Are Diatoms “Green” Aluminosilicate Synthesis Microreactors for Future Catalyst Production?

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Diatom Cultivation and Microscopic Characterization
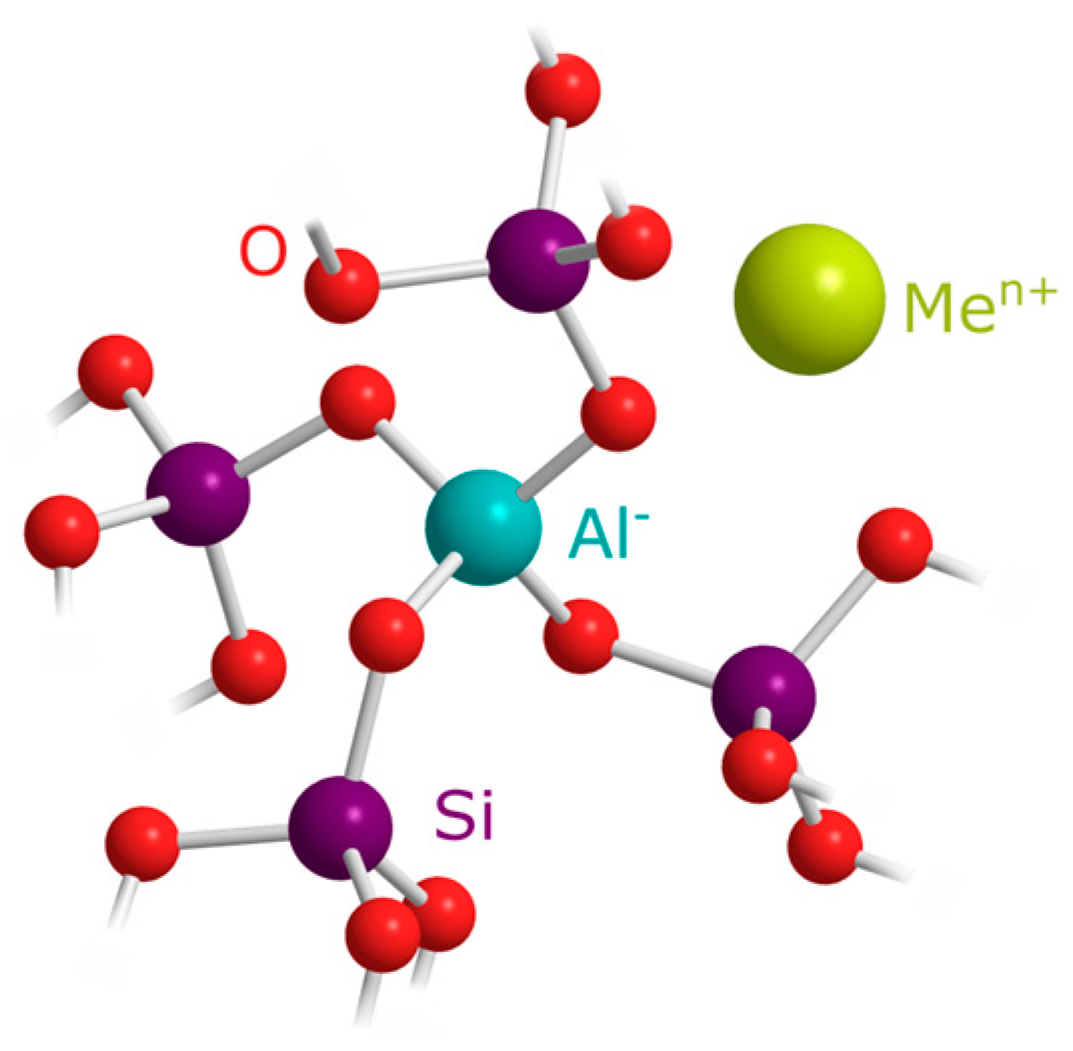
2.2. Chemical Structure and Elemental Composition
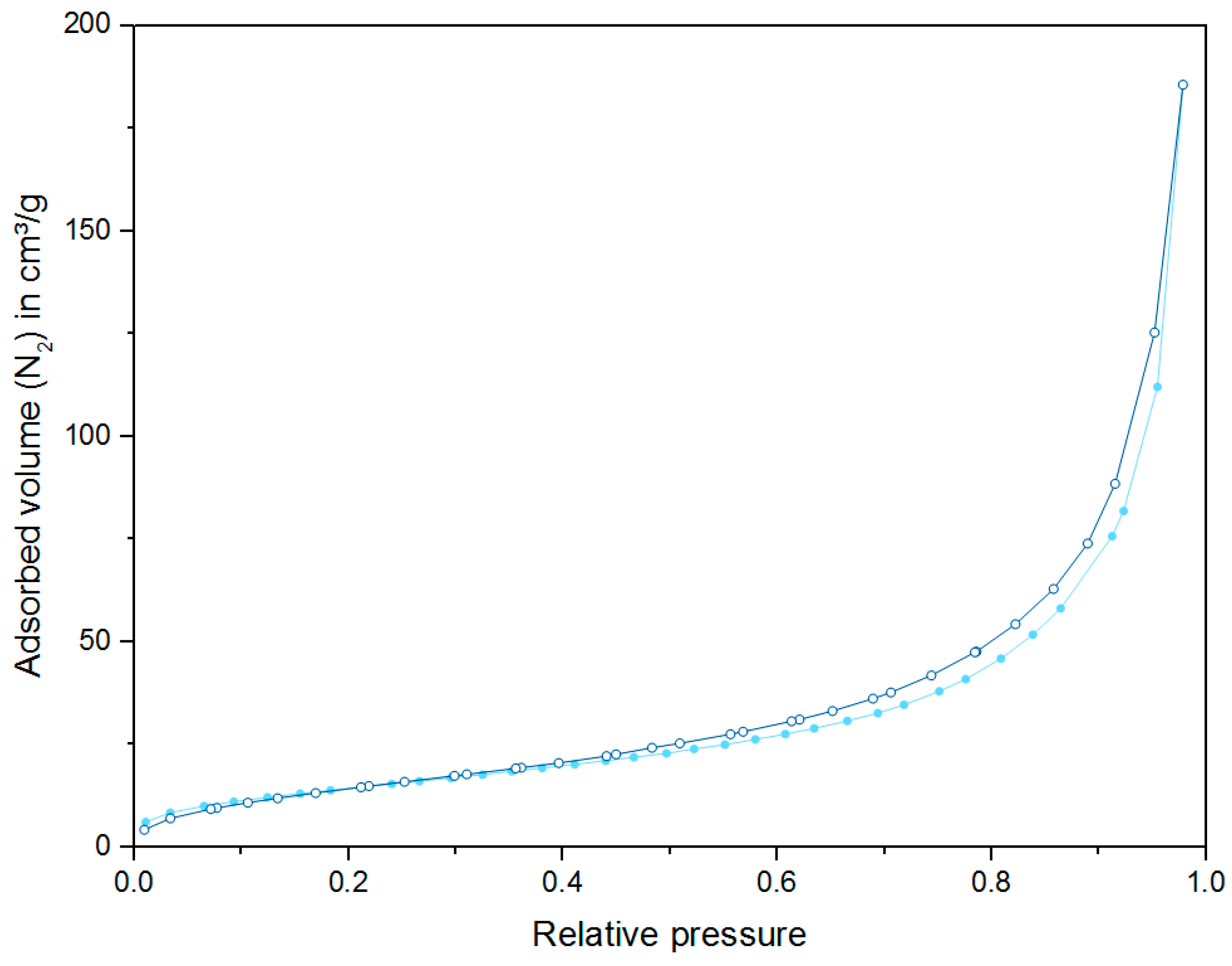
2.3. Nitrogen Physisorption
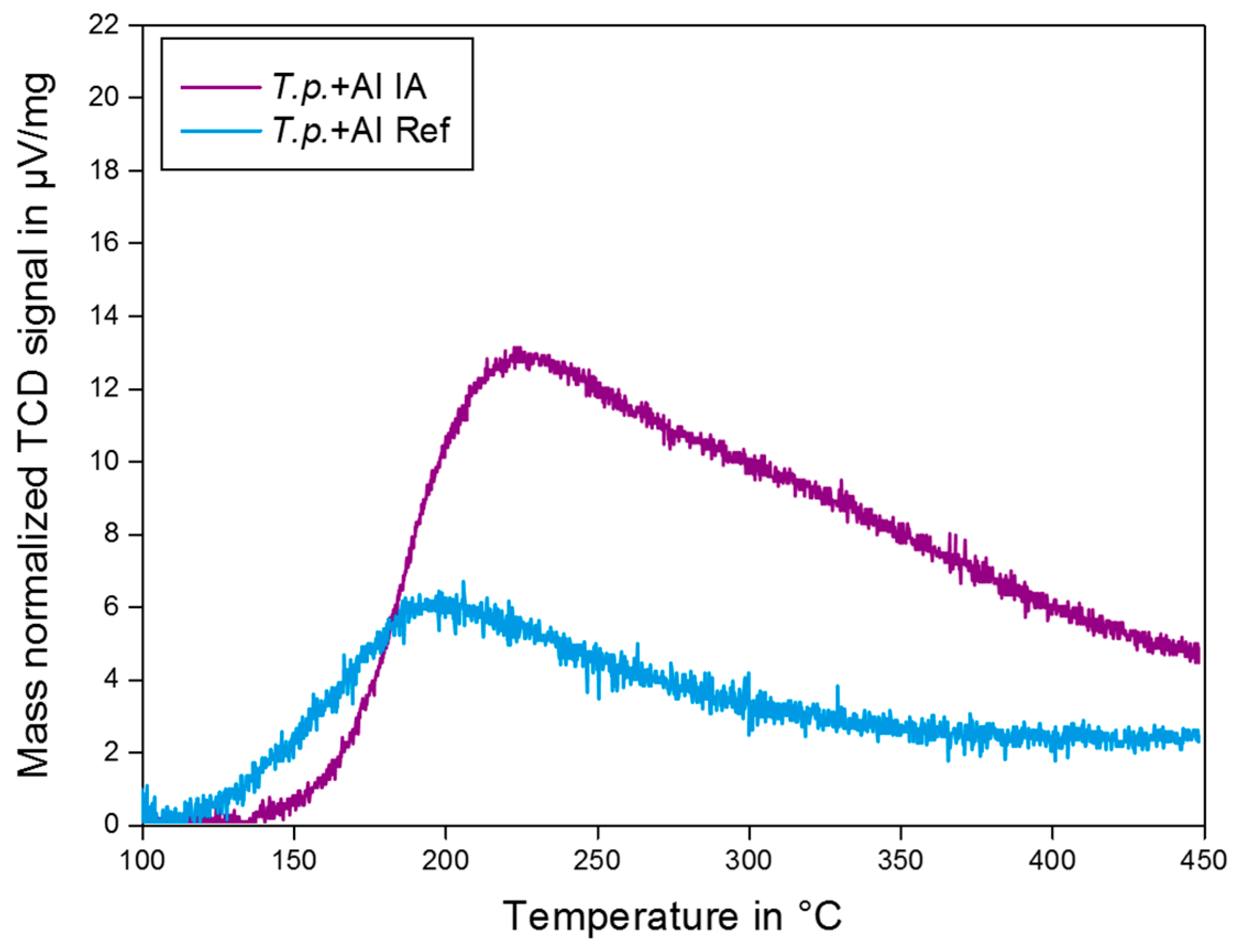
2.4. NH3 TPD
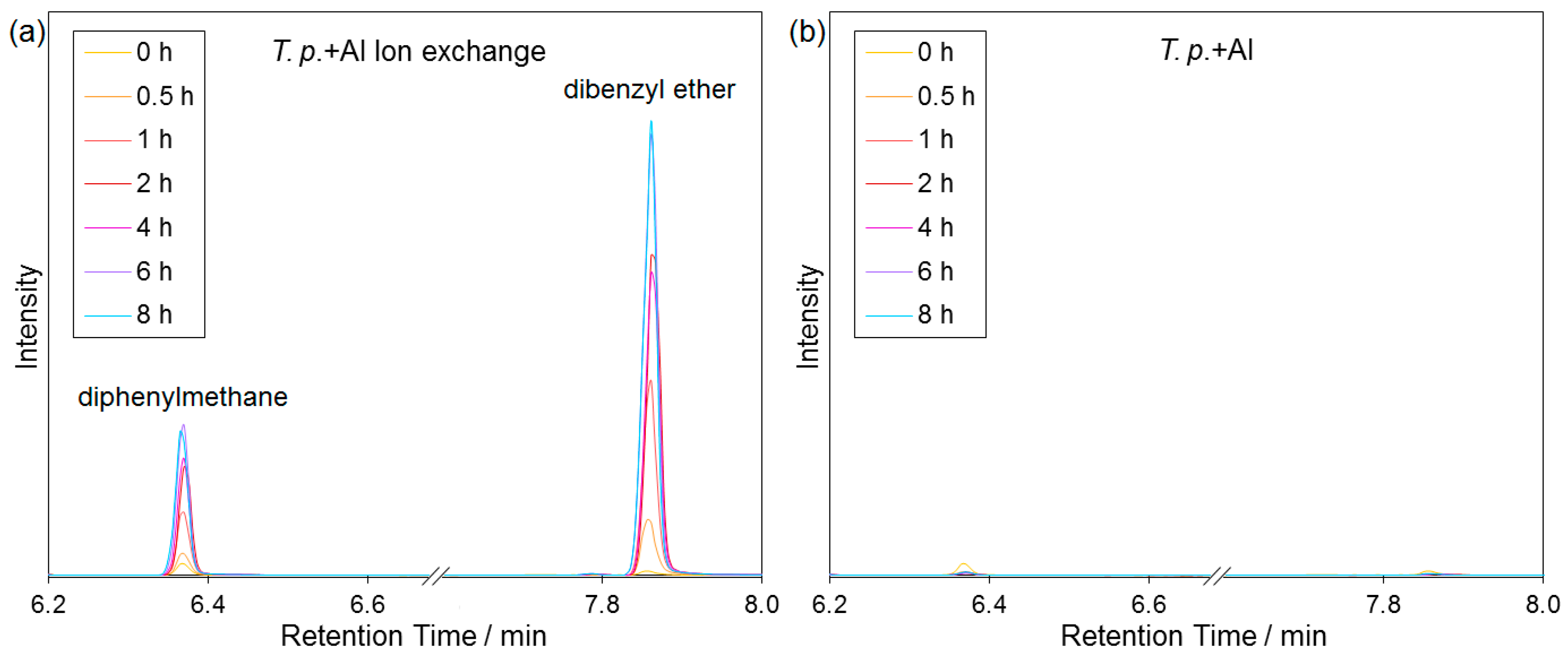
2.5. Catalytic Tests
3. Materials and Methods
3.1. Cultivation
3.2. Cleaning of the Frustules
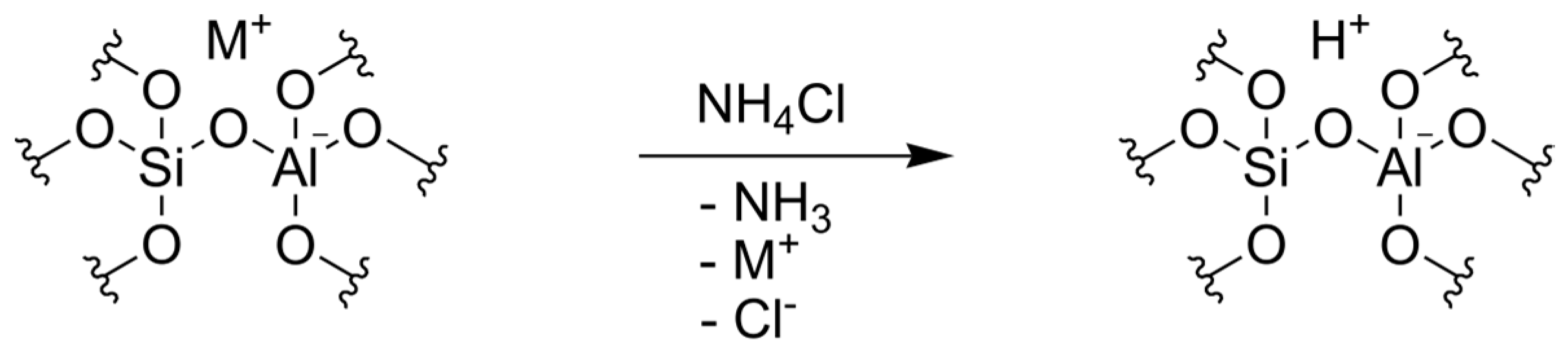
3.3. Ion Exchange
3.4. Scanning Electron Microscopy (SEM)
3.5. Inductively Coupled Plasma Optical Emission Spectroscopy (ICP-OES)
3.6. Nitrogen Physisorption
3.7. Temperature Programmed Desorption of Ammonia (NH3 TPD)
3.8. Catalytic Tests
3.9. Gas Chromatography Coupled to Mass Spectrometry (GC-MS)
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Corma, A. State of the Art and Future Challenges of Zeolites as Catalysts. J. Catal. 2003, 216, 298–312. [Google Scholar] [CrossRef]
- Strohmaier, K.G. Synthesis Approaches. In Zeolites and Catalysis—Synthesis, Reactions and Applications; Čejka, J., Corma, A., Zones, S., Eds.; Wiley-VCH: Weinheim, Germany, 2010; pp. 57–86. [Google Scholar]
- Davidovits, J. Geopolymer Chemistry and Applications, 2nd ed.; Institut Géopolymère: Saint-Quentin, France, 2008. [Google Scholar]
- Werner, D. The Biology of Diatoms; University of California Press: Berkeley, CA, USA; Los Angeles, CA, USA, 1977; Volume 13. [Google Scholar]
- Round, F.E.; Crawford, R.M.; Mann, D.G. Diatoms: Biology and Morphology of the Genera; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar]
- Mann, D.G. The species concept in diatoms. Phycologia 1999, 38, 437–495. [Google Scholar] [CrossRef]
- Smol, J.P.; Stoermer, E.F. The Diatoms: Applications for the Environmental and Earth Sciences, 2nd ed.; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Kogel, J.E.; Trivedi, N.C.; Barker, J.M.; Krukowski, S.T. (Eds.) Industrial Minerals & Rocks: Commodities, Markets, and Uses, 7th ed.; Society for Mining, Metallurgy, and Exploration: Littleton, CO, USA, 2006. [Google Scholar]
