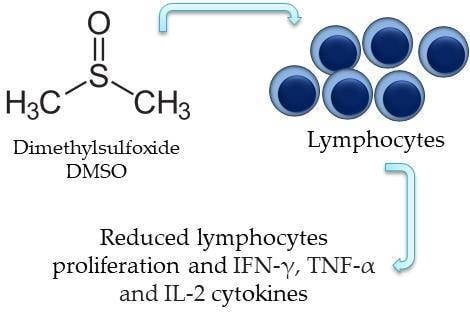
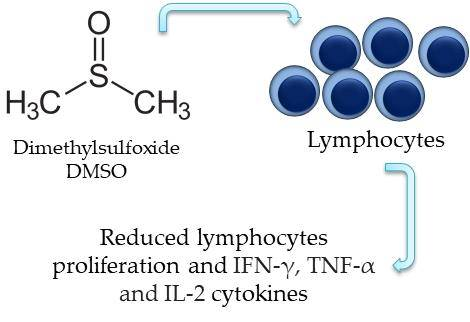
Dimethyl Sulfoxide (DMSO) Decreases Cell Proliferation and TNF-α, IFN-γ, and IL-2 Cytokines Production in Cultures of Peripheral Blood Lymphocytes

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
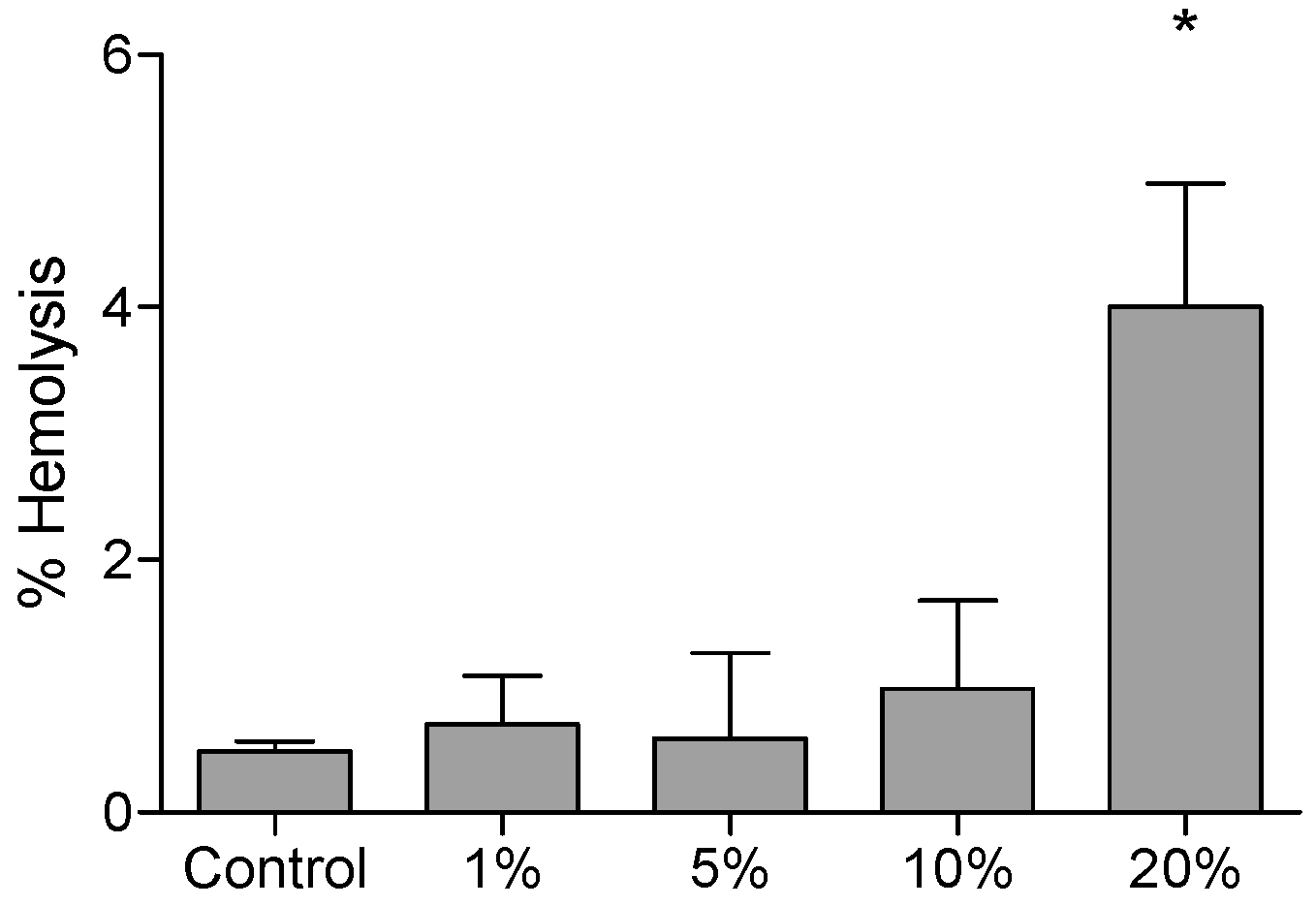
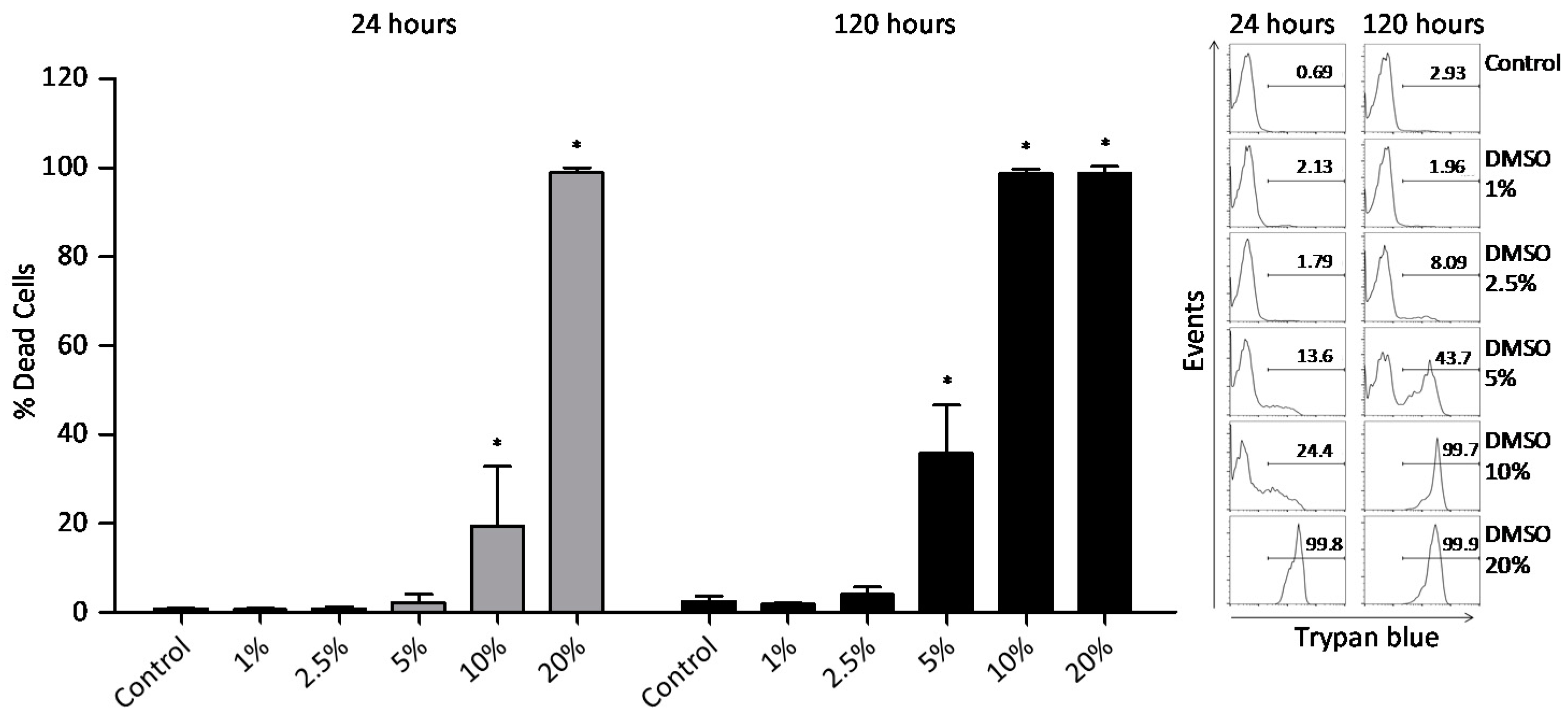
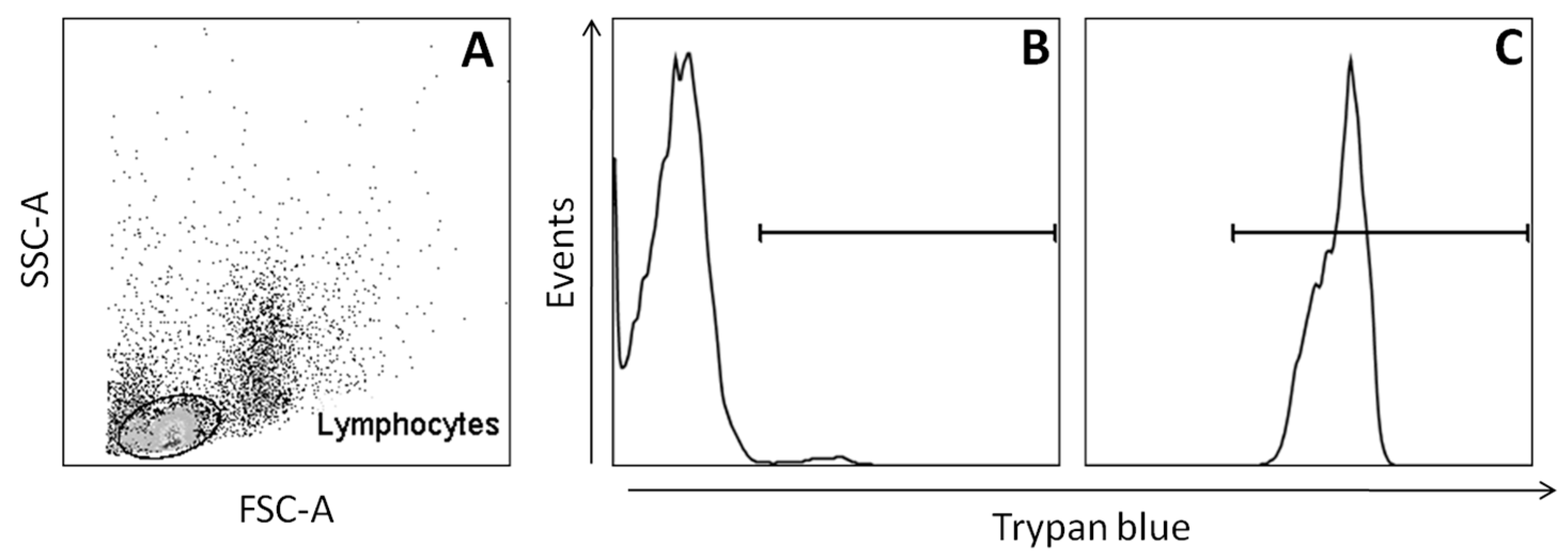
2.1. Cytotoxicity Tests (Assays)
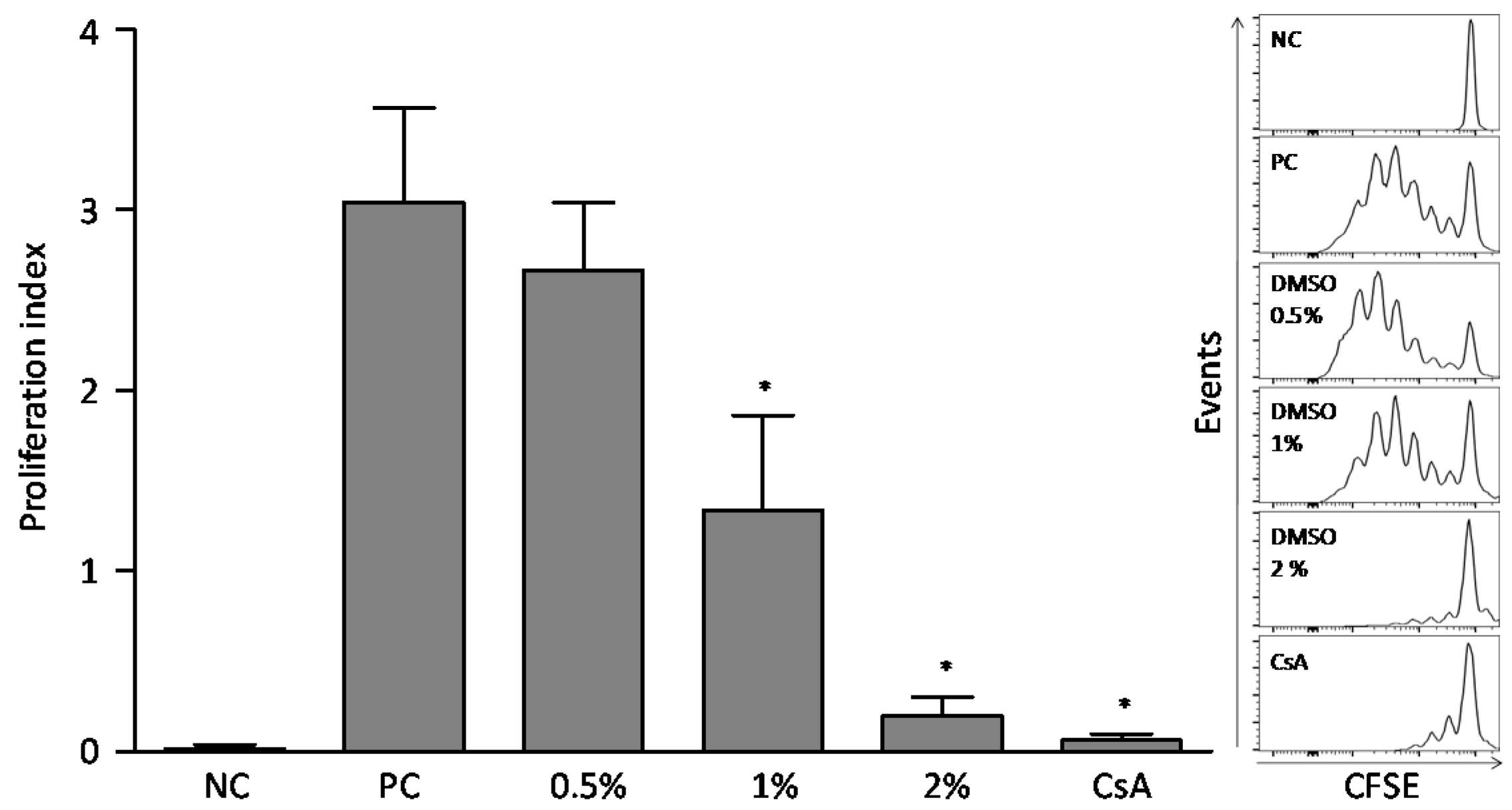
2.2. Dimethylsulfoxide Effect on Lymphocytes Proliferation
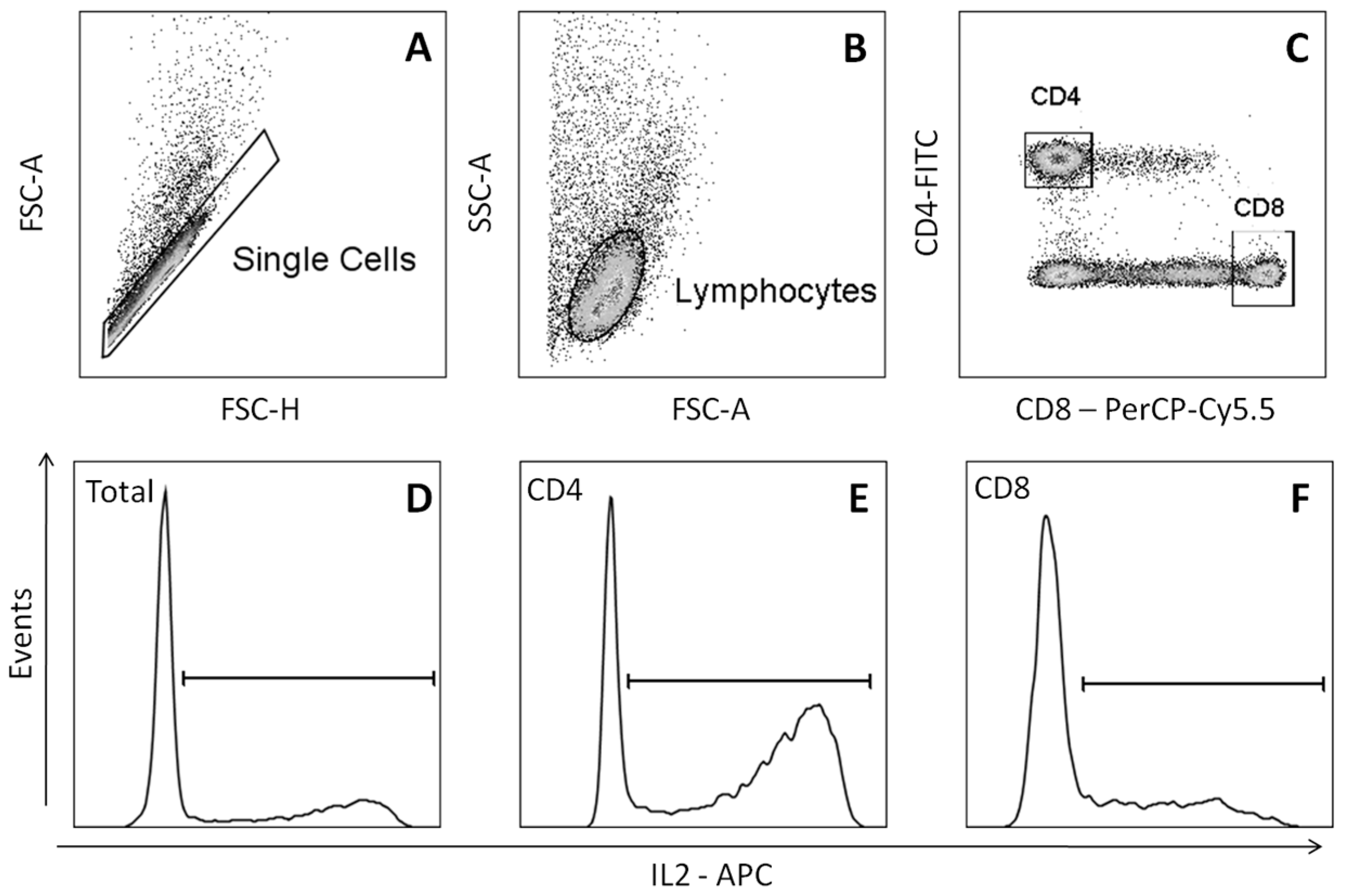
2.3. Effect of Dimethyl sulfoxide on the Production of Cytokines by Lymphocytes
3. Discussion
4. Materials and Methods
4.1. Study Subjects and Biological Samples
4.2. Hemolysis Tests
4.3. Peripheral Blood Mononuclear Cells (PBMC) Isolation
4.4. Peripheral Blood Mononuclear Cells (PBMC) Viability Analysis
4.5. Proliferation Analysis with 5-(and-6)-Carboxy-fluorescein succinimidyl ester (CFSE) Label Decay Technique
4.6. Analysis of the Cytokines Production by Lymphocytes
4.7. Statistical Analyses
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Brayton, C.F. Dimethyl sulfoxide (DMSO): A review. Cornell Vet. 1986, 76, 61–90. [Google Scholar] [PubMed]
