
Exploring Adenosine Receptor Ligands: Potential Role in the Treatment of Cardiovascular Diseases


Abstract
:1. Introduction
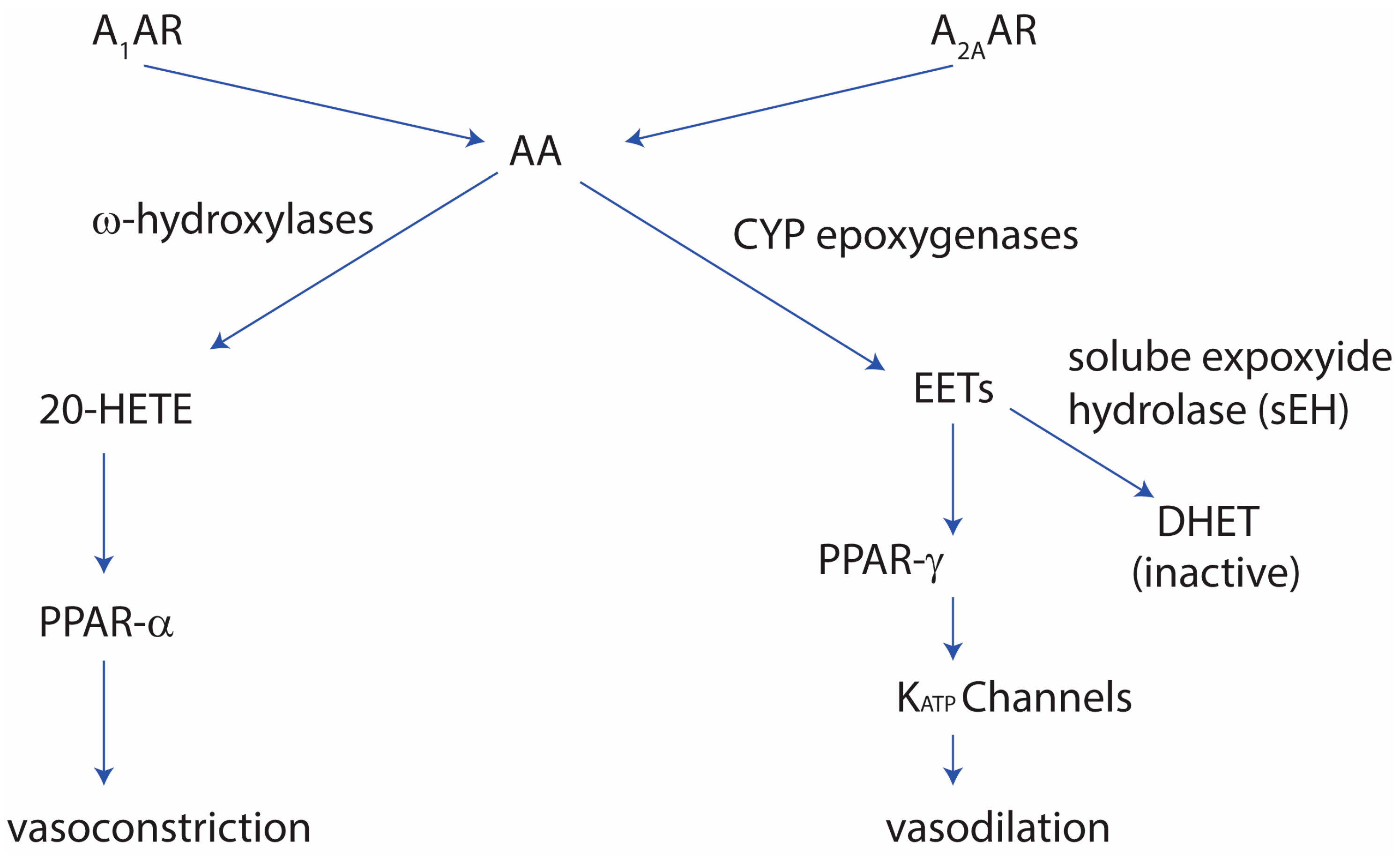
