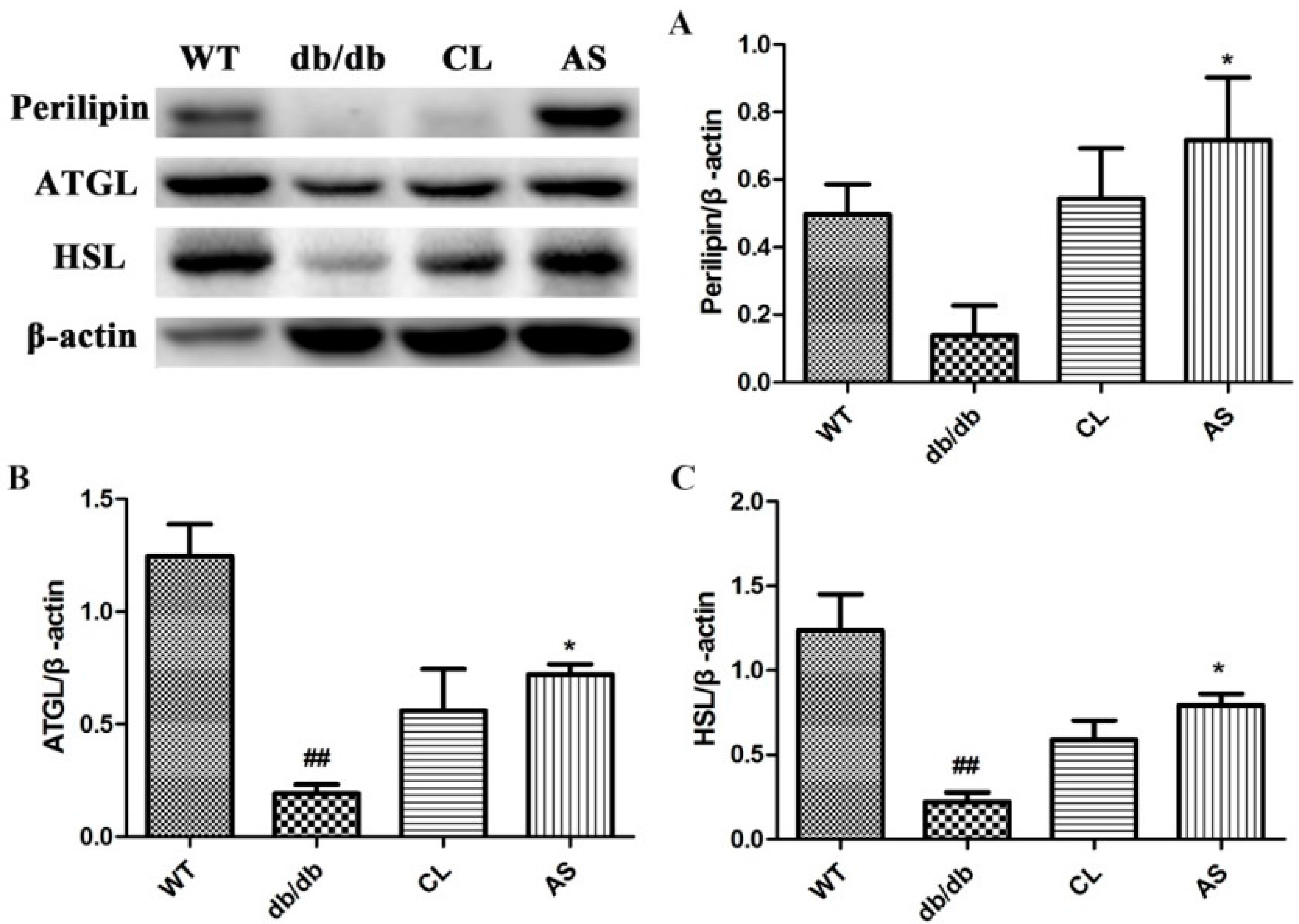
2.5. AS Increased the Expressions of Lipid-Metabolizing Enzymes in Adipose Tissue
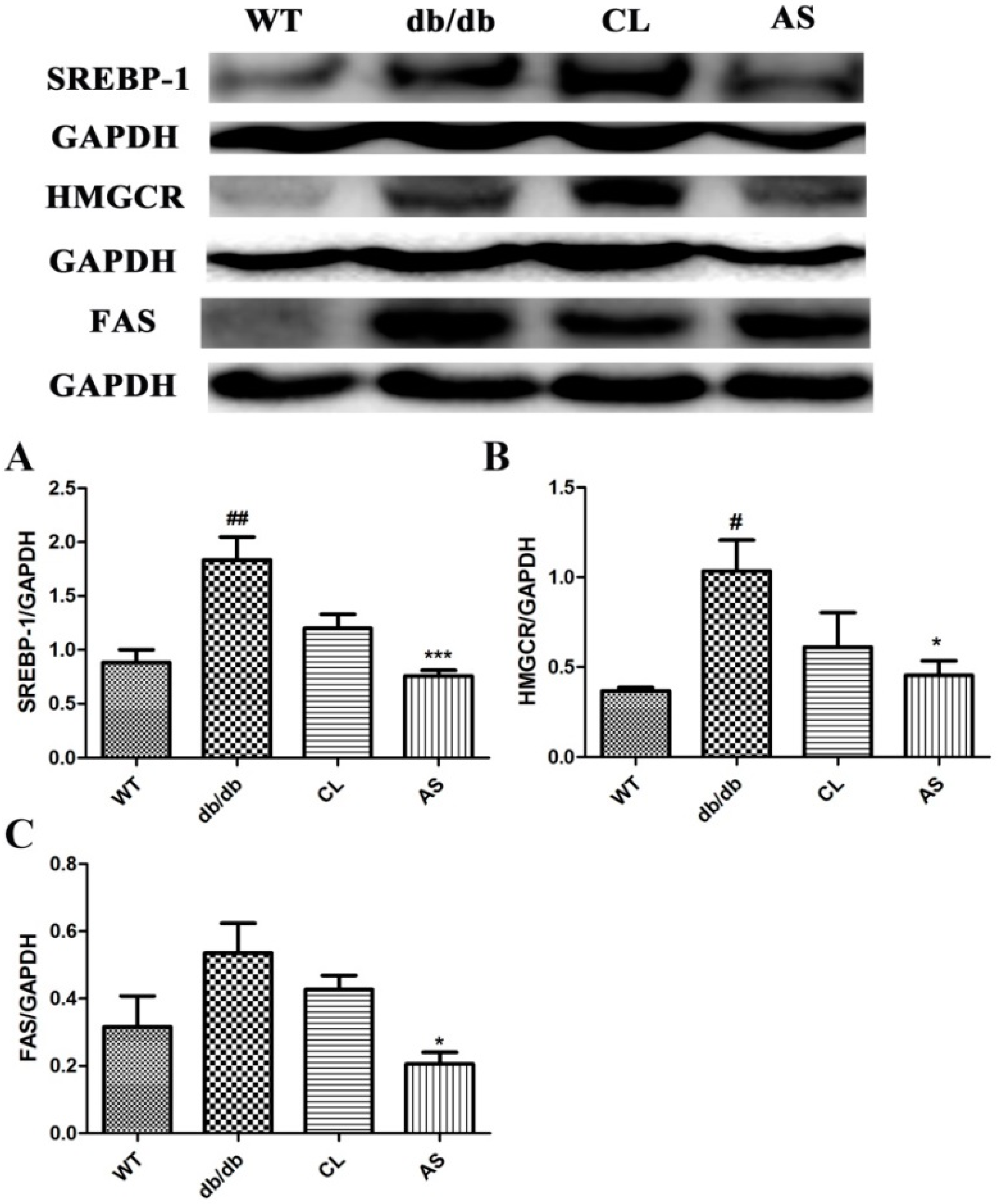
2.6. AS Inhibited the Lipid Synthesis-Related Protein Expressions (SREBP-1, FAS and HMGCR) in the Liver
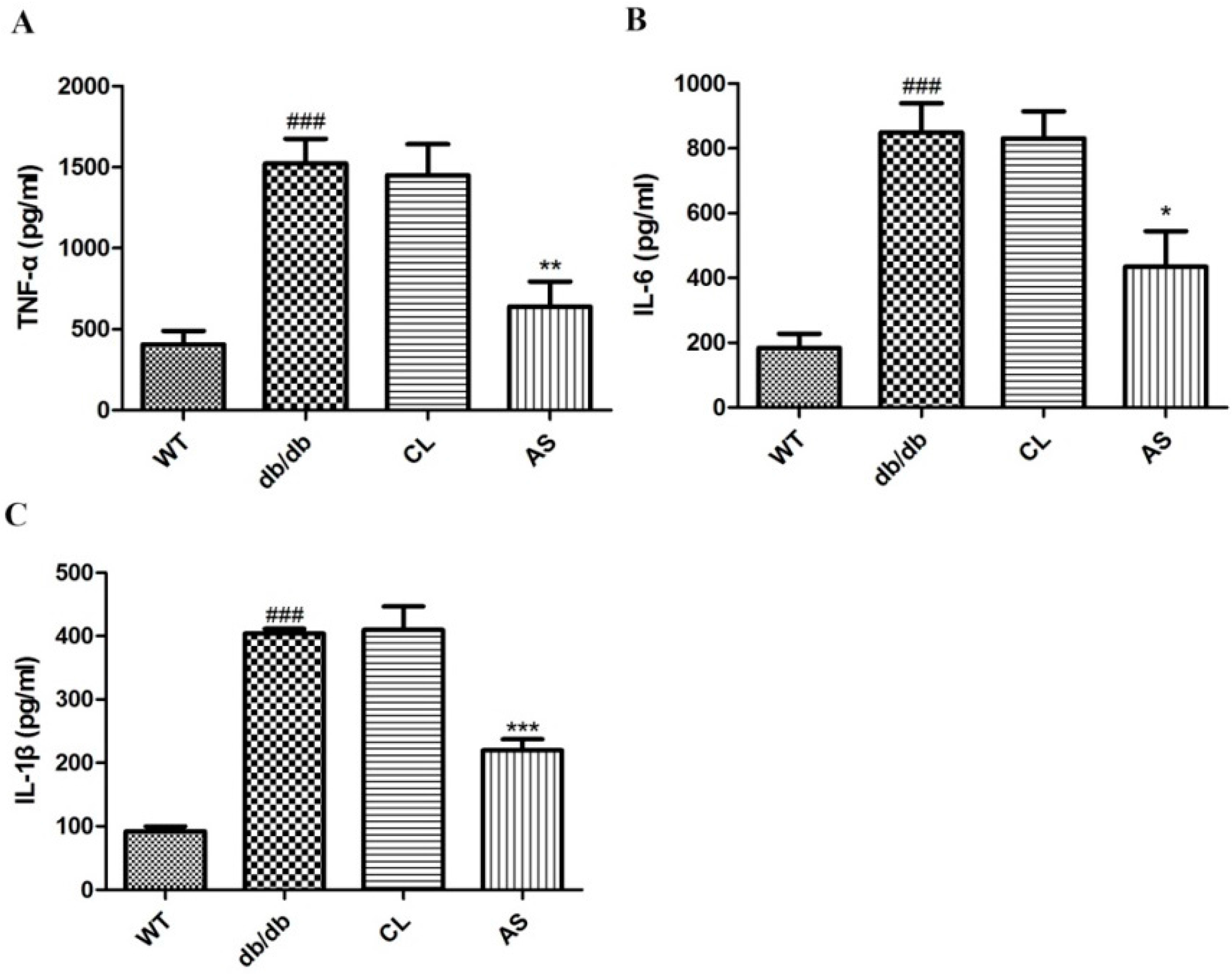
2.7. Effect of AS on Serum Pro-Inflammatory Cytokines in db/db Mice
3. Experimental Section
3.1. Reagents and Antibodies
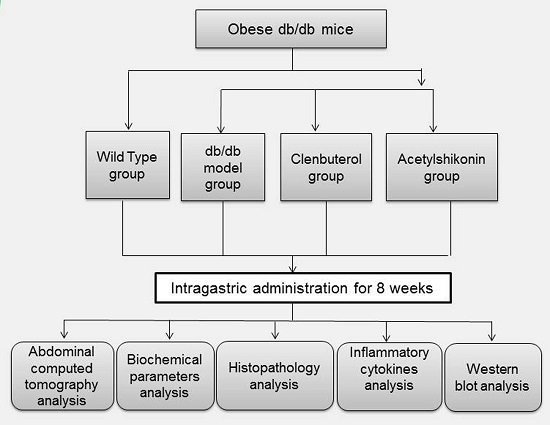
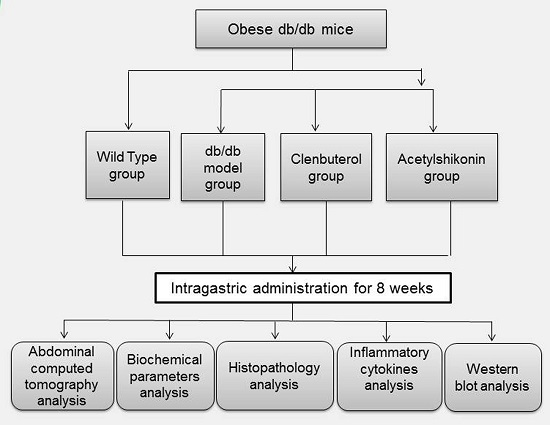
3.2. Mice and Experimental Design
3.3. Abdominal Computed Tomography Analysis
3.4. Liver Lipid Content
3.5. Histological Analysis
3.6. Biochemical Parameters Analysis
3.7. Western Blot Analysis
3.8. Statistical Analysis
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AS | acetylshikonin |
| WT | Wild Type |
| CL | clenbuterol |
| NAFLD | nonalcoholic fatty liver disease |
| NASH | nonalcoholic steatohepatitis |
| TG | triglyceride |
| FFA | free fatty acid |
| CHE | cholinesterase |
| AST | alanine aminotransferase |
| ALT | alanine aminotransferase |
| SREBP-1 | sterol regulatory element-binding protein-1 |
| FAS | fatty acid synthetase |
| HMGCR | 3-hydroxy-3-methylglutaryl-coenzyme A reductase |
| HSL | hormone-sensitive lipase |
| ATGL | adipose TG lipase |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| ELISA | enzyme-linked immunosorbent assay |
| TNF-α | tumour necrosis factor-a |
| IL-6 | interleukin-6 |
| IL-1β | interleukin-1β BMI body mass index |
| T2DM | type 2 diabetes mellitus |
| ANOVA | one-way analysis of variance |
References
- Kyrou, I.; Randeva, H.S.; Weickert, M.O. Clinical Problems Caused by Obesity. In Endotext (Internet); MDText.com, Inc.: Dartmouth, MA, USA, 2000. [Google Scholar]
