
Stability of Bioactive Compounds in Broccoli as Affected by Cutting Styles and Storage Time

 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Effect of Cutting Style and Storage Time on the Content of Phenolic Compounds
2.2. Effect of Cutting Style and Storage Time on the Content of Glucosinolates
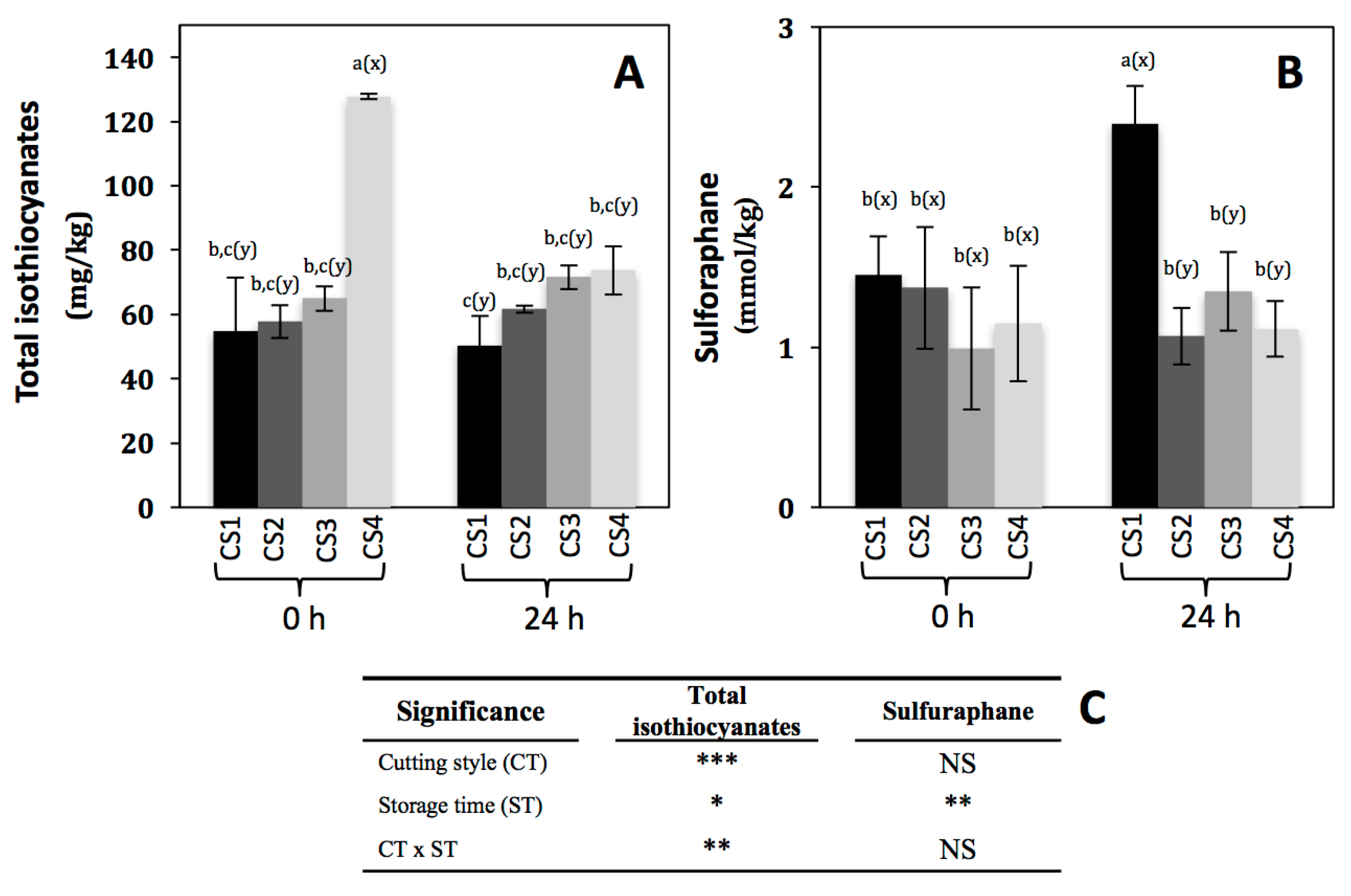
2.3. Effect of Cutting Style and Storage Time on the Content of Total Isothiocyanates and Sulforaphane
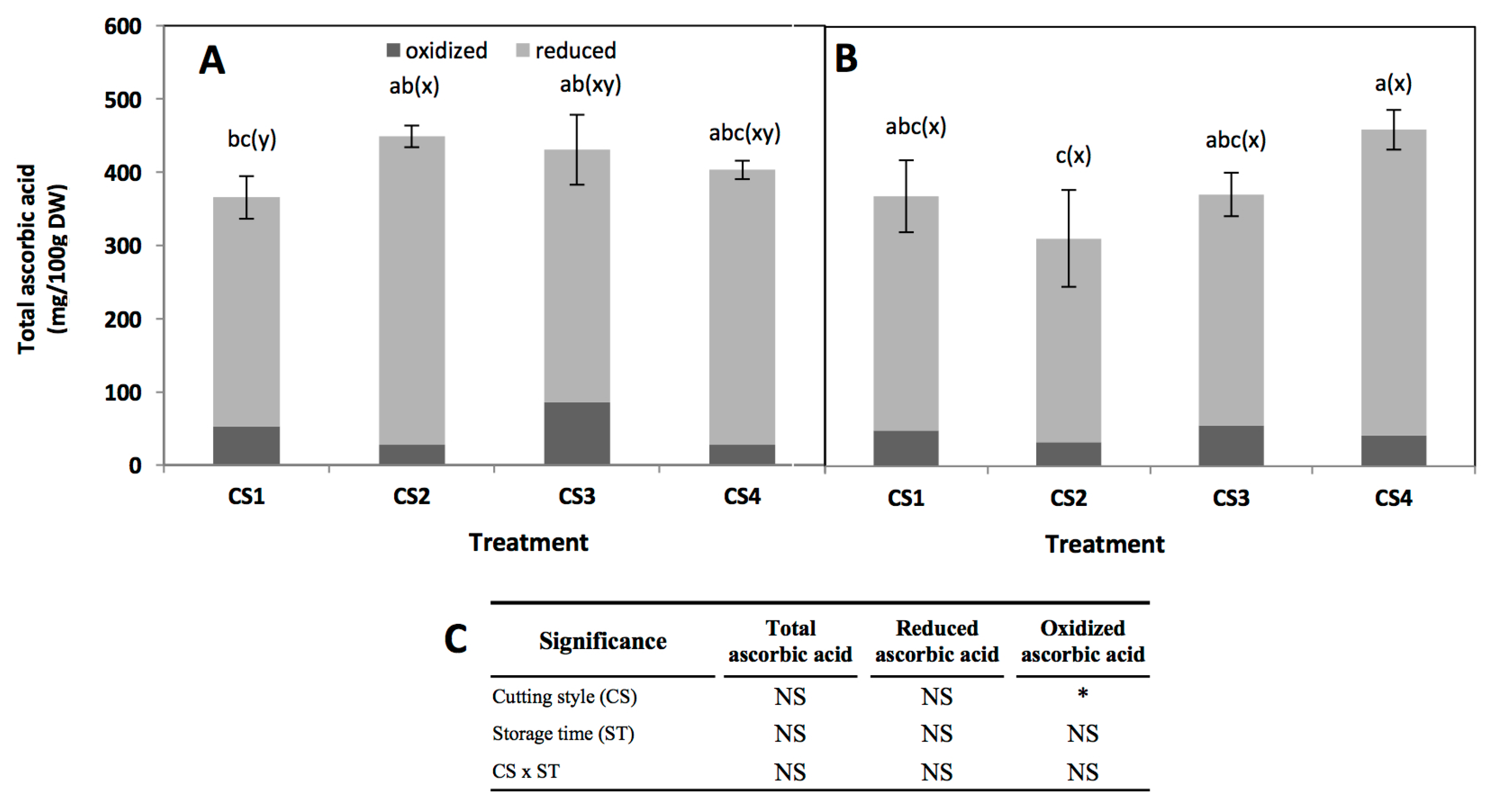
2.4. Effect of Cutting Style and Storage Time on Ascorbic Acid Content
3. Materials and Methods
3.1. Chemicals
3.2. Plant Material, Processing, and Storage Studies
3.3. Analysis of Phenolic Compounds by High-Performance Liquid Chromatography–Diode Array Detection and High-Performance Liquid Chromatography–Electrospray Ionization Mass Spectrometry
3.4. Analysis of Glucosinolates by High-Performance Liquid Chromatography–Diode Array Detection and High-Performance Liquid Chromatography–Electrospray Ionization Mass Spectrometry
3.5. Analysis of Total Isothiocyanates by Ciclocondensation with 1,2-bezenedithiol
3.6. Analysis of Sulforaphane
3.7. Extraction and Quantification of Reduced, Oxidized, and Total Ascorbate
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wells, H.F. United States Department of Agriculture. Available online: http://usda.mannlib.cornell.edu/usda/ers/VEGANDPULSESYEARBOOK/2016/VegetableandPulsesYearbook2016.pdf (accessed on 28 February 2017).
