
Sulfated Alginates as Heparin Analogues: A Review of Chemical and Functional Properties


Abstract
:1. Introduction
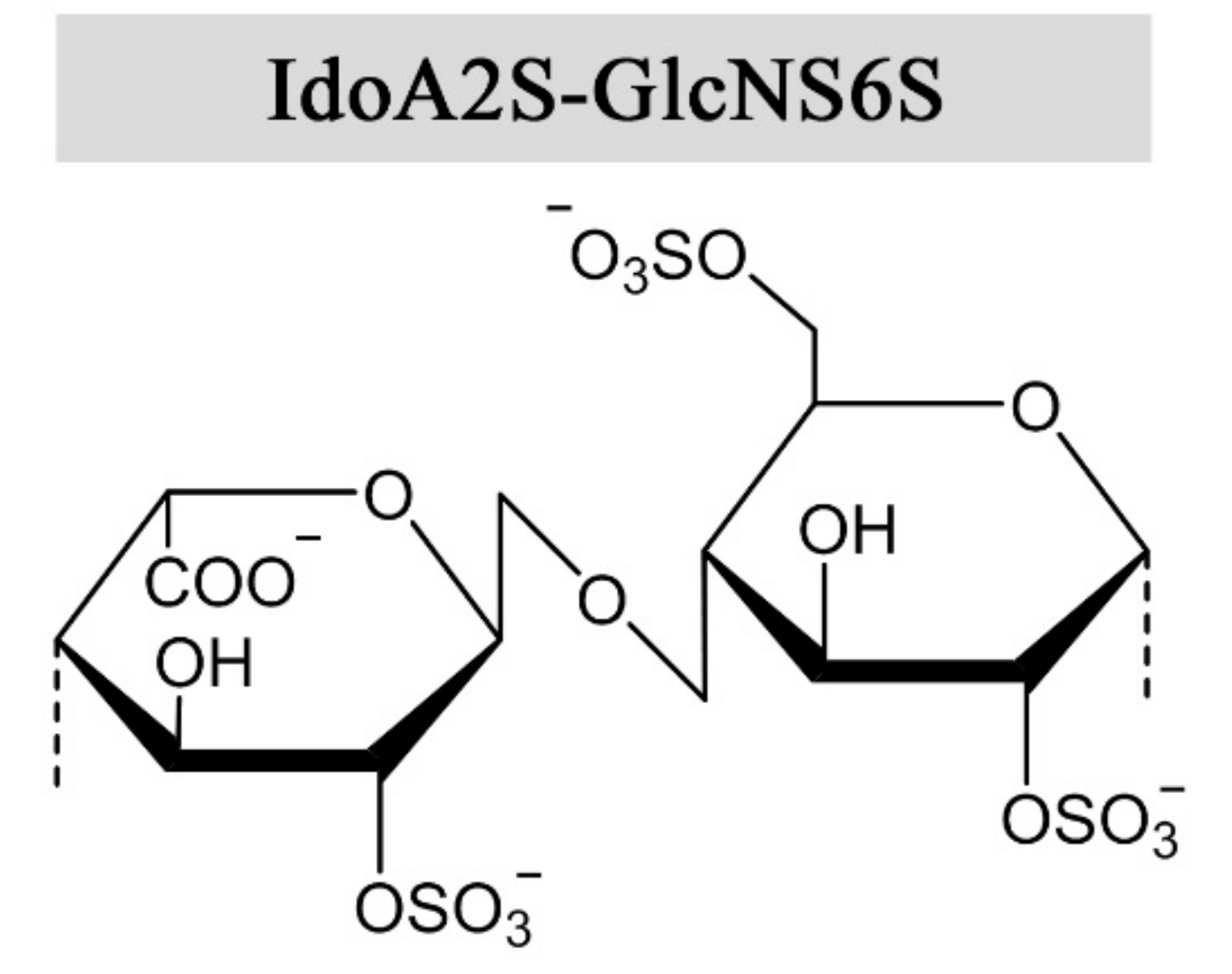
1.1. Heparin Molecular Structure and Physical Properties
1.2. Biological Properties of Heparin and Structure-Function Relationships
2. Sulfated Alginates
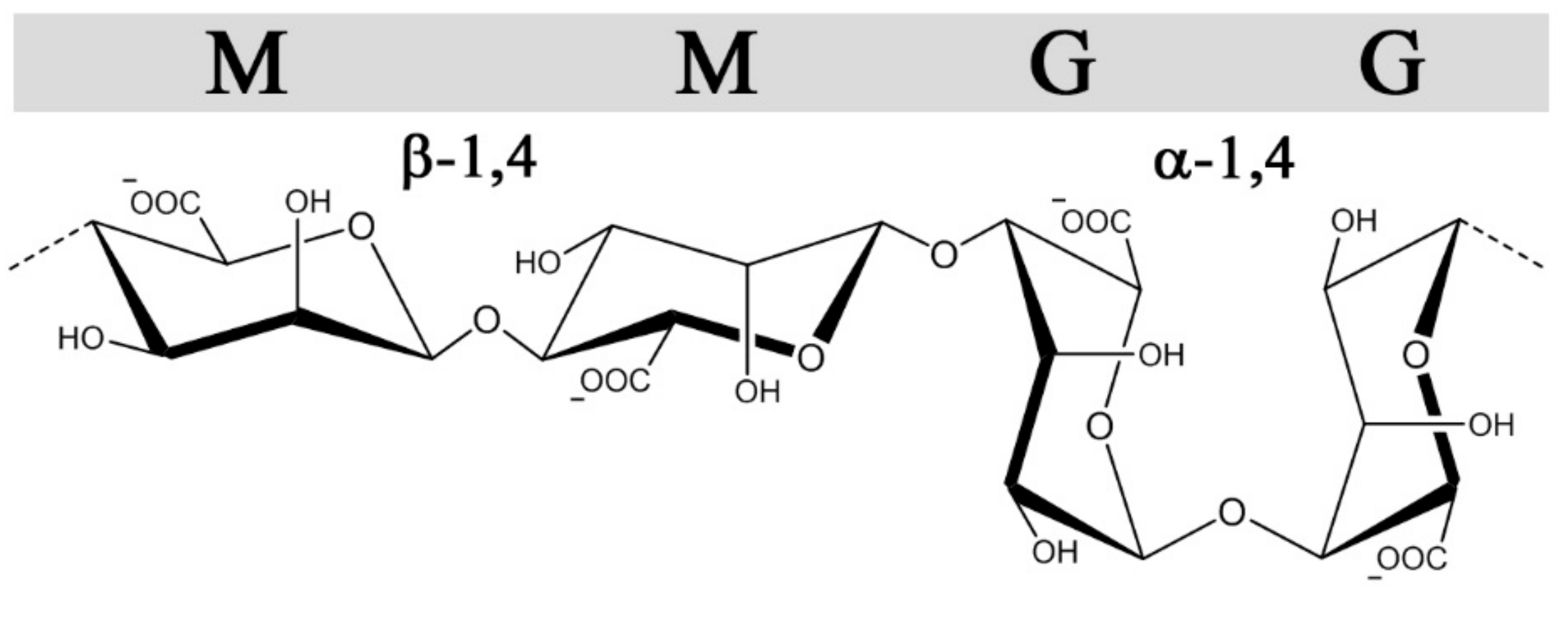
2.1. Properties of Alginate
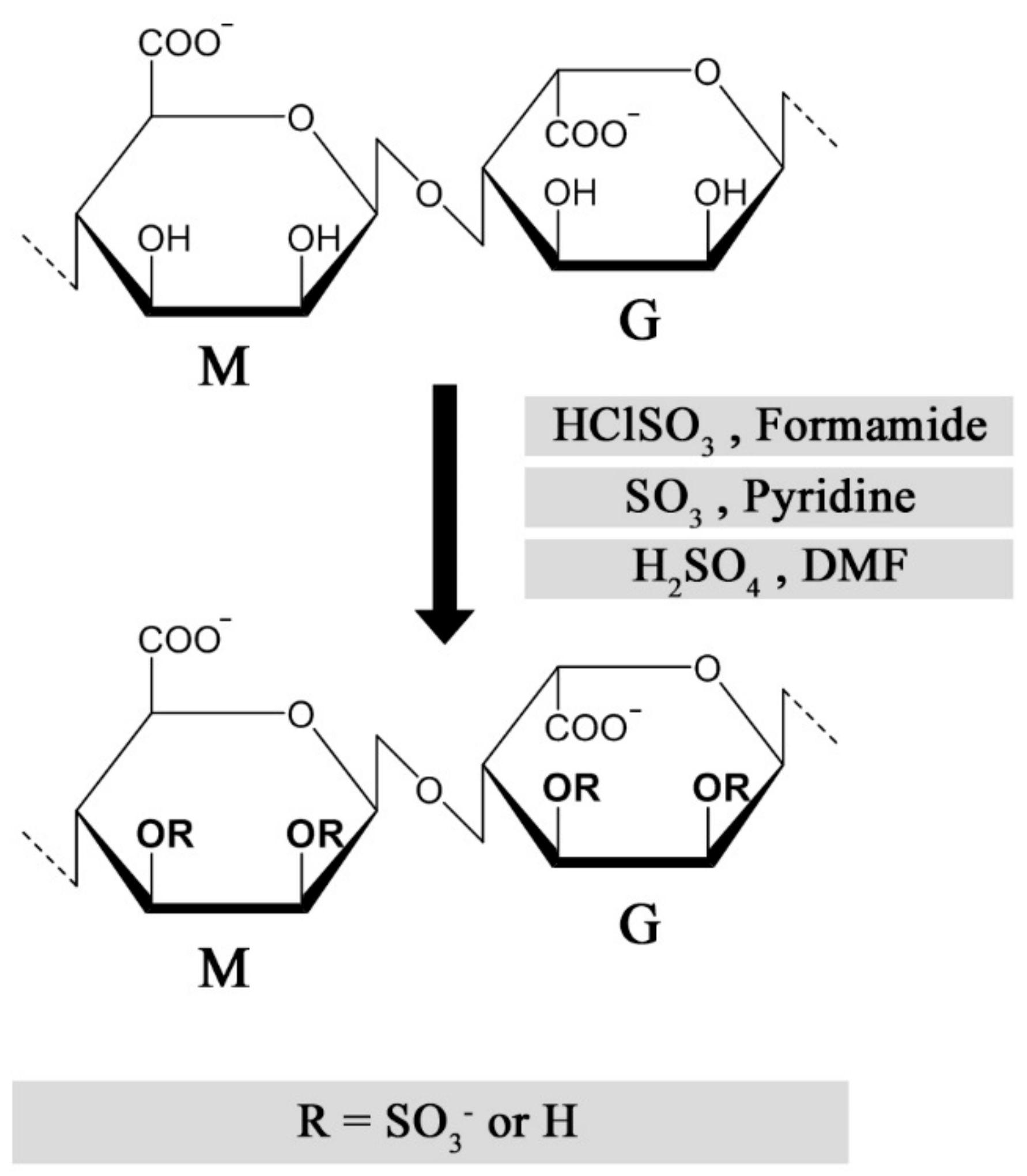
2.2. Synthesis and Characterization of Sulfated Alginates
2.3. Chemical and Physical Properties of Sulfated Alginates
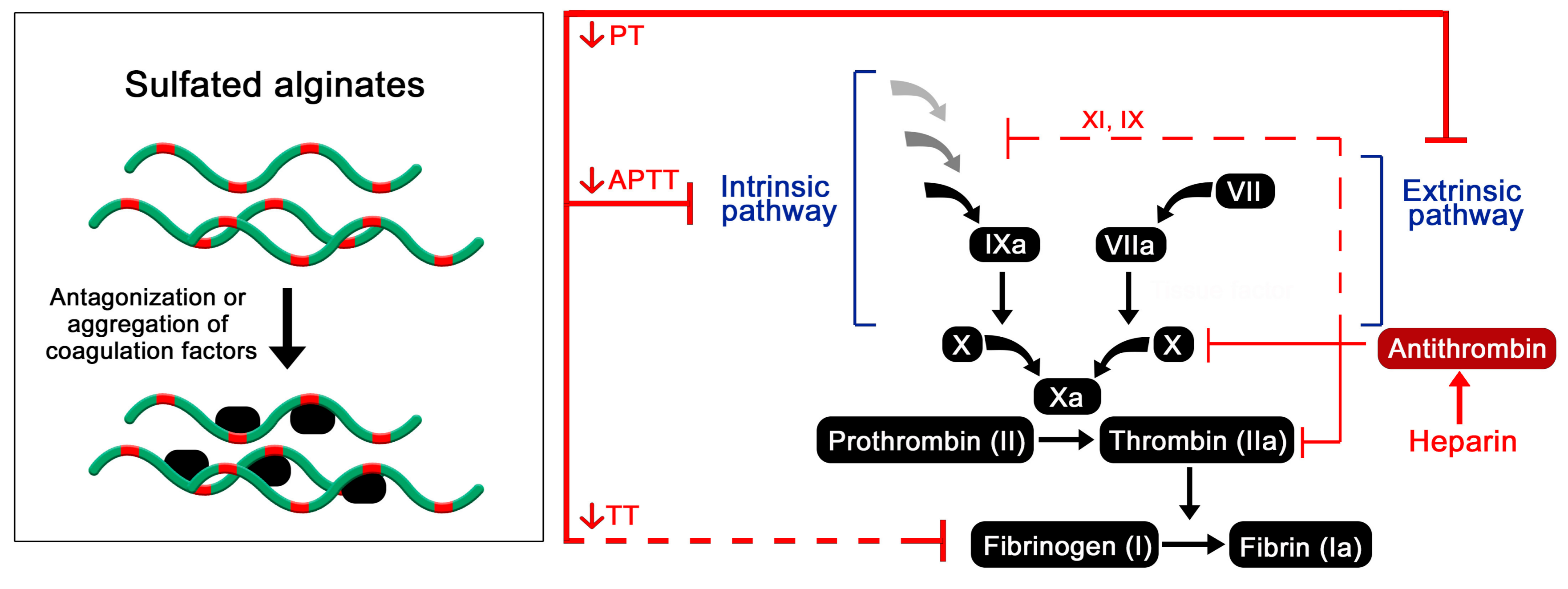
2.4. Effects of Sulfated Alginates on the Coagulation Cascade
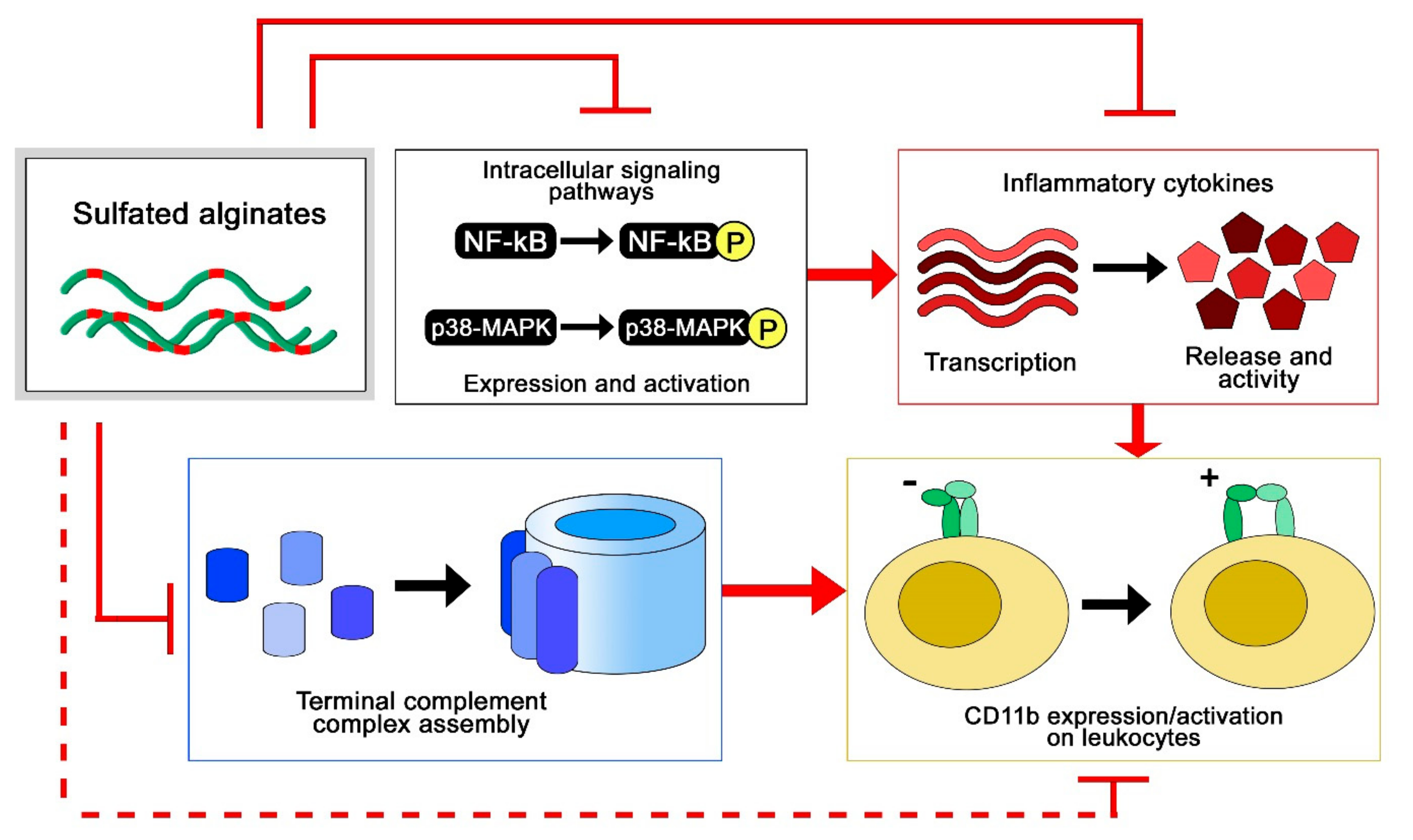
2.5. Immunological Effects of Sulfated Alginates
2.6. Sulfated Alginates in Tissue Engineering and Drug Delivery
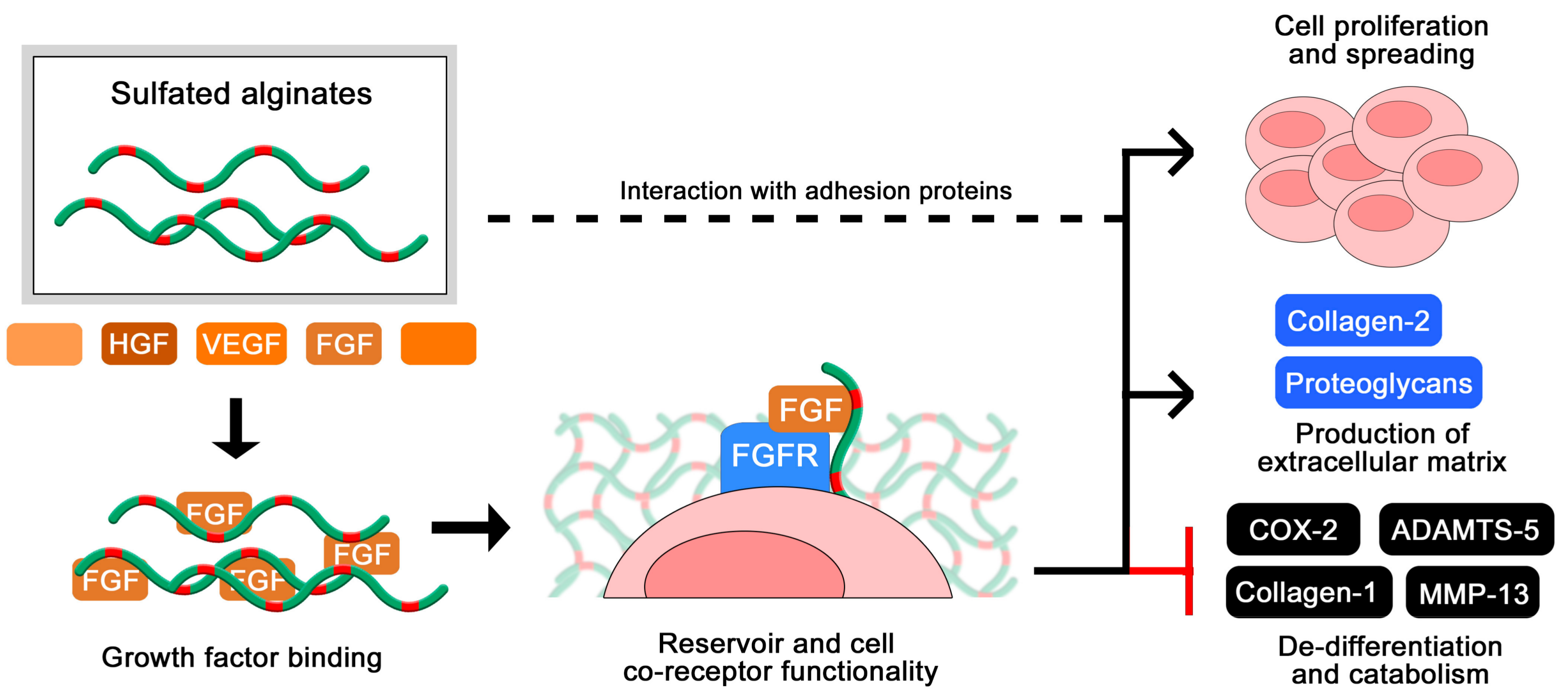
2.7. Structure-Function Relationships in Sulfated Alginates
3. Conclusions and Future Directions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Liu, H.Y.; Zhang, Z.Q.; Linhardt, R.J. Lessons Learned from the Contamination of Heparin. Nat. Prod. Rep. 2009, 26, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, U.; Li, J.P.; Kusche-Gullberg, M.; Salmivirta, M.; Alaranta, S.; Veromaa, T.; Emeis, J.; Roberts, I.; Taylor, C.; Oreste, P.; et al. Generation of “Neoheparin“ from E Coli K5 Capsular Polysaccharide. J. Med. Chem. 2005, 48, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Codee, J.D.C.; Stubba, B.; Schiattarella, M.; Overkleeft, H.S.; van Boeckel, C.A.A.; van Boom, J.H.; van der Marel, G.A. A Modular Strategy toward the Synthesis of Heparin-Like Oligosaccharides Using Monomeric Building Blocks in a Sequential Glycosylation Strategy. J. Am. Chem. Soc. 2005, 127, 3767–3773. [Google Scholar] [CrossRef] [PubMed]
- Jiao, G.L.; Yu, G.L.; Zhang, J.Z.; Ewart, H.S. Chemical Structures and Bioactivities of Sulfated Polysaccharides from Marine Algae. Mar. Drugs 2011, 9, 196–223. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Liu, X.Y.; Ma, L.; Cheng, C.; Shi, W.B.; Nie, C.X.; Zhao, C.S. Heparin-Mimicking Multilayer Coating on Polymeric Membrane Via Lbl Assembly of Cyclodextrin-Based Supramolecules. ACS Appl. Mater. Interfaces 2014, 6, 21603–21614. [Google Scholar] [CrossRef] [PubMed]
- Sasisekharan, R.; Venkataraman, G. Heparin and Heparan Sulfate: Biosynthesis, Structure and Function. Curr. Opin. Chem. Biol. 2000, 4, 626–631. [Google Scholar] [CrossRef]
- Raman, R.; Venkataraman, G.; Ernst, S.; Sasisekharan, V.; Sasisekharan, R. Structural Specificity of Heparin Binding in the Fibroblast Growth Factor Family of Proteins. Proc. Natl. Acad. Sci. USA 2003, 100, 2357–2362. [Google Scholar] [CrossRef] [PubMed]