- Timm, M.; Saaby, L.; Moesby, L.; Hansen, E.W. Considerations regarding use of solvents in in vitro cell based assays. Cytotechnology 2013, 65, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Hollebeeck, S.; Raas, T.; Piront, N.; Schneider, Y.J.; Toussaint, O.; Larondelle, Y.; During, A. Dimethyl sulfoxide (DMSO) attenuates the inflammatory response in the in vitro intestinal Caco-2 cell model. Toxicol. Lett. 2011, 206, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Kloverpris, H.; Fomsgaard, A.; Handley, A.; Ackland, J.; Sullivan, M.; Goulder, P. Dimethyl sulfoxide (DMSO) exposure to human peripheral blood mononuclear cells (PBMCs) abolish T cell responses only in high concentrations and following coincubation for more than two hours. J. Immunol. Methods 2010, 356, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Misch, D.W.; Misch, M.S. Dimethyl sulfoxide: Activation of lysosomes in vitro. Proc. Natl. Acad. Sci. USA 1967, 58, 2463–2467. [Google Scholar] [CrossRef]
- Wedner, H.J.; Bass, G. Induction of the tyrosine phosphorylation of a 66 KD soluble protein by DMSO in human peripheral blood T lymphocytes. Biochem. Biophys. Res. Commun. 1986, 140, 743–749. [Google Scholar] [CrossRef]
- Kloesch, B.; Liszt, M.; Broell, J.; Steiner, G. Dimethyl sulphoxide and dimethyl sulphone are potent inhibitors of IL-6 and IL-8 expression in the human chondrocyte cell line C-28/I2. Life Sci. 2011, 89, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Abbruzzese, L.; Agostini, F.; Durante, C.; Toffola, R.T.; Rupolo, M.; Rossi, F.M.; Lleshi, A.; Zanolin, S.; Michieli, M.; Mazzucato, M. Long term cryopreservation in 5% DMSO maintains unchanged CD34(+) cells viability and allows satisfactory hematological engraftment after peripheral blood stem cell transplantation. Vox Sang. 2013, 105, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Thibeault, S. Effect of DMSO concentration, cell density and needle gauge on the viability of cryopreserved cells in three dimensional hyaluronan hydrogel. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2013, 2013, 6228–6231. [Google Scholar] [CrossRef] [PubMed]
- Marthyn, P.; Beuscart, A.; Coll, J.; Moreau-Gachelin, F.; Righi, M. DMSO reduces CSF1 receptor levels and causes apoptosis in v-myc immortalized mouse macrophages. Exp. Cell Res. 1998, 243, 94–100. [Google Scholar] [CrossRef] [PubMed]
- León-García, M.C.; Ríos-Castro, E.; López-Romero, E.; Cuéllar-Cruz, M. Evaluation of cell wall damage by dimethyl sulfoxide in Candida species. Res. Microbiol. 2017, 168, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Liu, M.; Luo, Q.; Zhuo, H.; Cao, H.; Wang, J.; Han, Y. Toxic effects of dimethyl sulfoxide on red blood cells, platelets, and vascular endothelial cells in vitro. FEBS Open Bio 2017, 7, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Ansel, H.C.; Leake, W.F. Hemolysis of