- Fischer, C.; Adam, M.; Mueller, A.C.; Sperling, E.; Wustmann, M.; van Pée, K.-H.; Kaskel, S.; Brunner, E. Gold Nanoparticle-Decorated Diatom Biosilica: A Favorable Catalyst for the Oxidation of d-Glucose. ACS Omega 2016, 1, 1253–1261. [Google Scholar] [CrossRef]
- Loucaides, S.; Behrends, T.; Van Cappellen, P. Reactivity of biogenic silica: Surface versus bulk charge density. Geochim. Cosmochim. Acta 2010, 74, 517–530. [Google Scholar] [CrossRef]
- Sverjensky, D.A.; Sahai, N. Theoretical prediction of single-site surface-protonation equilibrium constants for oxides and silicates in water. Geochim. Cosmochim. Acta 1996, 60, 3773–3797. [Google Scholar] [CrossRef]
- Losic, D.; Yu, Y.; Aw, M.S.; Simovic, S.; Thierry, B.; Addai-Mensah, J. Surface functionalisation of diatoms with dopamine modified iron-oxide nanoparticles: Toward magnetically guided drug microcarriers with biologically derived morphologies. Chem. Commun. 2010, 46, 6323–6325. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lu, Y.; Chen, R.; Ma, L.; Jiang, Y.; Wang, H. Lead Ions Sorption from Waste Solution Using Aluminum Hydroxide Modified Diatomite. J. Environ. Prot. 2014, 5, 509–516. [Google Scholar] [CrossRef]
- Van Eynde, E.; Hu, Z.-Y.; Tytgat, T.; Verbruggen, S. Diatom silica–titania photocatalysts for air purification by bio-accumulation of different titanium sources. Environ. Sci. Nano 2016, 3, 1052–1061. [Google Scholar] [CrossRef]
- Lang, Y.; Del Monte, F.; Collins, L.; Rodriguez, B.J.; Thompson, K.; Dockery, P.; Finn, D.P.; Pandit, A. Functionalization of the living diatom Thalassiosira weissflogii with thiol moieties. Nat. Commun. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Kucki, M. Biological Photonic Crystals: Diatoms. Dye Functionalization of Biological Silica Nanostructures. Ph.D. Thesis, University of Kassel, Kassel, Germany, 2009. [Google Scholar]
- Kröger, N.; Sumper, M. Diatom Cell Wall Proteins and the Cell Biology of Silica Biomineralization. Protist 1998, 149, 213–219. [Google Scholar] [CrossRef]
- Sumper, M.; Kröger, N. Silica formation in diatoms: The function of long-chain polyamines and silaffins. J. Mater. Chem. 2004, 14, 2059–2065. [Google Scholar] [CrossRef]