- Williams, K.H.; Shackel, N.A.; Gorrell, M.D.; McLennan, S.V.; Twigg, S.M. Diabetes and nonalcoholic Fatty liver disease: A pathogenic duo. Endocr. Rev. 2013, 34, 84–129. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.W.; Adams, L.A. Nonalcoholic fatty liver disease and diabetes mellitus: Pathogenesis and treatment. Nat. Rev. Endocrinol. 2011, 7, 456–465. [Google Scholar] [CrossRef] [PubMed]
- Kaswala, D.H.; Lai, M.; Afdhal, N.H. Fibrosis Assessment in Nonalcoholic Fatty Liver Disease (NAFLD) in 2016. Dig. Dis. Sci. 2016, 61, 1356–1364. [Google Scholar] [CrossRef] [PubMed]
- Kratky, D.; Obrowsky, S.; Kolb, D.; Radovic, B. Pleiotropic regulation of mitochondrial function by adipose triglyceride lipase-mediated lipolysis. Biochimie 2014, 96, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Morak, M.; Schmidinger, H.; Riesenhuber, G.; Rechberger, G.N.; Kollroser, M.; Haemmerle, G.; Zechner, R.; Kronenberg, F.; Hermetter, A. Adipose triglyceride lipase (ATGL) and hormone-sensitive lipase (HSL) deficiencies affect expression of lipolytic activities in mouse adipose tissues. Mol. Cell. Proteom. 2012, 11, 1777–1789. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, J.Y.; Han, M.; Wang, W.L.; Li, Y.Z. Prevention and treatment effect of total flavonoids in Stellera chamaejasme L. on nonalcoholic fatty liver in rats. Lipids Health Dis. 2015, 14. [Google Scholar] [CrossRef] [PubMed]
- Moon, Y.A.; Liang, G.; Xie, X.; Frank-Kamenetsky, M.; Fitzgerald, K.; Koteliansky, V.; Brown, M.S.; Goldstein, J.L.; Horton, J.D. The Scap/SREBP pathway is essential for developing diabetic fatty liver and carbohydrate-induced hypertriglyceridemia in animals. Cell Metab. 2012, 15, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Devisscher, L.; Verhelst, X.; Colle, I.; van Vlierberghe, H.; Geerts, A. The role of macrophages in obesity-driven chronic liver disease. J. Leukoc. Biol. 2016, 99, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Lanthier, N.; Molendi-Coste, O.; Cani, P.D.; van Rooijen, N.; Horsmans, Y.; Leclercq, I.A. Kupffer cell depletion prevents but has no therapeutic effect on metabolic and inflammatory changes induced by a high-fat diet. FASEB J. 2011, 25, 4301–4311. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Moschen, A.R.; Szabo, G. Interleukin-1 and inflammasomes in ALD/AAH and NAFLD/NASH. Hepatology 2016. [Google Scholar] [CrossRef] [PubMed]
- Moschen, A.R.; Kaser, S.; Tilg, H. Non-alcoholic steatohepatitis: A microbiota-driven disease. Trends Endocrinol. Metab. 2013, 24, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Tsochatzis, E.A.; Papatheodoridis, G.V.; Archimandritis, A.J. Adipokines in nonalcoholic steatohepatitis: From pathogenesis to implications in diagnosis and therapy. Mediat. Inflamm. 2009, 2009. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yang, L.; Oppenheim, J.J.; Howard, M.Z. Cellular pharmacology studies of shikonin derivatives. Phytother. Res. 2002, 16, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Zhu, B.H. Arnebin-1 promotes the angiogenesis of human umbilical vein endothelial cells and accelerates the wound healing process in diabetic rats. J. Ethnopharmacol. 2014, 154, 653–662. [Google Scholar] [CrossRef] [PubMed]
- Andujar, I.; Rios, J.L.; Giner, R.M.; Recio, M.C. Pharmacological properties of shikonin—A review of literature since 2002. Planta Med. 2013, 79, 1685–1697. [Google Scholar] [CrossRef] [PubMed]
- Su, M.; Huang, W.; Zhu, B. Acetylshikonin from Zicao Prevents Obesity in Rats on a High-Fat Diet by Inhibiting Lipid Accumulation and Inducing Lipolysis. PLoS ONE 2016, 11, e146884. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.A.; Mirza, M.H.; Barker, S.A.; Morgan, T.W.; Bauer, R.W.; McConnico, R.S. Clenbuterol toxicosis in three Quarter Horse racehorses after administration of a compounded product. J. Am. Vet. Med. Assoc. 2011, 239, 842–849. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane, S.G. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Chalasani, N.; Younossi, Z.; Lavine, J.E.; Diehl, A.M.; Brunt, E.M.; Cusi, K.; Charlton, M.; Sanyal, A.J. The diagnosis and management of non-alcoholic fatty liver disease: Practice Guideline by the American Association for the Study of Liver Diseases, American College of Gastroenterology, and the American Gastroenterological Association. Hepatology 2012, 55, 2005–2023. [Google Scholar] [CrossRef] [PubMed]
- Leung, W.Y.; Thomas, G.N.; Chan, J.C.; Tomlinson, B. Weight management and current options in pharmacotherapy: Orlistat and sibutramine. Clin. Ther. 2003, 25, 58–80. [Google Scholar] [CrossRef]
- Padwal, R.S.; Majumdar, S.R. Drug treatments for obesity: Orlistat, sibutramine, and rimonabant. Lancet 2007, 369, 71–77. [Google Scholar] [CrossRef]
- Kim, H.K.; Della-Fera, M.A.; Hausman, D.B.; Baile, C.A. Effect of clenbuterol on apoptosis, adipogenesis, and lipolysis in adipocytes. J. Physiol. Biochem. 2010, 66, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Yen, M.; Ewald, M.B. Toxicity of weight loss agents. J. Med. Toxicol. 2012, 8, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, Y.; Ohno, A.; Saito, Y.; Satake, M. Accelerative effect of shikonin, alkannin and acetylshikonin on the proliferation of granulation tissue in rats. Biol. Pharm. Bull. 1994, 17, 1075–1077. [Google Scholar] [CrossRef] [PubMed]
- Kuo, H.M.; Hsia, T.C.; Chuang, Y.C.; Lu, H.F.; Lin, S.Y.; Chung, J.G. Shikonin inhibits the growth and N-acetylation of 2-aminofluorene in Helicobacter pylori from ulcer patients. Anticancer Res. 2004, 24, 1587–1592. [Google Scholar] [PubMed]