- Jacobo-Velázquez, D.A.; Cisneros-Zevallos, L. Correlations of antioxidant activity against phenolic content revisited: A new approach in data analysis for food and medicinal plants. J. Food Sci. 2009, 74, R107–R113. [Google Scholar] [CrossRef] [PubMed]
- Johnson, I.T. Glucosinolates: Bioavailability and importance to health. Int. J. Vitam. Nutr. Res. 2002, 72, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Gamet-Payrastre, L.; Li, P.; Lumeau, S.; Cassar, G.; Dupont, M.A.; Chevolleau, S.; Gasc, N.; Tulliez, J.; Tercé, F. Sulforaphane, a naturally occurring isothiocyanate, induces cell cycle arrest and apoptosis in HT29 human colon cancer cells. Cancer Res. 2000, 60, 1426–1433. [Google Scholar] [PubMed]
- Samaha, H.S.; Kelloff, G.J.; Steele, V.; Rao, C.V.; Reddy, B.S. Modulation of apoptosis by sulindac, curcumin, phenylethyl-3-methylcaffeate, and 6-phenylhexyl isothiocyanate: Apoptotic index as a biomarker in colon cancer chemoprevention and promotion. Cancer Res. 1997, 57, 1301–1305. [Google Scholar] [PubMed]
- Yu, R.; Mandlekar, S.; Harvey, K.J.; Ucker, D.S.; Kong, A.T. Chemopreventive isothiocyanates induce apoptosis and caspase-3-like protease activity. Cancer Res. 1998, 58, 402–408. [Google Scholar] [PubMed]
- Villarreal-García, D.; Jacobo-Velázquez, D.A. Glucosinolates from broccoli: Nutraceutical properties and their purification. Curr. Trends Nutraceuticals 2016, 1, 1–5. [Google Scholar]
- Yanaka, A. Sulforaphane enhances protection and repair of gastric mucosa against oxidative stress in vitro, and demonstrates anti-inflammatory effects on Helicobacter pylori infected gastric mucosae in mice and human subjects. Curr. Pharm. Des. 2011, 17, 1532–1540. [Google Scholar] [CrossRef] [PubMed]
- Heiss, E.; Herhaus, C.; Klimo, K.; Bartsch, H.; Gerhäuser, C. Nuclear factor κB is a molecular target for sulforaphane-mediated anti-inflammatory mechanisms. J. Biol. Chem. 2001, 276, 32008–32015. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Kader, A.A. Preharvest and postharvest factors influencing vitamin C content of horticultural crops. Postharvest Biol. Technol. 2000, 20, 207–220. [Google Scholar] [CrossRef]
- Frei, B. Reactive oxygen species and antioxidant vitamins: Mechanisms of action. Am. J. Med. 1994, 97, S5–S13. [Google Scholar] [CrossRef]
- Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A.; Pech, J.C.; Koiwa, H. Signaling Molecules Involved in the Postharvest Stress Response of Plants. In Handbook of Plant and Crop Physiology, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 259–276. [Google Scholar]
- Jacobo-Velázquez, D.A.; González-Agüero, M.; Cisneros-Zevallos, L. Cross-talk between signaling pathways: The link between plant secondary metabolite production and wounding stress response. Sci. Rep. 2015, 5, 8608. [Google Scholar] [CrossRef] [PubMed]
- Verkerk, R.; Dekker, M.; Jongen, W.M.F. Post-harvest increase of indolyl glucosinolates in response to chopping and storage of Brassica vegetables. J. Sci. Food Agric. 2001, 81, 953–958. [Google Scholar] [CrossRef]