2. Role of Adenosine in Cardiovascular Function
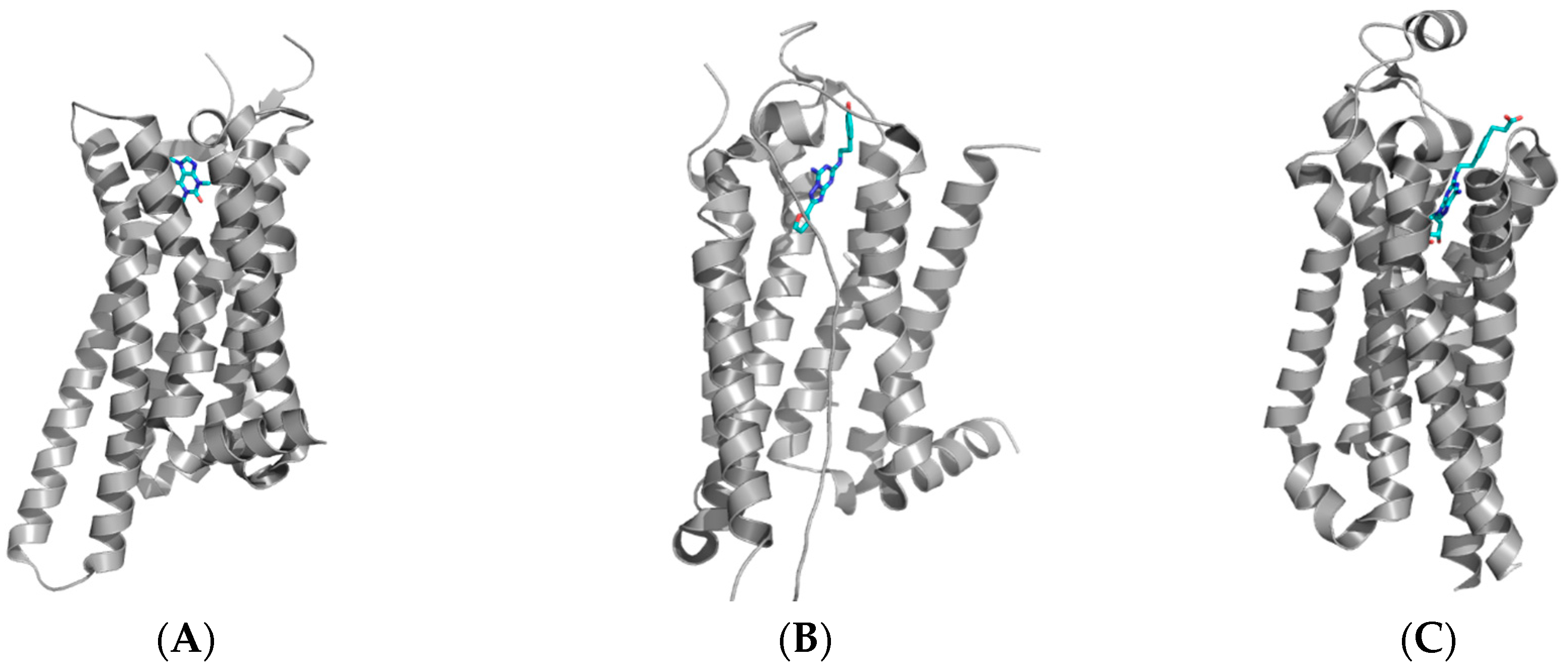
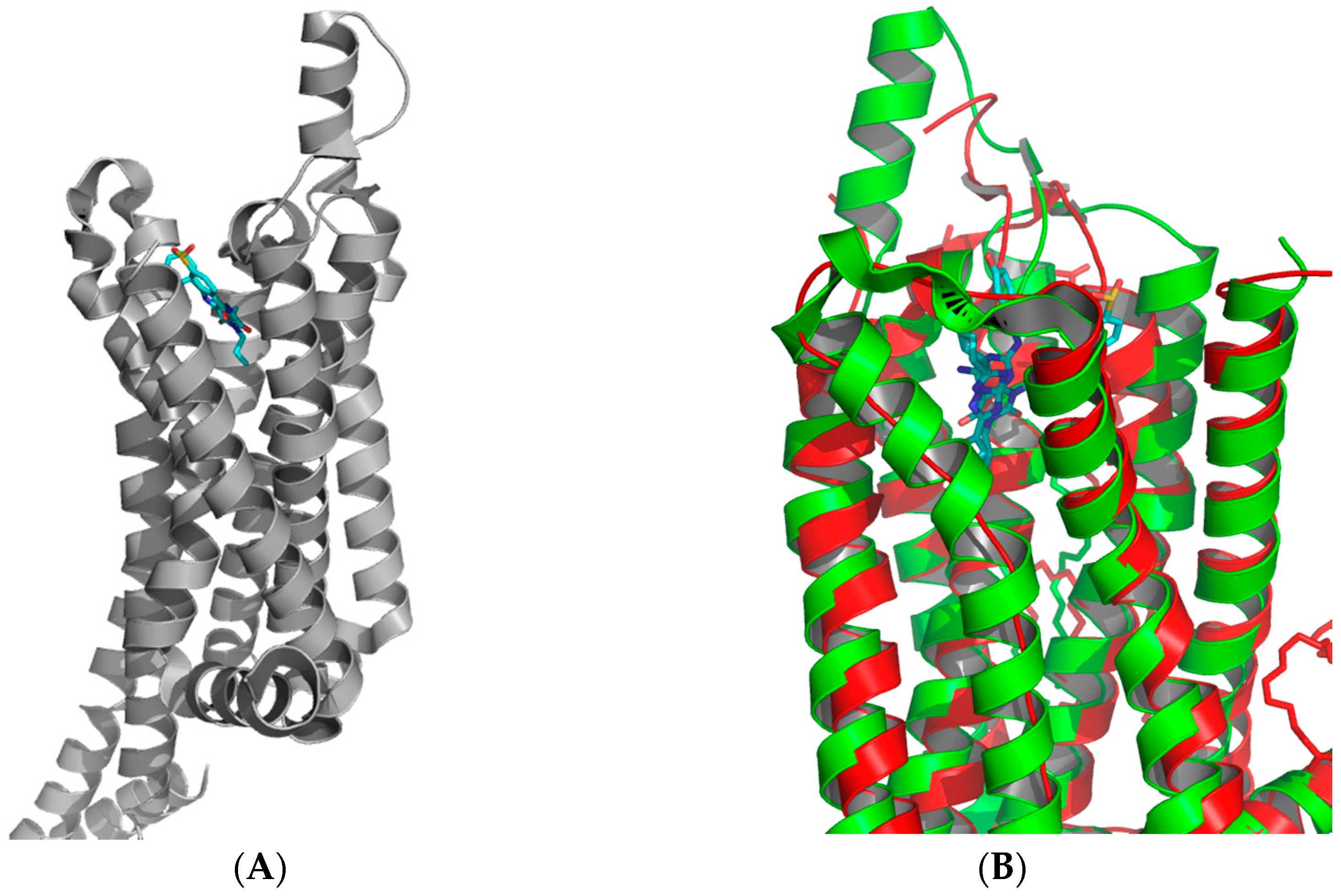