- Ruhl, C.E.; Everhart, J.E. Determinants of the association of overweight with elevated serum alanine aminotransferase activity in the United States. Gastroenterology 2003, 124, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Wronska, A.; Kmiec, Z. Structural and biochemical characteristics of various white adipose tissue depots. Acta Physiol. (Oxf.) 2012, 205, 194–208. [Google Scholar] [CrossRef] [PubMed]
- Fabbrini, E.; Sullivan, S.; Klein, S. Obesity and nonalcoholic fatty liver disease: Biochemical, metabolic, and clinical implications. Hepatology 2010, 51, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Fabbrini, E.; Magkos, F. Hepatic Steatosis as a Marker of Metabolic Dysfunction. Nutrients 2015, 7, 4995–5019. [Google Scholar] [CrossRef] [PubMed]
- Carmen, G.Y.; Victor, S.M. Signalling mechanisms regulating lipolysis. Cell Signal. 2006, 18, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Holm, C. Molecular mechanisms regulating hormone-sensitive lipase and lipolysis. Biochem. Soc. Trans. 2003, 31, 1120–1124. [Google Scholar] [CrossRef] [PubMed]
- Semiserin, V.A.; Karakozov, A.G.; Mal’Kuta, M.A.; Zolotareva, L.A.; Levchenko, O.B.; Kalyagin, I.E.; Eremin, M.N. Evaluation of the efficiency of hepatoprotective monotherapy using succinic acid and methionine for nonalcoholic fatty liver disease at the stage of steatohepatitis. Ter Arkh 2016, 88, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Ameer, F.; Scandiuzzi, L.; Hasnain, S.; Kalbacher, H.; Zaidi, N. De novo lipogenesis in health and disease. Metabolism 2014, 63, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Day, C.P.; James, O.F. Steatohepatitis: A tale of two “hits”? Gastroenterology 1998, 114, 842–845. [Google Scholar] [CrossRef]
- Inzaugarat, M.E.; Ferreyra, S.N.; Billordo, L.A.; Abecasis, R.; Gadano, A.C.; Chernavsky, A.C. Altered phenotype and functionality of circulating immune cells characterize adult patients with nonalcoholic steatohepatitis. J. Clin. Immunol. 2011, 31, 1120–1130. [Google Scholar] [CrossRef] [PubMed]