- Habuchi, H.; Tanaka, M.; Habuchi, O.; Yoshida, K.; Suzuki, H.; Ban, K.; Kimata, K. The Occurrence of Three Isoforms of Heparan Sulfate 6-O-Sulfotransferase Having Different Specificities for Hexuronic Acid Adjacent to the Targeted N-Sulfoglucosamine. J. Biol. Chem. 2000, 275, 2859–2868. [Google Scholar] [CrossRef] [PubMed]
- Aikawa, J.; Grobe, K.; Tsujimoto, M.; Esko, J.D. Multiple Isozymes of Heparan Sulfate/Heparin Glcnac N-Deacetylase/Glcn N-Sulfotransferase-Structure and Activity of the Fourth Member, Ndst4. J. Biol. Chem. 2001, 276, 5876–5882. [Google Scholar] [CrossRef] [PubMed]
- Sudo, M.; Sato, K.; Chaidedgumjorn, A.; Toyoda, H.; Toida, T.; Imanari, T. H-1 Nuclear Magnetic Resonance Spectroscopic Analysis for Determination of Glucuronic and Iduronic Acids in Dermatan Sulfate, Heparin, and Heparan Sulfate. Anal. Biochem. 2001, 297, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Guerrini, M.; Bisio, A.; Torri, G. Combined Quantitative H-1 and C-13 Nuclear Magnetic Resonance Spectroscopy for Characterization of Heparin Preparations. Semin. Thromb. Hemost. 2001, 27, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Maccarana, M.; Sakura, Y.; Tawada, A.; Yoshida, K.; Lindahl, U. Domain Structure of Heparan Sulfates from Bovine Organs. J. Biol. Chem. 1996, 271, 17804–17810. [Google Scholar] [CrossRef] [PubMed]
- Hagner-McWhirter, A.; Lindahl, U.; Li, J.P. Biosynthesis of Heparin/Heparan Sulphate: Mechanism of Epimerization of Glucuronyl C-5. Biochem. J. 2000, 347, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Raedts, J.M.; Lundgren, M.; Kengen, S.W.M.; Li, J.P.; van der Oost, J. A Novel Bacterial Enzyme with D-Glucuronyl C5-Epimerase Activity. J. Biol. Chem. 2013, 288, 24332–24339. [Google Scholar] [CrossRef] [PubMed]
- Lidholt, K.; Fjelstad, M.; Jann, K.; Lindahl, U. Biosynthesis of Heparin 25. Substrate Specificities of Glycosyltransferases Involved in Formation of Heparin Precursor and Escherichia-Coli K5 Capsular Polysaccharides. Carbohydr. Res. 1994, 255, 87–101. [Google Scholar] [CrossRef]
- Lindahl, U.; Backstrom, G.; Hook, M.; Thunberg, L.; Fransson, L.A.; Linker, A. Structure of the Antithrombin-Binding Site in Heparin. Proc. Natl. Acad. Sci. USA 1979, 76, 3198–3202. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.U.; Miller, G.J.; Cliff, M.J.; Jaysonc, G.C.; Gardiner, J.M. Making the Longest Sugars: A Chemical Synthesis of Heparin-Related [4](N) Oligosaccharides from 16-Mer to 40-Mer. Chem. Sci. 2015, 6, 6158–6164. [Google Scholar] [CrossRef]