- Villarreal-García, D.; Nair, V.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Plants as biofactories: Postharvest stress-induced accumulation of phenolic compounds and glucosinolates in broccoli subjected to wounding stress and exogenous phytohormones. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Barth, M.M.; Zhuang, H. Packaging design affects antioxidant vitamin retention and quality of broccoli florets during postharvest storage. Postharvest Biol. Technol. 1996, 9, 141–150. [Google Scholar] [CrossRef]
- Vallejo, F.; Tomás-Barberán, F.A.; Benavente-García, A.G.; García-Viguera, C. Total and individual glucosinolate contents in inflorescences of eight broccoli cultivars grown under various climatic and fertilisation conditions. J. Sci. Food Agric. 2003, 83, 307–313. [Google Scholar] [CrossRef]
- Torres-Contreras, A.M.; Nair, V.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Effect of exogenous amylolytic enzymes on the accumulation of chlorogenic acid isomers in wounded potato tubers. J. Agric. Food Chem. 2014, 62, 7671–7675. [Google Scholar] [CrossRef] [PubMed]
- Torres-Contreras, A.M.; Nair, V.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Plants as biofactories: Stress-induced production of chlorogenic acid isomers in potato tubers as affected by wounding intensity and storage time. Ind. Crops Prod. 2014, 62, 61–66. [Google Scholar] [CrossRef]
- Jacobo-Velázquez, D.A.; Martínez-Hernández, G.B.; Rodríguez, S.C.; Cao, C.-M.; Cisneros-Zevallos, L. Plants as biofactories: Physiological role of reactive oxygen species on the accumulation of phenolic antioxidants in carrot tissue under wounding and hyperoxia stress. J. Agric. Food Chem. 2011, 59, 6583–6593. [Google Scholar] [CrossRef] [PubMed]
- Reyes, L.F.; Villarreal, J.E.; Cisneros-Zevallos, L. The increase in antioxidant capacity after wounding depends on the type of fruit and vegetable tissue. Food Chem. 2007, 101, 1254–1262. [Google Scholar] [CrossRef]
- Lemmens, L.; Van Buggenhout, S.; Van Loey, A.M.; Hendrickx, M.E. Particle size reduction leading to cell wall rupture is more important for the β-carotene bioaccessibility of raw compared to thermally processed carrots. J. Agric. Food Chem. 2010, 58, 12769–12776. [Google Scholar] [CrossRef] [PubMed]
- Moelants, K.R.; Lemmens, L.; Vandebroeck, M.; Van Buggenhout, S.; Van Loey, A.M.; Hendrickx, M.E. Relation between particle size and carotenoid bioaccessibility in carrot-and tomato-derived suspensions. J. Agric. Food Chem. 2012, 60, 11995–12003. [Google Scholar] [CrossRef] [PubMed]
- Riehle, P.; Rusche, N.; Saake, B.; Rohn, S. Influence of the leaf content and herbal particle size on the presence and extractability of quantitated phenolic compounds in Cistus incanus herbal teas. J. Agric. Food Chem. 2014, 62, 10978–10988. [Google Scholar] [CrossRef] [PubMed]
- Rosa, N.N.; Barron, C.; Gaiani, C.; Dufour, C.; Micard, V. Ultra-fine grinding increases the antioxidant capacity of wheat bran. J. Cereal Sci. 2013, 57, 84–90. [Google Scholar] [CrossRef]
- Brewer, L.R.; Kubola, J.; Siriamornpun, S.; Herald, T.J.; Shi, Y.C. Wheat bran particle size influence on phytochemical extractability and antioxidant properties. Food Chem. 2014, 152, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; He, F.; Chen, G. Improving bioaccessibility and bioavailability of phenolic compounds in cereal grains through processing technologies: A concise review. J. Funct. Foods. 2014, 7, 101–111. [Google Scholar] [CrossRef]