- Braunersreuther, V.; Viviani, G.L.; Mach, F.; Montecucco, F. Role of cytokines and chemokines in non-alcoholic fatty liver disease. World J. Gastroenterol. 2012, 18, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Hotamisligil, G.S. Nonalcoholic fatty liver disease: Cytokine-adipokine interplay and regulation of insulin resistance. Gastroenterology 2006, 131, 934–945. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are available from the authors.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | WT | db/db | CL | AS |
|---|---|---|---|---|
| TG (mM) | 0.65 ± 0.13 | 1.92 ± 0.63 ## | 1.82 ± 0.75 | 0.83 ± 0.15 ** |
| FFA (mM) | 0.58 ± 0.07 | 1.26 ± 0.19 ## | 0.76 ± 0.20 ** | 0.68 ± 0.11 ** |
| Glucose(mM) | 5.72 ± 1.01 | 27.80 ± 4.07 ## | 25.75 ± 5.96 | 18.32 ± 2.63 ** |
| CHE(mM) | 3.40 ± 0.45 | 6.89 ± 0.57 ## | 6.63 ± 0.73 | 3.76 ± 0.37 ** |
| AST(IU/L) | 73.63 ± 8.33 | 116.42 ± 22.59 ## | 112.43 ± 23.18 | 84.78 ± 12.23 * |
| ALT(IU/L) | 20.82 ± 3.15 | 94.48 ± 24.90 ## | 99.02 ± 39.22 | 51.170 ± 14.74 * |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, M.-L.; He, Y.; Li, Q.-S.; Zhu, B.-H. Efficacy of Acetylshikonin in Preventing Obesity and Hepatic Steatosis in db/db Mice. Molecules 2016, 21, 976. https://doi.org/10.3390/molecules21080976
Su M-L, He Y, Li Q-S, Zhu B-H. Efficacy of Acetylshikonin in Preventing Obesity and Hepatic Steatosis in db/db Mice. Molecules. 2016; 21(8):976. https://doi.org/10.3390/molecules21080976
Chicago/Turabian StyleSu, Mei-Ling, Yu He, Qi-Sen Li, and Bang-Hao Zhu. 2016. "Efficacy of Acetylshikonin in Preventing Obesity and Hepatic Steatosis in db/db Mice" Molecules 21, no. 8: 976. https://doi.org/10.3390/molecules21080976
APA StyleSu, M. -L., He, Y., Li, Q. -S., & Zhu, B. -H. (2016). Efficacy of Acetylshikonin in Preventing Obesity and Hepatic Steatosis in db/db Mice. Molecules, 21(8), 976. https://doi.org/10.3390/molecules21080976