- Hedrich, R.; Machill, S.; Brunner, E. Biomineralization in diatoms—Phosphorylated saccharides are part of Stephanopyxis turris biosilica. Carbohydr. Res. 2013, 365, 2–60. [Google Scholar] [CrossRef] [PubMed]
- Ellwood, M.J.; Hunter, K.A. The incorporation of zinc and iron into the frustule of the marine diatom Thalassiosira pseudonana. Limnol. Oceanogr. 2000, 45, 1517–1524. [Google Scholar] [CrossRef]
- Azam, F.; Hemmingsen, B.B.; Volcani, B.E. Germanium incorporation into the silica of diatom cell walls. Arch. Mikrobiol. 1973, 92, 11–20. [Google Scholar] [CrossRef]
- Godinho, R.M.; Cabrita, M.T.; Alves, L.C.; Pinheiro, T. Changes of the elemental distributions in marine diatoms as a reporter of sample preparation artefacts. A nuclear microscopy application. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. At. 2015, 348, 265–268. [Google Scholar] [CrossRef]
- Machill, S.; Köhler, L.; Ueberlein, S.; Hedrich, R.; Kunaschk, M.; Paasch, S.; Schulze, R.; Brunner, E. Analytical studies on the incorporation of aluminium in the cell walls of the marine diatom Stephanopyxis turris. BioMetals 2013, 26, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Van Beusekom, J.E.E.; Weber, A. Decreasing diatom abundance in the North Sea: The possible significance of aluminum. In Marine Eutrophication and Population Dynamics: 25th European Marine Biology Symposium; Olsen & Olsen: Fredensborg, Denmark, 1992. [Google Scholar]
- Ren, J.-L.; Zhang, G.-L.; Zhang, J.; Shi, J.-H.; Liu, S.-M.; Li, F.-M.; Jin, J.; Liu, C.-G. Distribution of dissolved aluminum in the Southern Yellow Sea: Influences of a dust storm and the spring bloom. Mar. Chem. 2011, 125, 69–81. [Google Scholar] [CrossRef]
- Gensemer, R.W.; Playle, R.C. The Bioavailability and Toxicity of Aluminum in Aquatic Environments. Crit. Rev. Environ. Sci. Technol. 1999, 29, 315–450. [Google Scholar] [CrossRef]
- Xie, J.; Bai, X.; Lavoie, M.; Lu, H.; Fan, X.; Pan, X.; Fu, Z.; Qian, H. Analysis of the Proteome of the Marine Diatom Phaeodactylum tricornutum Exposed to Aluminum Providing Insights into Aluminum Toxicity Mechanisms. Environ. Sci. Technol. 2015, 49, 11182–11190. [Google Scholar] [CrossRef] [PubMed]