- Rosenberg, R.D. Actions and Interactions of Antithrombin and Heparin. N. Engl. J. Med. 1975, 292, 146–151. [Google Scholar] [PubMed]
- Sahu, A.; Pangburn, M.K. Identification of Multiple Sites of Interaction between Heparin and the Complement-System. Mol. Immunol. 1993, 30, 679–684. [Google Scholar] [CrossRef]
- Amara, U.; Flierl, M.A.; Rittirsch, D.; Klos, A.; Chen, H.; Acker, B.; Bruckner, U.B.; Nilsson, B.; Gebhard, F.; Lambris, J.D.; et al. Molecular Intercommunication between the Complement and Coagulation Systems. J. Immunol. 2010, 185, 5628–5636. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, N.S.; Mancera, R.L. The Structure of Glycosaminoglycans and Their Interactions with Proteins. Chem. Biol. Drug Des. 2008, 72, 455–482. [Google Scholar] [CrossRef] [PubMed]
- Nelson, R.M.; Cecconi, O.; Roberts, W.G.; Aruffo, A.; Linhardt, R.J.; Bevilacqua, M.P. Heparin Oligosaccharides Bind L-Selectin and P-Selectin and Inhibit Acute-Inflammation. Blood 1993, 82, 3253–3258. [Google Scholar] [PubMed]
- Joyce, J.G.; Tung, J.S.; Przysiecki, C.T.; Cook, J.C.; Lehman, E.D.; Sands, J.A.; Jansen, K.U.; Keller, P.M. The L1 Major Capsid Protein of Human Papillomavirus Type 11 Recombinant Virus-Like Particles Interacts with Heparin and Cell-Surface Glycosaminoglycans on Human Keratinocytes. J. Biol. Chem. 1999, 274, 5810–5822. [Google Scholar] [CrossRef] [PubMed]
- Ascencio, F.; Fransson, L.A.; Wadstrom, T. Affinity of the Gastric Pathogen Helicobacter-Pylori for the N-Sulfated Glycosaminoglycan Heparan-Sulfate. J. Med. Microbiol. 1993, 38, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Spillmann, D.; Lindahl, U. Glycosaminoglycan Protein Interactions-a Question of Specificity. Curr. Opin. Struct. Biol. 1994, 4, 677–682. [Google Scholar] [CrossRef]
- Lindahl, U. A Personal Voyage through the Proteoglycan Field. Matrix Biol. 2014, 35, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.P.; Zhong, Y.Q.; Chen, Z.G.; Chen, C.Y.; Shi, Z.; Zulueta, M.M.; Ku, C.C.; Lee, P.Y.; Wang, C.C.; Hung, S.C. Divergent Synthesis of 48 Heparan Sulfate-Based Disaccharides and Probing the Specific Sugar-Fibroblast Growth Factor-1 Interaction. J. Am. Chem. Soc. 2012, 134, 20722–20727. [Google Scholar] [CrossRef] [PubMed]