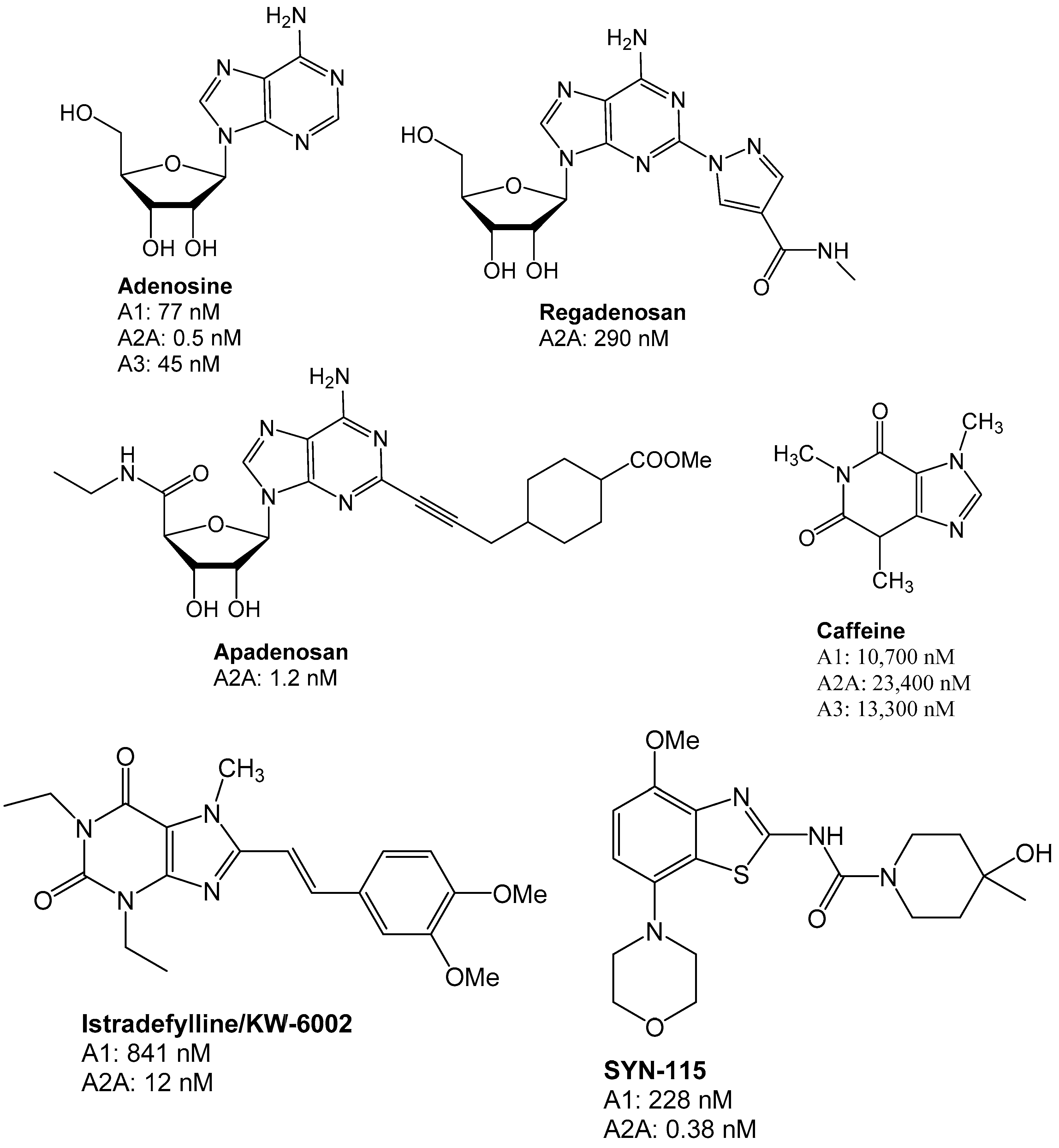
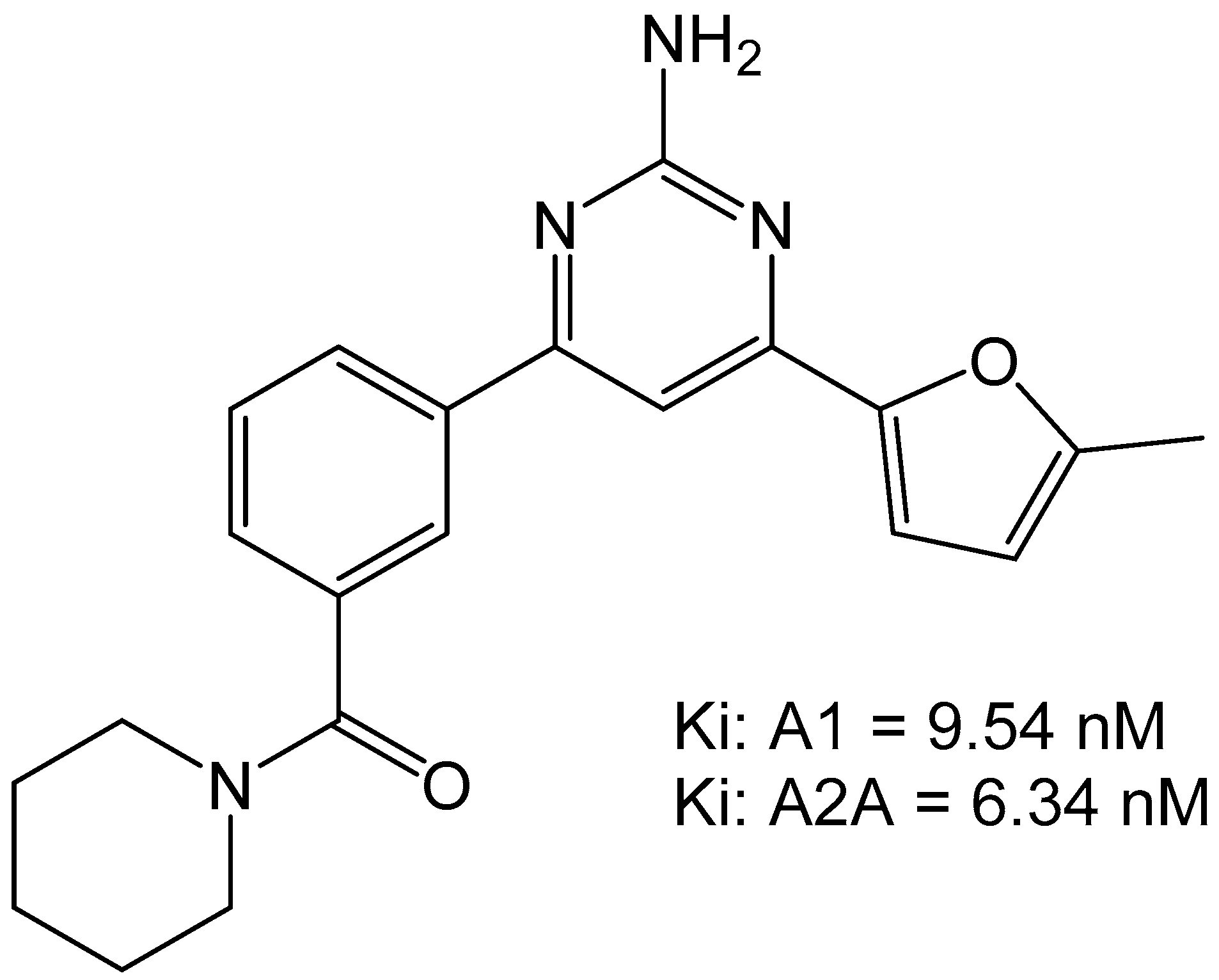