- Exley, C.; Tollervey, A.; Gray, G.; Roberts, S.; Birchall, J.D. Silicon, aluminium and the biological availability of phosphorus in algae. Proc. R. Soc. Lond. B Biol. Sci. 1993, 253, 93–99. [Google Scholar] [CrossRef]
- Wustman, B.A.; Gretz, M.R.; Hoagland, K.D. Extracellular Matrix Assembly in Diatoms (Bacillariophyceae) (I. A Model of Adhesives Based on Chemical Characterization and Localization of Polysaccharides from the Marine Diatom Achnanthes longipes and Other Diatoms). Plant Physiol. 1997, 113, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- Geresh, S.; Arad, S.; Shefer, A. Chemically crosslinked polysaccharide of the red microalga Rhodella Reticulata—An ion exchanger for toxic metal ions. J. Carbohydr. Chem. 1997, 16, 703–708. [Google Scholar] [CrossRef]
- Iler, R.K. Effect of adsorbed alumina on the solubility of amorphous silica in water. J. Colloid Interface Sci. 1973, 43, 399–408. [Google Scholar] [CrossRef]
- Lewin, J.C. The dissolution of silica from diatom walls. Geochim. Cosmochim. Acta 1961, 21, 182–198. [Google Scholar] [CrossRef]
- Koning, E.; Gehlen, M.; Flank, A.-M.; Calas, G.; Epping, E. Rapid post-mortem incorporation of aluminum in diatom frustules: Evidence from chemical and structural analyses. Mar. Chem. 2007, 106, 208–222. [Google Scholar] [CrossRef]
- Stone, W.E.E.; El Shafei, G.M.S.; Sanz, J.; Selim, S.A. Association of soluble aluminum ionic species with a silica-gel surface: A solid-state NMR study. J. Phys. Chem. 1993, 97, 10127–10132. [Google Scholar] [CrossRef]
- Gensemer, R.W.; Smith, R.E.H.; Duthie, H.C.; Schiff, S.L. pH Tolerance and Metal Toxicity in Populations of the Planktonic Diatom Asterionella: Influences of Synthetic and Natural Dissolved Organic Carbon. Can. J. Fish. Aquat. Sci. 1993, 50, 121–132. [Google Scholar] [CrossRef]
- Zhou, L.; Tan, Y.; Huang, L.; Wang, W.-X. Enhanced utilization of organic phosphorus in a marine diatom Thalassiosira weissflogii: A possible mechanism for aluminum effect under P limitation. J. Exp. Mar. Biol. Ecol. 2016, 478, 77–85. [Google Scholar] [CrossRef]