- Foxall, C.; Holme, K.R.; Liang, W.H.; Wei, Z. An Enzyme-Linked-Immunosorbent-Assay Using Biotinylated Heparan-Sulfate to Evaluate the Interactions of Heparin-Like Molecules and Basic Fibroblast Growth-Factor. Anal. Biochem. 1995, 231, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.C.; Brown, J.R.; Varki, A.; Esko, J.D. Heparin’s Anti-Inflammatory Effects Require Glucosamine 6-O-Sulfation and Are Mediated by Blockade of L- and P-Selectins. J. Clin. Investig. 2002, 110, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Koenig, A.; Norgard-Sumnicht, K.; Linhardt, R.; Varki, A. Differential Interactions of Heparin and Heparan Sulfate Glycosaminoglycans with the Selectins-Implications for the Use of Unfractionated and Low Molecular Weight Heparins as Therapeutic Agents. J. Clin Investig. 1998, 101, 877–889. [Google Scholar] [CrossRef] [PubMed]
- Skjåk-Bræk, G.; Donari, I.; Paoletti, S. Alginate Hydrogels: Properties and Applications. In Polysaccharide Hydrogels; Matricardi, P., Alhaique, F., Coviello, T., Eds.; Pan Stanford: Boca Raton, FL, USA, 2015. [Google Scholar]
- Sarrazin, S.; Lamanna, W.C.; Esko, J.D. Heparan Sulfate Proteoglycans. Cold Spring Harbor Perspect. Biol. 2011, 3, 1–33. [Google Scholar] [CrossRef] [PubMed]
- Stokke, B.T.; Smidsrød, O.; Bruheim, P.; Skjåk-Bræk, G. Distribution of Uronate Residues in Alginate Chains in Relation to Alginate Gelling Properties. Macromolecules 1991, 24, 4637–4645. [Google Scholar] [CrossRef]
- Mørch, Y.A.; Donati, I.; Strand, B.L.; Skjåk-Bræk, G. Molecular Engineering as an Approach to Design New Functional Properties of Alginate. Biomacromolecules 2007, 8, 2809–2814. [Google Scholar] [CrossRef] [PubMed]
- Høidal, H.K.; Ertesvåg, H.; Skjåk-Bræk, G.; Stokke, B.T.; Valla, S. The Recombinant Azotobacter Vinelandii Mannuronan C-5-Epimerase Alge4 Epimerizes Alginate by a Nonrandom Attack Mechanism. J. Biol. Chem. 1999, 274, 12316–12322. [Google Scholar] [CrossRef] [PubMed]
- Pawar, S.N.; Edgar, K.J. Alginate Derivatization: A Review of Chemistry, Properties and Applications. Biomaterials 2012, 33, 3279–3305. [Google Scholar] [CrossRef] [PubMed]
- Ertesvåg, H. Alginate-Modifying Enzymes: Biological Roles and Biotechnological Uses. Front. Microbiol. 2015, 6, 1–10. [Google Scholar]
- Jacobs-Tulleneers-Thevissen, D.; Chintinne, M.; Ling, Z.; Gillard, P.; Schoonjans, L.; Delvaux, G.; Strand, B.L.; Gorus, F.; Keymeulen, B.; Pipeleers, D.; et al. Sustained Function of Alginate-Encapsulated Human Islet Cell Implants in the Peritoneal Cavity of Mice Leading to a Pilot Study in a Type 1 Diabetic Patient. Diabetologia 2013, 56, 1605–1614. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, Q.; Wang, A. In Situ Generation of Sodium Alginate/Hydroxyapatite Nanocomposite Beads as Drug-Controlled Release Matrices. Acta Biomate. 2010, 6, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Sandvig, I.; Karstensen, K.; Rokstad, A.M.; Aachmann, F.L.; Formo, K.; Sandvig, A.; Skjåk-Bræk, G.; Strand, B.L. Rgd-Peptide Modified Alginate by a Chemoenzymatic Strategy for Tissue Engineering Applications. J. Biomed. Mater. Res. Part A 2015, 103, 896–906. [Google Scholar] [CrossRef] [PubMed]