- Saltveit, M.E. Wound induced changes in phenolic metabolism and tissue browning are altered by heat shock. Postharvest Biol. Technol. 2000, 21, 61–69. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085. [Google Scholar] [CrossRef] [PubMed]
- Becerra-Moreno, A.; Redondo-Gil, M.; Benavides, J.; Nair, V.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Combined effect of water loss and wounding stress on gene activation of metabolic pathways associated with phenolic biosynthesis in carrot. Front. Plant Sci. 2015, 6, 837. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gu, H.; Yu, H.; Zhao, Z.; Sheng, X.; Zhang, X. Genotypic variation of glucosinolates in broccoli (Brassica oleracea var. italica) florets from China. Food Chem. 2012, 133, 735–741. [Google Scholar] [CrossRef]
- Bhandari, S.R.; Jo, J.S.; Lee, J.G. Comparison of Glucosinolate Profiles in Different Tissues of Nine Brassica Crops. Molecules 2015, 20, 15827–15841. [Google Scholar] [CrossRef] [PubMed]
- Fahey, J.W.; Zhang, Y.; Talalay, P. Broccoli sprouts: An exceptionally rich source of inducers of enzymes that protect against chemical carcinogens. Proc. Natl. Acad. Sci. USA 1999, 94, 10367–10372. [Google Scholar] [CrossRef]
- Bones, A.M.; Rossiter, J.T. The myrosinase-glucosinolate system, its organisation and biochemistry. Physiol. Plant. 1996, 97, 194–208. [Google Scholar] [CrossRef]
- Yan, X.; Chen, S. Regulation of plant glucosinolate metabolism. Planta 2007, 226, 1343–1352. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, M.D.; Petersen, B.L.; Glawischnig, E.; Jensen, A.B.; Andreasson, E.; Halkier, B.A. Modulation of CYP79 genes and glucosinolate profiles in Arabidopsis by defense signaling pathways. Plant Physiol. 2003, 131, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Gigolashvili, T.; Berger, B.; Mock, H.P.; Müller, C.; Weisshaar, B.; Flügge, U.I. The transcription factor HIG1/MYB51 regulates indolic glucosinolate biosynthesis in Arabidopsis Thaliana. Plant J. 2007, 50, 886–901. [Google Scholar] [CrossRef] [PubMed]
- Constabel, C.P.; Ryan, C.A. A survey of wound-and methyl jasmonate-induced leaf polyphenol oxidase in crop plants. Phytochemistry 1998, 47, 507–511. [Google Scholar] [CrossRef]
- Starzyńska, A.; Leja, M.; Mareczek, A. Physiological changes in the antioxidant system of broccoli flower buds senescing during short-term storage, related to temperature and packaging. Plant Sci. 2003, 165, 1387–1395. [Google Scholar] [CrossRef]
- Fimognari, C.; Turrini, E.; Ferruzzi, L.; Lenzi, M.; Hrelia, P. Natural isothiocyanates: Genotoxic potential versus chemoprevention. Mutat. Res. 2012, 750, 107–131. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Liang, H.; Yuan, Q.; Wang, T.; Yan, X. Preparation and stability investigation of the inclusion complex of sulforaphane with hydroxypropyl-β-cyclodextrin. Carbohydr. Polym. 2010, 82, 613–617. [Google Scholar] [CrossRef]