- Turnau, K.; Henriques, F.; Wolowski, K. Differences in Metal Distribution and Concentration in Algal Species Living in a Highly Acidic, Metal-rich Pond of a Pyrite Mine in Portugal. Acta Protozool. 2009, 48, 339–343. [Google Scholar]
- Armbrust, E.V.; Berges, J.A.; Bowler, C.; Green, B.R.; Martinez, D.; Putnam, N.H.; Zhou, S.; Allen, A.E.; Apt, K.E.; Bechner, M.; et al. The Genome of the Diatom Thalassiosira pseudonana: Ecology, Evolution, and Metabolism. Science 2004, 306, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Bucciarelli, E.; Sunda, W.G. Influence of CO2, nitrate, phosphate, and silicate limitation on intracellular dimethylsulfoniopropionate in batch cultures of the coastal diatom Thalassiosira pseudonana. Limnol. Oceanogr. 2003, 48, 2256–2265. [Google Scholar] [CrossRef]
- Fisher, N.S. On the Differential Sensitivity of Estuarine and Open-Ocean Diatoms to Exotic Chemical Stress. Am. Nat. 1977, 111, 871–895. [Google Scholar] [CrossRef]
- Rijstenbil, J.W.; Sandee, A.; Van Drie, J.; Wijnholds, J.A. Interaction of toxic trace metals and mechanisms of detoxification in the planktonic diatoms Ditylum brightwellii and Thalassiosira pseudonana. FEMS Microbiol. Rev. 1994, 14, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Hinga, K.R. Effects of pH on coastal marine phytoplankton. Mar. Ecol. Prog. Ser. 2002, 238, 281–300. [Google Scholar] [CrossRef]
- Guillard, R.R.L.; Ryther, J.H. Studies of marine planctonic Diatoms: I. Cyclotella nana Hustedt, and Detonula confervacea (Cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Sobrino, C.; Neale, P.J. Short-Term and Long-Term Effects of Temperature on Photosynthesis in the Diatom Thalassiosira pseudonana under UVR Exposures. J. Phycol. 2007, 43, 426–436. [Google Scholar] [CrossRef]
- Beck, L.; Gehlen, M.; Flank, A.-M.; Van Bennekom, A.J.; Van Beusekom, J.E.E. The relationship between Al and Si in biogenic silica as determined by PIXE and XAS. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. At. 2002, 189, 180–184. [Google Scholar] [CrossRef]
- Mazumder, N.; Gogoi, A.; Buragohain, A.K.; Choudhury, A. Structural and optical characterization of fresh water diatoms (Cyclotella sp.): Nature’s nanoporous silica manufacturing plant. In Proceedings of the Quantum Dots and Nanostructures: Synthesis, Characterization, and Modeling XI, San Diego, CA, USA, 1–6 February 2014. [Google Scholar]