- Markstedt, K.; Mantas, A.; Tournier, I.; Martinez, H.; Hägg, D.; Gatenholm, P. 3D Bioprinting Human Chondrocytes with Nanocellulose-Alginate Bioink for Cartilage Tissue Engineering Applications. Biomacromolecules 2015, 16, 1489–1496. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.H.; Du, Y.M.; Yang, J.H. Preparation and in Vitro Anticoagulant Activities of Alginate Sulfate and Its Quaterized Derivatives. Carbohydr. Polym. 2003, 52, 19–24. [Google Scholar]
- Arlov, Ø.; Aachmann, F.L.; Feyzi, E.; Sundan, A.; Skjåk-Bræk, G. The Impact of Chain Length and Flexibility in the Interaction between Sulfated Alginates and Hgf and Fgf-2. Biomacromolecules 2015, 16, 3417–3424. [Google Scholar] [CrossRef] [PubMed]
- Arlov, Ø.; Aachmann, F.L.; Sundan, A.; Espevik, T.; Skjåk-Bræk, G. Heparin-Like Properties of Sulfated Alginates with Defined Sequences and Sulfation Degrees. Biomacromolecules 2014, 15, 2744–2750. [Google Scholar] [CrossRef] [PubMed]
- Miyaji, H.; Misaki, A. Distribution of Sulfate Groups in the Partially Sulfated Dextrans. J. Biochem. 1973, 74, 1131–1139. [Google Scholar] [CrossRef] [PubMed]
- Mhanna, R.; Kashyap, A.; Palazzolo, G.; Vallmajo-Martin, Q.; Becher, J.; Möller, S.; Schnabelrauch, M.; Zenobi-Wong, M. Chondrocyte Culture in Three Dimensional Alginate Sulfate Hydrogels Promotes Proliferation While Maintaining Expression of Chondrogenic Markers. Tissue Engl. Part A 2014, 20, 1454–1464. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Cheng, C.; Nie, C.X.; He, C.; Deng, J.; Wang, L.R.; Xia, Y.; Zhao, C.S. Anticoagulant Sodium Alginate Sulfates and Their Mussel-Inspired Heparin-Mimetic Coatings. J. Mater. Chem. B 2016, 4, 203–215. [Google Scholar] [CrossRef]
- Freeman, I.; Kedem, A.; Cohen, S. The Effect of Sulfation of Alginate Hydrogels on the Specific Binding and Controlled Release of Heparin-Binding Proteins. Biomaterials 2008, 29, 3260–3268. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.H.; Jiang, L.; Xu, Y.M.; Zhou, Y.; Shen, Y.A.; Xie, W.G.; Long, Z.H.; Zhou, J.P. Synthesis and Anticoagulant Activity of Sodium Alginate Sulfates. Carbohydr. Polym. 2011, 83, 1797–1803. [Google Scholar] [CrossRef]
- Zhao, X.; Yu, G.L.; Guan, H.S.; Yue, N.; Zhang, Z.Q.; Li, H.H. Preparation of Low-Molecular-Weight Polyguluronate Sulfate and Its Anticoagulant and Anti-Inflammatory Activities. Carbohydr. Polym. 2007, 69, 272–279. [Google Scholar] [CrossRef]