erythrocytes by antibacterial preservatives III: Influence of dimethyl sulfoxide on the hemolytic activity of phenol. J. Pharm. Sci. 1966, 55, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Notman, R.; Noro, M.; O’malley, B.; Anwar, J. Molecular Basis for Dimethylsulfoxide (DMSO) Action on Lipid Membranes. J. Am. Chem. Soc. 2006, 128, 13982–13983. [Google Scholar] [CrossRef] [PubMed]
- Boyman, J.F.; Purton, C.D.; Surh, J.; Sprent, J. Cytokines and T-cell homeostasis. Curr. Opin. Immunol. 2007, 19, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, T.; Bandheka, L.P.; Sainis, K.B. Immunomodulatory effect of Tylophoraindica on Con A induced lymphoproliferation. Phytomedicine 2001, 8, 431–437. [Google Scholar] [CrossRef]
- Malek, T.R. The biology of interleukin-2. Annu. Rev. Immunol. 2008, 26, 453–479. [Google Scholar] [CrossRef] [PubMed]
- Gharagozloo, M.; Velardi, E.; Bruscoli, S.; Agostini, M.; Di Sante, M.; Donato, V.; Amirghofran, Z.; Riccardi, C. Silymarin suppress CD4+ T cell activation and proliferation: effects on NF-κB activity and IL-2 production. Pharmacol. Res. 2010, 61, 405–409. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.G.; Almeida, V.G.; Avelar-Freitas, B.A.; Grael, C.F.F.; Gregório, L.E.; Pereira, W.F.; Brito-Melo, G.E.A. Phytochemical screening of the dichloromethane–ethanolic extract of Eriosema campestre var. macrophylum roots and its antiproliferative effect on human peripheral blood lymphocytes. Rev. Bras. Farmacogn. 2016, 26, 464–470. [Google Scholar] [CrossRef]