- Koh, E.; Wimalasiri, K.M.S.; Chassy, A.W.; Mitchell, A.E. Content of ascorbic acid, quercetin, kaempferol and total phenolics in commercial broccoli. J. Food Comp. Anal. 2009, 22, 637–643. [Google Scholar] [CrossRef]
- Nicoletto, C.; Santagata, S.; Pino, S.; Sambo, P. Antioxidant characterization of different italian broccoli landraces. Hortic. Bras. 2016, 34, 74–79. [Google Scholar] [CrossRef]
- Suza, W.P.; Avila, C.A.; Carruthers, K.; Kulkarni, S.; Goggin, F.L.; Lorence, A. Exploring the impact of wounding and jasmonates on ascorbate metabolism. Plant Physiol. Biochem. 2010, 48, 337–350. [Google Scholar] [CrossRef] [PubMed]
- Ioannidi, E.; Kalamaki, M.S.; Engineer, C.; Pateraki, I.; Alexandrou, D.; Mellidou, I.; Giovannonni, J.; Kanellis, A.K. Expression profiling of ascorbic acid-related genes during tomato fruit development and ripening and in response to stress conditions. J. Exp. Bot. 2009, 60, 663–678. [Google Scholar] [CrossRef] [PubMed]
- Kiddle, G.; Bennett, R.N.; Botting, N.P.; Davidson, N.E.; Robertson, A.A.B.; Wallsgrove, R.M. High-performance liquid chromatographic separation of natural and synthetic desulphoglucosinolates and their chemical validation by UV, NMR and chemical ionisation-MS methods. Phytochem. Anal. 2001, 12, 226–242. [Google Scholar] [CrossRef] [PubMed]
- Clarke, D.B. Glucosinolates, structures and analysis in food. Anal. Methods 2010, 2, 310–325. [Google Scholar] [CrossRef]
- Zhang, Y.; Wade, K.L.; Prestera, T.; Talalay, P. Quantitative determination of isothiocyanates, dithiocarbamates, carbon disulfide, and related thiocarbonyl compounds by cyclocondensation with 1,2-benzenedithiol. Anal. Biochem. 1996, 239, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Ku, K.M.; Choi, J.H.; Kim, H.S.; Kushad, M.M.; Jeffery, E.H.; Juvik, J.A. Methyl jasmonate and 1-methylcyclopropene treatment effects on quinone reductase inducing activity and post-harvest quality of broccoli. PLoS ONE 2013, 8, e77127. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, K.M.; Ainsworth, E.A. Measurement of reduced, oxidized and total ascorbate content in plants. Nat. Protoc. 2007, 2, 871–874. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
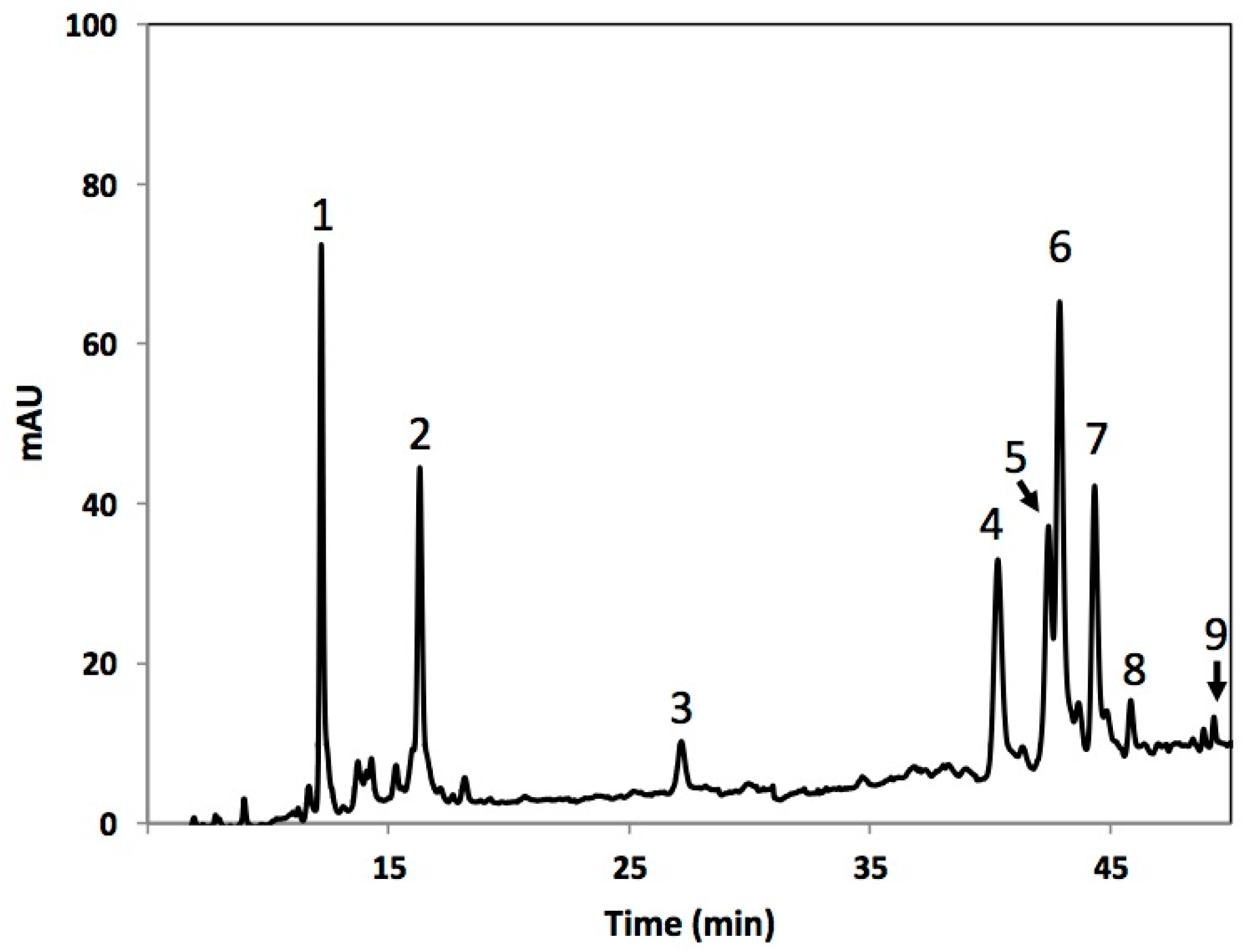
| Peak Number (Retention Time in min) | λ Max (nm) | Tentative Identification | [M − Z]− (m/z) | MS Fragments c |
|---|---|---|---|---|
| 1 (12.8) | 295, 320 | 3-O-CQA a,b | 353 | 179, 173, 135 |
| 2 (16.4) | 295, 320 | 5-O-CQA a,b | 353 | 172.5, 110.6, 92.7, 84.8 |
| 3 (27.5) | 225, 325 | CA b | 179 | 179, 161, 143, 133, 105 |
| 4 (40.6) | 235, 320 | 1-S-2-FG b | 723 | 449, 223 |
| 5 (42.7) | 215, 235, 320 | 1,2,2-TSG b | 959 | 735, 223 |
| 6 (43.1) | 235, 320 | 1,2-DFG b | 693 | 499, 175 |
| 7 (44.6) | 215, 235, 320 | 1,2-DS-2-FG b | 929 | 705, 223 |
| 8 (46.0) | 215, 235, 320 | 1-S-2,2-DFG b | 899 | 705, 223 |
| 9 (49.4) | 235, 320 | 1,2,2-TSPG b | 959 | 735, 223 |
| Storage Time | Cutting Style A | Phenolic Concentration (mg/kg DW) B,C,D,E | |||||||||
| 3-O-CQA | 5-O-CQA | CA | 1-S-2-FG | 1,2,2-TSG | |||||||
| 0 h | CS1 | 321.7 ± 17.42 | bc (x) | 180.5 ± 56.7 | bc (y) | 267.4 ± 14.4 | b (y) | 233.3 ± 16.9 | bc (x) | 250.8 ± 16.1 | abc (x) |