- Lindahl, U.; Kusche-Gullberg, M.; Kjellen, L. Regulated Diversity of Heparan Sulfate. J. Biol. Chem. 1998, 273, 24979–24982. [Google Scholar] [CrossRef] [PubMed]
- Dodgson, K.S.; Price, R.G. A Note on the Determination of the Ester Sulphate Content of Sulphated Polysaccharides. Biochem. J. 1962, 84, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.H.; Du, Y.M.; Huang, R.H.; Wan, Y.Y.; Wen, Y. The Structure-Anticoagulant Activity Relationships of Sulfated Lacquer Polysaccharide—Effect of Carboxyl Group and Position of Sulfation. Int. J. Biol. Macromol. 2005, 36, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Heymann, B.; Grubmuller, H. Chair-Boat′ Transitions and Side Groups Affect the Stiffness of Polysaccharides. Chem. Phys. Lett. 1999, 305, 202–208. [Google Scholar] [CrossRef]
- Smidsrød, O.; Glover, R.M.; Whittington, S.G. The Relative Extension of Alginates Having Different Chemical Composition. Carbohydr. Res. 1973, 27, 107–118. [Google Scholar] [CrossRef]
- Öztürk, E.; Arlov, Ø.; Aksel, S.; Li, L.; Ornitz, D.M.; Skjåk-Bræk, G.; Zenobi-Wong, M. Sulfated Hydrogel Matrices Direct Mitogenicity and Maintenance of Chondrocyte Phenotype through Activation of Fgf Signaling. Adv. Funct. Mater. 2016, 26, 3649–3662. [Google Scholar] [CrossRef]
- Grant, G.T.; Morris, E.R.; Rees, D.A.; Smith, P.J.C.; Thom, D. Biological Interactions between Polysaccharides and Divalent Cations-Egg-Box Model. Febs. Lett. 1973, 32, 195–198. [Google Scholar] [CrossRef]
- Arlov, Ø.; Steinwachs, M.; Skjåk-Bræk, G.; Zenobi-Wong, M. Biomimetic Sulphated Alginate Hydrogels Suppress Il-1beta-Induced Inflammatory Responses in Human Chondrocytes. Eur. Cell. Mater. 2017, 33, 76–89. [Google Scholar] [CrossRef] [PubMed]
- Mørch, Ý.A.; Donati, I.; Strand, B.L.; Skjåk-Bræk, G. Effect of Ca2+, Ba2+, and Sr2+ on Alginate Microbeads. Biomacromolecules 2006, 7, 1471–1480. [Google Scholar] [CrossRef] [PubMed]
- Draget, K.I.; Østgaard, K.; Smidsrød, O. Homogeneous Alginate Gels-a Technical Approach. Carbohydr. Polym. 1990, 14, 159–178. [Google Scholar] [CrossRef]
- Li, Q.C.; Zeng, Y.Y.; Wang, L.L.; Guan, H.S.; Li, C.X.; Zhang, L.J. The Heparin-Like Activities of Negatively Charged Derivatives of Low-Molecular-Weight Polymannuronate and Polyguluronate. Carbohydr. Polym. 2017, 155, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Mousa, S.A.; Mohamed, S. Inhibition of Endothelial Cell Tube Formation by the Low Molecular Weight Heparin, Tinzaparin, Is Mediated by Tissue Factor Pathway Inhibitor. Thromb. Haemost. 