- Gehlen, M.; Beck, L.; Calas, G.; Flank, A.-M.; Van Bennekom, A.J.; Van Beusekom, J.E.E. Unraveling the atomic structure of biogenic silica: Evidence of the structural association of Al and Si in diatom frustules. Geochim. Cosmochim. Acta 2002, 66, 1601–1609. [Google Scholar] [CrossRef]
- Kern, C.; Jess, A. Verkokung und Koksabbrand in heterogenen Katalysatoren. Chemie Ingenieur Technik 2006, 78, 1033–1048. [Google Scholar] [CrossRef]
- Misra, D.N.; Bowen, R.L.; Mattamal, G.J. Surface Area of Dental Enamel, Bone, and Hydroxyapatite: Chemisorption from Solution. Calcif. Tissue Res. 1978, 26, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Thommes, M.; Kaneko, K.; Neimark, A.V.; Olivier, J.P.; Rodriguez-Reinoso, F.; Rouquerol, J.; Sing, K.S.W. Physisorption of Gases, with Special Reference to the Evaluation of Surface Area and Pore Size Distribution (IUPAC Technical Report). Pure Appl. Chem. 2015, 87, 1051–1069. [Google Scholar] [CrossRef]
- Katada, N.; Niwa, M. Analysis of Acidic Properties of Zeolitic and Non-Zeolitic Solid Acid Catalysts Using Temperature-Programmed Desorption of Ammonia. Catal. Surv. Asia 2004, 8, 161–170. [Google Scholar] [CrossRef]
- Macquarrie, D.J. Industrial Friedel-Crafts Chemistry. In Catalytic Asymmetric Friedel-Crafts Alkylations; Wiley-VCH Verlag GmbH: Weinheim, Germany, 2009; pp. 271–288. [Google Scholar]
- Krauch, H.; Kunz, W.; Nonnenmacher, E. Reaktionen der Organischen Chemie, 6th ed.; WILEY-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2009; pp. 1–154. [Google Scholar]
- Narender, N.; Krishna Mohan, K.V.V.; Kulkarni, S.J.; Ajit Kumar Reddy, I. Liquid phase benzylation of benzene and toluene with benzyl alcohol over modified zeolites. Catal. Commun. 2006, 7, 583–588. [Google Scholar] [CrossRef]
- Verboekend, D.; Pérez-Ramírez, J. Design of Hierarchical Zeolite Catalysts by Desilication. Catal. Sci. Technol. 2011, 1, 879–890. [Google Scholar] [CrossRef]
- Driscoll, C.T.; Schecher, W.D. The chemistry of aluminum in the environment. Environ. Geochem. Health 1990, 12, 28–49. [Google Scholar] [CrossRef] [PubMed]