- Zhang, X.Q.; Eyzaguirre, C. Effects of hypoxia induced by Na2S2O4 on intracellular calcium and resting potential of mouse glomus cells. Brain Res. 1999, 818, 118–126. [Google Scholar] [CrossRef]
- Medeiros, M.C.; Frasnelli, S.C.; Bastos Ade, S.; Orrico, S.R.; Rossa, C., Jr. Modulation of cell proliferation, survival and gene expression by RAGE and TLR signaling in cells of the innate and adaptive immune response: role of p38 MAPK and NF-κB. J. Appl. Oral Sci. 2014, 22, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Tonello, S.; Rizzi, M.; Miqliario, M.; Rocchetti, V.; Reno, F. Low concentrations of neutrophil extracellular traps induce proliferation in human keratinocytes via NF-kB activation. J. Dermatol. Sci. 2017, 88, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim. Biophys. Acta 2014, 1843, 2563–2582. [Google Scholar] [CrossRef] [PubMed]
- Avelar-Freitas, B.A.; Almeida, V.G.; Santos, M.G.; Santos, J.A.T.; Barroso, P.R.; Grael, C.F.F.; Gregório, L.E.; Rocha-Vieira, E.; Brito-Melo, G.E.A. Essential oil from Ageratum fastigiatum reduces expression of the pro-inflammatory cytokine tumor necrosisfactor-alpha in peripheral blood leukocytes subjected to in vitro stimulation with phorbol myristateacetate. Rev. Bras. Farmacogn. 2015, 25, 129–133. [Google Scholar] [CrossRef]