2004, 92, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Cheng, C.; He, C.; Nie, C.X.; Deng, J.; Sun, S.D.; Zhao, C.S. Substrate-Independent Robust and Heparin-Mimetic Hydrogel Thin Film Coating Via Combined Lbl Self-Assembly and Mussel-Inspired Post-Cross-Linking. ACS Appl. Mater. Interfaces 2015, 7, 26050–26062. [Google Scholar] [CrossRef] [PubMed]
- Niimi, Y.; Ichinose, F.; Ishiguro, Y.; Terui, K.; Uezono, S.; Morita, S.; Yamane, S. The Effects of Heparin Coating of Oxygenator Fibers on Platelet Adhesion and Protein Adsorption. Anesth. Analg. 1999, 89, 573–579. [Google Scholar] [PubMed]
- Arlov, Ø.; Skjåk-Bræk, G.; Rokstad, A.M. Sulfated Alginate Microspheres Associate with Factor H and Dampen the Inflammatory Cytokine Response. Acta Biomater. 2016, 42, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Weiler, J.M.; Edens, R.E.; Linhardt, R.J.; Kapelanski, D.P. Heparin and Modified Heparin Inhibit Complement Activation Invivo. J. Immunol. 1992, 148, 3210–3215. [Google Scholar] [PubMed]
- Oikonomopoulou, K.; Ricklin, D.; Ward, P.A.; Lambris, J.D. Interactions between Coagulation and Complement-Their Role in Inflammation. Semin. Immunopathol. 2012, 34, 151–165. [Google Scholar] [CrossRef] [PubMed]
- Spivak-Kroizman, T.; Lemmon, M.A.; Dikic, I.; Ladbury, J.E.; Pinchasi, D.; Huang, J.; Jaye, M.; Crumley, G.; Schlessinger, J.; Lax, I. Heparin-Induced Oligomerization of Fgf Molecules Is Responsible for Fgf Receptor Dimerization, Activation, and Cell-Proliferation. Cell 1994, 79, 1015–1024. [Google Scholar] [CrossRef]
- Izaki, S.; Goldstein, S.M.; Fukuyama, K.; Epstein, W.L. Fibrin Deposition and Clearance in Chronic Granulomatous Inflammation: Correlation with T-Cell Function and Proteinase Inhibitor Activity in Tissue. J. Investig. Dermatol. 1979, 73, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Lukacs, N.W.; Chensue, S.W.; Strieter, R.M.; Warmington, K.; Kunkel, S.L. Inflammatory Granuloma-Formation Is Mediated by Tnf-Alpha-Inducible Intercellular-Adhesion Molecule-1. J. Immunol. 1994, 152, 5883–5889. [Google Scholar] [PubMed]
- Ruvinov, E.; Leor, J.; Cohen, S. The Effects of Controlled Hgf Delivery from an Affinity-Binding Alginate Biomaterial on Angiogenesis and Blood Perfusion in a Hindlimb Ischemia Model. Biomaterials 2010, 31, 4573–4582. [Google Scholar] [CrossRef] [PubMed]
- Freeman, I.; Cohen, S. The Influence of the Sequential Delivery of Angiogenic Factors from Affinity-Binding Alginate Scaffolds on Vascularization. Biomaterials 2009, 30, 2122–2131. [Google Scholar] [CrossRef] [PubMed]