| CS2 | 414.8 ± 51.2 | ab (x) | 240.7 ± 15.9 | b (y) | 378.9 ± 12.8 | a (x) | 282.2 ± 46.8 | abc (x) | 294.4 ± 45.0 | abc (x) | |
| CS3 | 431.8 ± 58.7 | ab (x) | 171.9 ± 9.9 | bc (y) | 252.4 ± 63.8 | bc (y) | 289.0 ± 71.0 | abc (x) | 219.5 ± 34.7 | bc (x) | |
| CS4 | 353.8 ± 12.9 | b (x) | 401.5 ± 21.3 | a (x) | 161.3 ± 7.8 | cd (y) | 277.0 ± 5.6 | abc (x) | 267.0 ± 26.6 | abc (x) | |
| 24 h | CS1 | 472.0 ± 49.4 | a (x) | 357.8 ± 31.3 | a (x) | 114.1 ± 19.3 | d (z) | 369.9 ± 40.3 | a (x) | 359.4 ± 62.9 | a (x) |
| CS2 | 424.5 ± 42.5 | ab (x) | 143.4 ± 33.0 | c (y) | 267.6 ± 31.0 | b (y) | 308.5 ± 40.9 | ab (x) | 345.8 ± 57.3 | ab (xy) | |
| CS3 | 488.6 ± 0.5 | a (x) | 183.6 ± 30.6 | bc (y) | 368.9 ± 39.3 | a (x) | 304.4 ± 6.6 | ab (x) | 360.3 ± 6.8 | a (x) | |
| CS4 | 237.4 ± 11.8 | c (y) | 350.2 ± 15.2 | a (x) | 138.2 ± 6.6 | d (z) | 192.7 ± 6.0 | c (y) | 210.2 ± 9.0 | c (y) | |
| Significance | |||||||||||
| Cutting style (CS) | ** | *** | *** | NS | NS | ||||||
| Storage time (ST) | NS | NS | NS | NS | ** | ||||||
| CS × ST | * | ** | ** | NS | NS | ||||||
| Storage Time | Cutting Style A | Phenolic Concentration (mg/kg) B,C,D,E | |||||||||
| 1,2-DFG | 1,2-DS-2-FG | 1-S-2,2-DFG | 1,2,2-TPG | Total Phenolics | |||||||
| 0 h | CS1 | 374.8 ± 13.8 | cd (x) | 235.2 ± 32.5 | bc (x) | 61.8 ± 4.3 | ab (x) | 35.1 ± 0.5 | c (x) | 1668.9 ± 116.9 | bc (x) |
| CS2 | 443.8 ± 39.4 | bc (x) | 259.4 ± 39.8 | abc (x) | 64.7 ± 7.5 | ab (x) | 35.9 ± 1.9 | bc (x) | 2213.5 ± 272.4 | ab (x) | |
| CS3 | 323.2 ± 9.7 | d (y) | 330.5 ± 77.7 | ab (x) | 78.7 ± 16.5 | ab (x) | 46.2 ± 6.9 | abc (x) | 2140.7 ± 376.8 | abc (x) | |
| CS4 | 398.3 ± 5.4 | cd (x) | 232.8 ± 22.7 | bc (x) | 63.5 ± 3.3 | ab (x) | 36.7 ± 3.2 | bc (x) | 1987.9 ± 69.6 | abc (x) | |
| 24 h | CS1 | 600.4 ± 60.9 | a (x) | 367.4 ± 57.4 | ab (x) | 79.8 ± 11.4 | ab (xy) | 45.8 ± 4.2 | abc (x) | 2558.1 ± 272.4 | a (x) |
| CS2 | 520.5 ± 59.9 | ab (x) | 379.1 ± 64.0 | a (x) | 76.7 ± 11.8 | ab (xy) | 47.6 ± 4.6 | ab (x) | 2312.6 ± 247.2 | a (x) | |
| CS3 | 542.7 ± 2.5 | ab (x) | 349.4 ± 33.3 | ab (x) | 83.4 ± 3.2 | a (x) | 49.3 ± 5.9 | a (x) | 2528.6 ± 64.2 | a (x) | |
| CS4 | 322.6 ± 13.8 | d (y) | 177.9 ± 6.90 | c (y) | 53.7 ± 1.8 | b (y) | 35.8 ± 0.4 | bc (x) | 1513.4 ± 64.1 | c (y) | |
| Significance | |||||||||||
| Cutting style (CS) | ** | NS | NS | ** | NS | ||||||
| Storage time (ST) | ** | NS | NS | NS | NS | ||||||
| CS × ST | ** | NS | NS | NS | * | ||||||
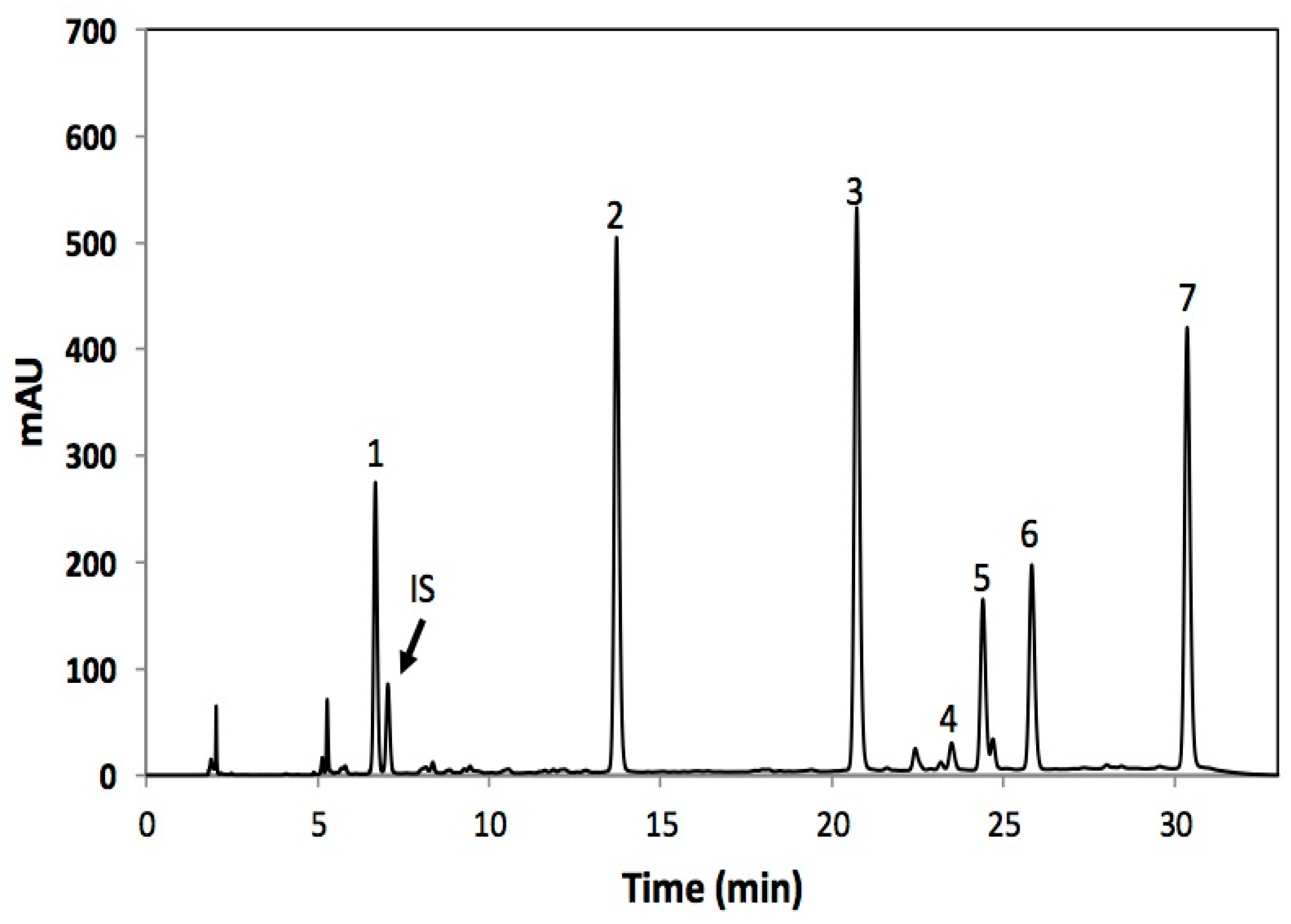
| Peak Number (Retention Time, min) | λ Max (nm) | Tentative Identification | [M − Z]− (m/z) | MS Fragments d |
|---|---|---|---|---|
| 1 (6.6) | 222 | Desulfoglucoraphanin a,b,c | 356 | 193 |
| 2 (13.0) | 222 | 4-hydroxydesulfoglucobrassicin b,c | 383 | 221, 203, 153 |
| 3 (20.8) | 222, 280 | Desulfoglucoerucin b,c | 340 | 177, 160, 129, 113 |
| 4 (23.5) | 222, 280 | Desulfoglucobrassicin b,c | 367 | 204, 187, 155, 129 |
| 5 (24.3) | 222, 265, 280 | Desulfogluconasturtiin b,c | 342 | 179, 162, 130, 104 |
| 6 (25.8) | 222, 290 | 4-methoxydesulfoglucobrassicin b,c | 397 | 234, 204, 154, 139 |
| 7 (30.3) | 222, 290 | Desulfoneoglucobrassicin | 397 | 234, 204, 154, 129 |
| Storage Time | Cutting Style A | Glucosinolate Concentration (μmol/kg DW) B,C,D,E | |||||||