- Elisia, I.; Nakamura, H.; Lam, V.; Hofs, E.; Cederberg, R.; Cait, J.; Hughes, M.R.; Lee, L.; Jia, W.; Adomat, H.H.; et al. DMSO represses inflammatory cytokyne production from human blood cells and reduces autoimmune arthritis. PLoS ONE 2016, 11, e0152538. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.J.; Sytwu, H.K.; Yu, J.C.; Chen, Y.W.; Kuo, Y.L.; Yu, C.C.; Chang, H.M.; Chan, D.C.; Huang, S.H. Dimethyl sulfoxide inhibits spontaneous diabetes and autoimmune recurrence in non-obese diabetic mice by inducing differentiation of regulatory T cells. Toxicol. Appl. Pharmacol. 2015, 28, 2207–2214. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.S.; Chen, C.L.; Huang, F.W.; Hou, W.H.; Huang, J.S. DMSO Enhances TGF-β Activity by Recruiting the Type II TGF-β Receptor from Intracellular Vesicles to the Plasma Membrane. J. Cell. Biochem. 2015, 117, 1568–1579. [Google Scholar] [CrossRef] [PubMed]
- Bicalho, H.M.S.; Gontijo, M.C.; Nogueira-Machado, J.A. A simple technique for simultaneous human leukocytes separation. J. Immunol. Meth. 1981, 40, 115–116. [Google Scholar] [CrossRef]