- Gensemer, R.W. Role of Aluminum and Growth Rate on Changes in Cell Size and Silica Content of Silica-Limited Populations of Asterionella Ralfsii var. Americana (Bacillariophyceae). J. Phycol. 1990, 26, 250–258. [Google Scholar] [CrossRef]
- Gillmore, M.L.; Golding, L.A.; Angel, B.M.; Adams, M.S.; Jolley, D.F. Toxicity of dissolved and precipitated aluminium to marine diatoms. Aquat. Toxicol. 2016, 174, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.-Y.; Chen, L.-H.; Li, Y.; Rooke, J.C.; Sanchez, C.; Su, B.-L. Hierarchically porous materials: Synthesis strategies and structure design. Chem. Soc. Rev. 2017, 46, 481–558. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.; Holmes, S.; Hanif, N.; Cundy, C. Hierarchical Pore Structures through Diatom Zeolitization. Angew. Chem. 2000, 112, 1521–3757. [Google Scholar] [CrossRef]
- Fowler, C.E.; Hoog, Y.; Vidal, L.; Lebeau, B. Mesoporosity in diatoms via surfactant induced silica rearrangement. Chem. Phys. Lett. 2004, 398, 414–417. [Google Scholar] [CrossRef]
- Georgianna, D.R.; Mayfield, S.P. Exploiting diversity and synthetic biology for the production of algal biofuels. Nature 2012, 488, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Hannon, M.; Gimpel, J.; Tran, M.; Rasala, B.; Mayfield, S. Biofuels from algae: Challenges and potential. Biofuels 2010, 1, 763–784. [Google Scholar] [CrossRef] [PubMed]
- Selzer, C.; Biemelt, T.; Werner, A.; Kaskel, S. Hierarchical zeolite ZSM-58 as shape selective catalyst for methanol-to-olefins reaction. Microporous Mesoporous Mater. 2018, 261, 51–57. [Google Scholar] [CrossRef]
- Thompson, P. The response of growth and biochemical composition to variations in daylength, temperature, and irradiance in the marine diatom Thalassiosira pseudonana (Bacillariophyceae). J. Phycol. 1999, 35, 1215–1223. [Google Scholar] [CrossRef]
- Verity, P.G. Effects of temperature, irradiance, and daylength on the marine diatom leptocylindrus danicus cleve. I. Photosynthesis and cellular composition. J. Exp. Mar. Biol. Ecol. 1981, 55, 79–91. [Google Scholar] [CrossRef]