- Re’em, T.; Witte, F.; Willbold, E.; Ruvinov, E.; Cohen, S. Simultaneous Regeneration of Articular Cartilage and Subchondral Bone Induced by Spatially Presented Tgf-Beta and Bmp-4 in a Bilayer Affinity Binding System. Acta Biomater. 2012, 8, 3283–3293. [Google Scholar] [CrossRef] [PubMed]
- Ruvinov, E.; Freeman, I.; Fredo, R.; Cohen, S. Spontaneous Coassembly of Biologically Active Nanoparticles Via Affinity Binding of Heparin-Binding Proteins to Alginate-Sulfate. Nano Lett. 2016, 16, 883–888. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.; Ece, Ö.; Øystein, A.; Paul, G.; Zenobi-Wong, M. Alginate Sulfate–Nanocellulose Bioinks for Cartilage Bioprinting Applications. Annals Biomed. Eng. 2016, 45, 210–223. [Google Scholar] [CrossRef] [PubMed]
- Makris, E.A.; Gomoll, A.H.; Malizos, K.N.; Hu, J.C.; Athanasiou, K.A. Repair and Tissue Engineering Techniques for Articular Cartilage. Nat. Rev. Rheumatol. 2015, 11, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Duflo, S.; Thibeault, S.L.; Li, W.H.; Shu, X.Z.; Prestwich, G.D. Vocal Fold Tissue Repair in Vivo Using a Synthetic Extracellular Matrix. Tissue Eng. 2006, 12, 2171–2180. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, M.; Dentini, M.; Draget, K.I.; Skjåk-Bræk, G. Enzymatic Modification of Alginates with the Mannuronan C-5 Epimerase Alge4 Enhances Their Solubility at Low Ph. Carbohydr. Poly. 2006, 63, 257–262. [Google Scholar] [CrossRef]
- Liu, H.Y.; Geng, M.Y.; Xin, X.L.; Li, F.C.; Zhang, Z.Q.; Li, J.; Ding, J. Multiple and Multivalent Interactions of Novel Anti-Aids Drug Candidates, Sulfated Polymannuronate (Spmg)-Derived Oligosaccharides, with Gp120 and Their Anti-Hiv Activities. Glycobiology 2005, 15, 501–510. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arlov, Ø.; Skjåk-Bræk, G. Sulfated Alginates as Heparin Analogues: A Review of Chemical and Functional Properties. Molecules 2017, 22, 778. https://doi.org/10.3390/molecules22050778
Arlov Ø, Skjåk-Bræk G. Sulfated Alginates as Heparin Analogues: A Review of Chemical and Functional Properties. Molecules. 2017; 22(5):778. https://doi.org/10.3390/molecules22050778
Chicago/Turabian StyleArlov, Øystein, and Gudmund Skjåk-Bræk. 2017. "Sulfated Alginates as Heparin Analogues: A Review of Chemical and Functional Properties" Molecules 22, no. 5: 778. https://doi.org/10.3390/molecules22050778
APA StyleArlov, Ø., & Skjåk-Bræk, G. (2017). Sulfated Alginates as Heparin Analogues: A Review of Chemical and Functional Properties. Molecules, 22(5), 778. https://doi.org/10.3390/molecules22050778