| Glucoraphanin | 4-Hydroxy Glucobrassicin | Glucoerucin | Glucobrassicin | ||||||
| 0 h | CS1 | 1081 ± 87 | bc (w) | 210 ± 21 | d (wx) | 2098 ± 240 | bc (w) | 116 ± 13 | a (x) |
| CS2 | 782 ± 162 | cd (wx) | 173 ± 43 | d (wx) | 1771 ± 434 | cd (wx) | 95 ± 12 | a (x) | |
| CS3 | 617 ± 13 | d (x) | 255 ± 11 | d (w) | 793 ± 53 | d (y) | 121 ± 7 | a (x) | |
| CS4 | 618 ± 43 | d (x) | 140 ± 17 | d (x) | 997 ± 38 | cd (xy) | 92 ± 6 | a (x) | |
| 24 h | CS1 | 4584 ± 198 | a (w) | 2744 ± 6 | a (w) | 11358 ± 263 | a (w) | 165 ± 8 | a (w) |
| CS2 | 854 ± 69 | cd (wx) | 491 ± 98 | c (y) | 1394 ± 578 | cd (y) | 186 ± 81 | a (w) | |
| CS3 | 603 ± 185 | d (y) | 177 ± 60 | d (z) | 1248 ± 559 | cd (y) | 142 ± 24 | a (w) | |
| CS4 | 1185 ± 155 | b (x) | 740 ± 62 | b (x) | 3179 ± 456 | b (x) | 118 ± 8 | a (w) | |
| Significance | |||||||||
| Cutting style (CS) | *** | *** | *** | NS | |||||
| Storage time (ST) | *** | *** | *** | NS | |||||
| CS × ST | *** | *** | *** | NS | |||||
| Storage Time | Cutting Style A | Glucosinolate Concentration (μmol/kg DW) B,C,D,E | |||||||
| Gluconasturtiin | 4-Methoxy Glucobrassicin | Neoglucobrassicin | Total Glucosinolates | ||||||
| 0 h | CS1 | 595 ± 17 | b (w) | 120 ± 7 | c (w) | 365 ± 30 | c (w) | 3505 ± 298 | c (w) |
| CS2 | 491 ± 87 | b (wx) | 81 ± 24 | cd (wx) | 280 ± 36 | c (wx) | 2891 ± 587 | cd (wx) | |
| CS3 | 295 ± 8.0 | b (x) | 71 ± 16 | cd (x) | 222 ± 33 | c (x) | 1757 ± 67 | d (x) | |
| CS4 | 441 ± 111 | b (wx) | 40 ± 2 | d (x) | 194 ± 2 | c (x) | 1906 ± 71 | cd (x) | |
| 24 h | CS1 | 3052 ± 210 | a (w) | 1155 ± 40 | a (w) | 1817 ± 65 | a (w) | 20289 ± 451 | a (w) |
| CS2 | 601 ± 265 | b (x) | 205 ± 13 | b (x) | 233 ± 17 | c (x) | 3032 ± 950 | cd (y) | |
| CS3 | 487 ± 69 | b (x) | 99 ± 40 | cd (y) | 359 ± 161 | c (x) | 2512 ± 848 | cd (y) | |
| CS4 | 3096 ± 383 | a (w) | 90 ± 7.5 | cd (y) | 606 ± 58 | b (x) | 7829 ± 964 | b (x) | |
| Significance | |||||||||
| Cutting style (CS) | ** | *** | *** | *** | |||||
| Storage time (ST) | *** | *** | *** | *** | |||||
| CS × ST | * | *** | *** | *** | |||||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres-Contreras, A.M.; Nair, V.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Stability of Bioactive Compounds in Broccoli as Affected by Cutting Styles and Storage Time. Molecules 2017, 22, 636. https://doi.org/10.3390/molecules22040636
Torres-Contreras AM, Nair V, Cisneros-Zevallos L, Jacobo-Velázquez DA. Stability of Bioactive Compounds in Broccoli as Affected by Cutting Styles and Storage Time. Molecules. 2017; 22(4):636. https://doi.org/10.3390/molecules22040636
Chicago/Turabian StyleTorres-Contreras, Ana Mariel, Vimal Nair, Luis Cisneros-Zevallos, and Daniel A. Jacobo-Velázquez. 2017. "Stability of Bioactive Compounds in Broccoli as Affected by Cutting Styles and Storage Time" Molecules 22, no. 4: 636. https://doi.org/10.3390/molecules22040636
APA StyleTorres-Contreras, A. M., Nair, V., Cisneros-Zevallos, L., & Jacobo-Velázquez, D. A. (2017). Stability of Bioactive Compounds in Broccoli as Affected by Cutting Styles and Storage Time. Molecules, 22(4), 636. https://doi.org/10.3390/molecules22040636