- Avelar-Freitas, B.A.; Almeida, V.G.; Pinto, M.C.; Mourao, F.A.; Massensini, A.R.; Martins-Filho, O.A.; Rocha-Vieira, E.; Brito-Melo, G.E.A. Trypan blue exclusion assay by flow cytometry. Braz. J. Med. Biol. Res. 2014, 47, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Tossige-Gomes, R.; Avelar, N.C.; Simao, A.P.; Neves, C.D.; Brito-Melo, G.E.; Coimbra, C.C.; Rocha-Vieira, E.; Lacerda, A.C. Whole-body vibration decreases the proliferative response of TCD4(+) cells in elderly individuals with knee osteoarthritis. Braz. J. Med. Biol. Res. 2012, 45, 1262–1268. [Google Scholar] [CrossRef] [PubMed]
- Lyons, A.B.; Hasbold, J.; Hodgkin, P.D. Flow cytometric analysis of cell division history using dilution of carboxyfluorescein diacetate succinimidyl ester, a stably integrated fluorescent probe. Methods Cell Biol. 2001, 63, 375–398. [Google Scholar] [CrossRef] [PubMed]
- Angulo, R.; Fulcher, D.A. Measurement of Candida-specific blastogenesis: Comparison of carboxyfluorescein succinimidyl ester labelling of T cells, thymidine incorporation, and CD69 expression. Cytometry 1998, 34, 143–151. [Google Scholar] [CrossRef]
Sample Availability: Not available. |






© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Abreu Costa, L.; Henrique Fernandes Ottoni, M.; Dos Santos, M.G.; Meireles, A.B.; Gomes de Almeida, V.; De Fátima Pereira, W.; Alves de Avelar-Freitas, B.; Eustáquio Alvim Brito-Melo, G. Dimethyl Sulfoxide (DMSO) Decreases Cell Proliferation and TNF-α, IFN-γ, and IL-2 Cytokines Production in Cultures of Peripheral Blood Lymphocytes. Molecules 2017, 22, 1789. https://doi.org/10.3390/molecules22111789