- Harrison, P.J.; Waters, R.E.; Taylor, F.J.R. A Broad Spectrum Artificial Sea Water Medium for Coastal and Open Ocean Phytoplankton. J. Phycol. 1980, 16, 28–35. [Google Scholar] [CrossRef]
- Nordstrom, D.K. Aqueous equilibrium data for mononuclear aluminum species. In The Environmental Chemistry of Aluminum, 2nd ed.; Sposito, G., Ed.; CRC Press: Boca Raton, FL, USA, 1995; pp. 39–80. [Google Scholar]
- Iler, R.K. The Chemistry of Silica: Solubility, Polymerization, Colloid and Surface Properties and Biochemistry of Silica; Wiley: New York, NY, USA, 1979. [Google Scholar]
Sample Availability: Small amounts of biosilica (reference and aluminum enriched) could be reproduced and would then be available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T.p. Reference | T.p. + Al | T.p. + Al + Ion Exchange | ||||
|---|---|---|---|---|---|---|
| Element | n(x):n(Si) | n(x):n(Si) | n(x):n(Si) | |||
| Al | 1.66 | 1:603 | 21.4 | 1:47 | 16.3 | 1:61 |
| Na | 39.1 | 1:26 | 58.2 | 1:17 | 7.35 | 1:136 |
| K | 0.307 | 1:3250 | 5.29 | 1:189 | 0.239 | 1:4190 |
| Ca | 0.451 | 1:2220 | 1.40 | 1:715 | 0.614 | 1:1630 |
| Mg | 2.58 | 1:387 | 0.828 | 1:1208 | 0.503 | 1:1990 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Köhler, L.; Machill, S.; Werner, A.; Selzer, C.; Kaskel, S.; Brunner, E. Are Diatoms “Green” Aluminosilicate Synthesis Microreactors for Future Catalyst Production? Molecules 2017, 22, 2232. https://doi.org/10.3390/molecules22122232
Köhler L, Machill S, Werner A, Selzer C, Kaskel S, Brunner E. Are Diatoms “Green” Aluminosilicate Synthesis Microreactors for Future Catalyst Production? Molecules. 2017; 22(12):2232. https://doi.org/10.3390/molecules22122232
Chicago/Turabian StyleKöhler, Lydia, Susanne Machill, Anja Werner, Carolin Selzer, Stefan Kaskel, and Eike Brunner. 2017. "Are Diatoms “Green” Aluminosilicate Synthesis Microreactors for Future Catalyst Production?" Molecules 22, no. 12: 2232. https://doi.org/10.3390/molecules22122232
APA StyleKöhler, L., Machill, S., Werner, A., Selzer, C., Kaskel, S., & Brunner, E. (2017). Are Diatoms “Green” Aluminosilicate Synthesis Microreactors for Future Catalyst Production? Molecules, 22(12), 2232. https://doi.org/10.3390/molecules22122232