De Abreu Costa L, Henrique Fernandes Ottoni M, Dos Santos MG, Meireles AB, Gomes de Almeida V, De Fátima Pereira W, Alves de Avelar-Freitas B, Eustáquio Alvim Brito-Melo G. Dimethyl Sulfoxide (DMSO) Decreases Cell Proliferation and TNF-α, IFN-γ, and IL-2 Cytokines Production in Cultures of Peripheral Blood Lymphocytes. Molecules. 2017; 22(11):1789. https://doi.org/10.3390/molecules22111789
Chicago/Turabian StyleDe Abreu Costa, Lucas, Marcelo Henrique Fernandes Ottoni, Michaelle Geralda Dos Santos, Agnes Batista Meireles, Valéria Gomes de Almeida, Wagner De Fátima Pereira, Bethânia Alves de Avelar-Freitas, and Gustavo Eustáquio Alvim Brito-Melo. 2017. "Dimethyl Sulfoxide (DMSO) Decreases Cell Proliferation and TNF-α, IFN-γ, and IL-2 Cytokines Production in Cultures of Peripheral Blood Lymphocytes" Molecules 22, no. 11: 1789. https://doi.org/10.3390/molecules22111789
APA StyleDe Abreu Costa, L., Henrique Fernandes Ottoni, M., Dos Santos, M. G., Meireles, A. B., Gomes de Almeida, V., De Fátima Pereira, W., Alves de Avelar-Freitas, B., & Eustáquio Alvim Brito-Melo, G. (2017). Dimethyl Sulfoxide (DMSO) Decreases Cell Proliferation and TNF-α, IFN-γ, and IL-2 Cytokines Production in Cultures of Peripheral Blood Lymphocytes. Molecules, 22(11), 1789. https://doi.org/10.3390/molecules22111789