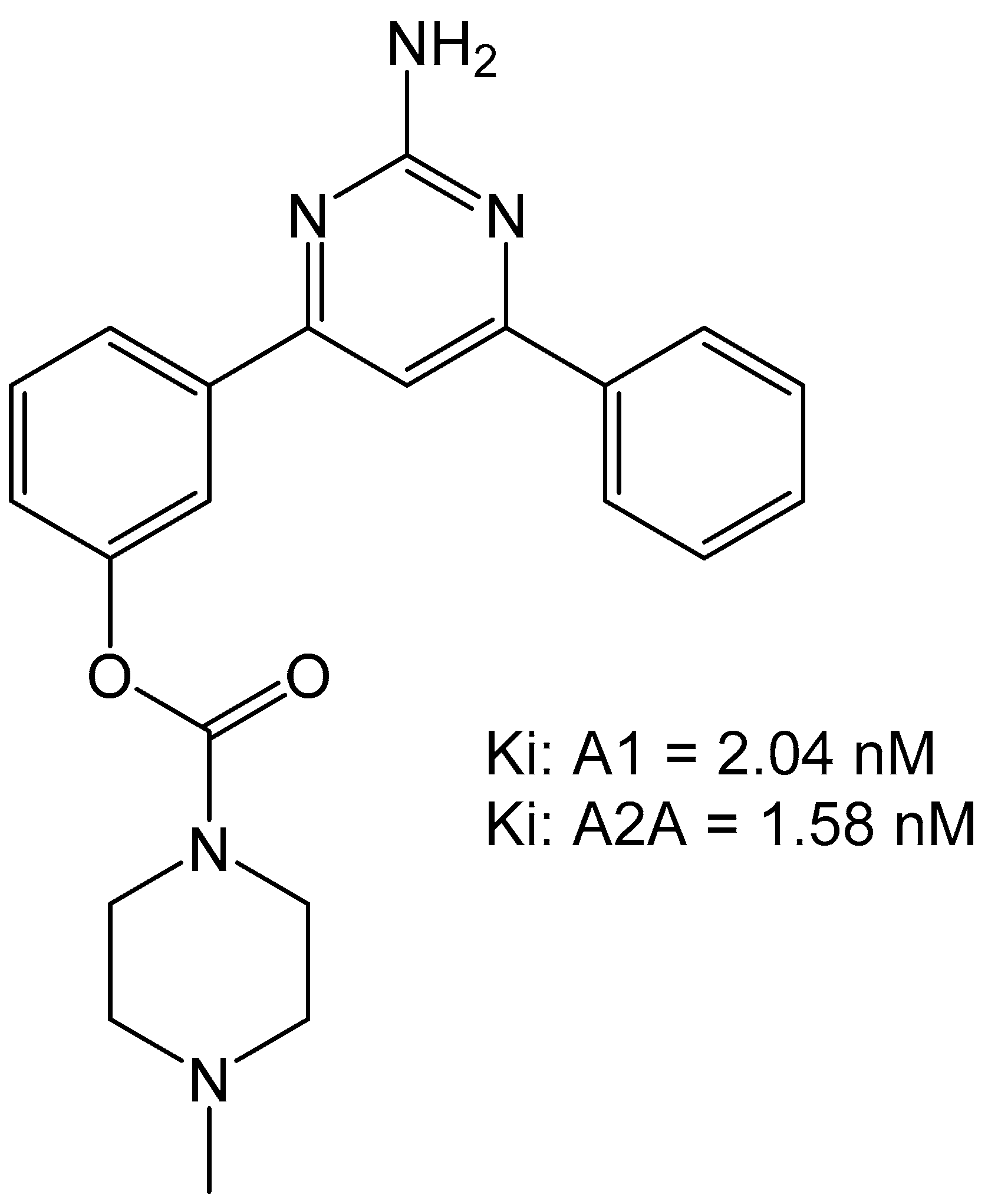
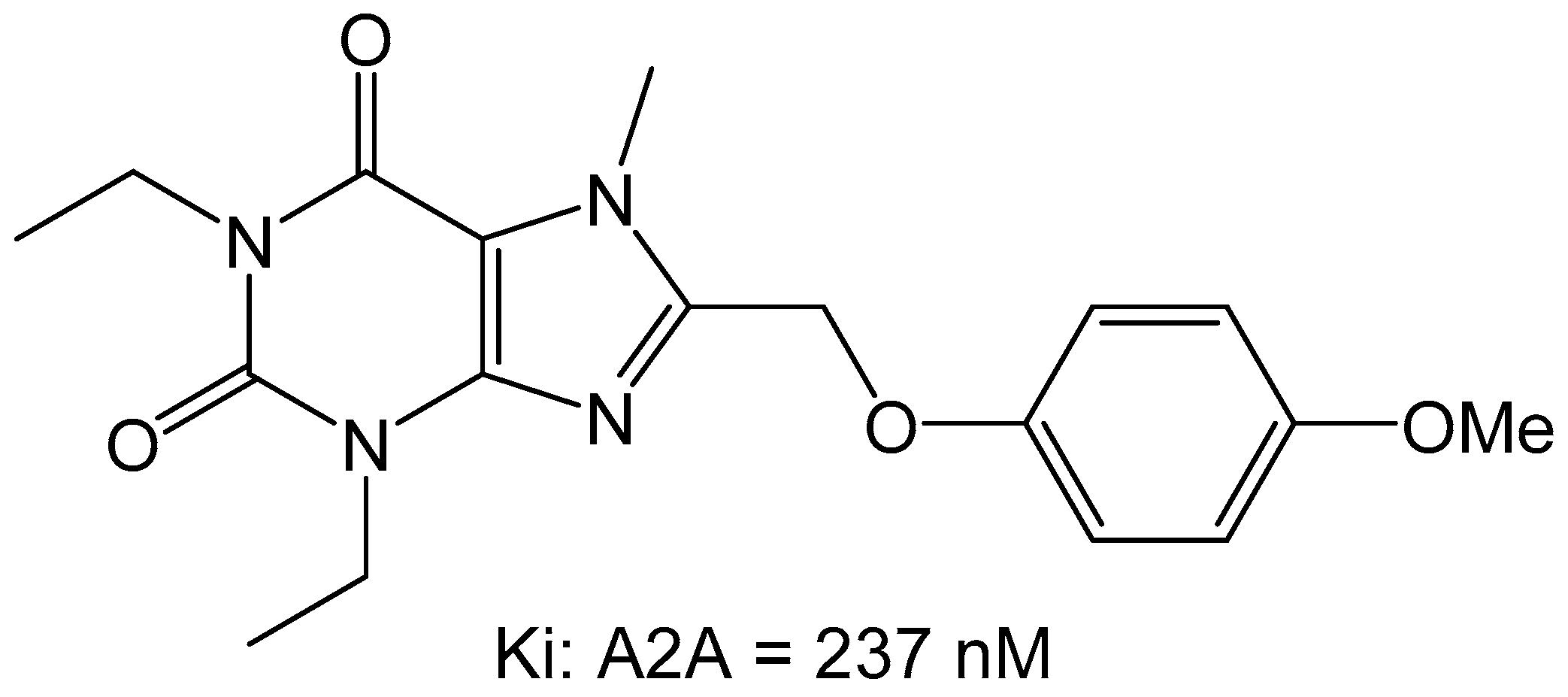
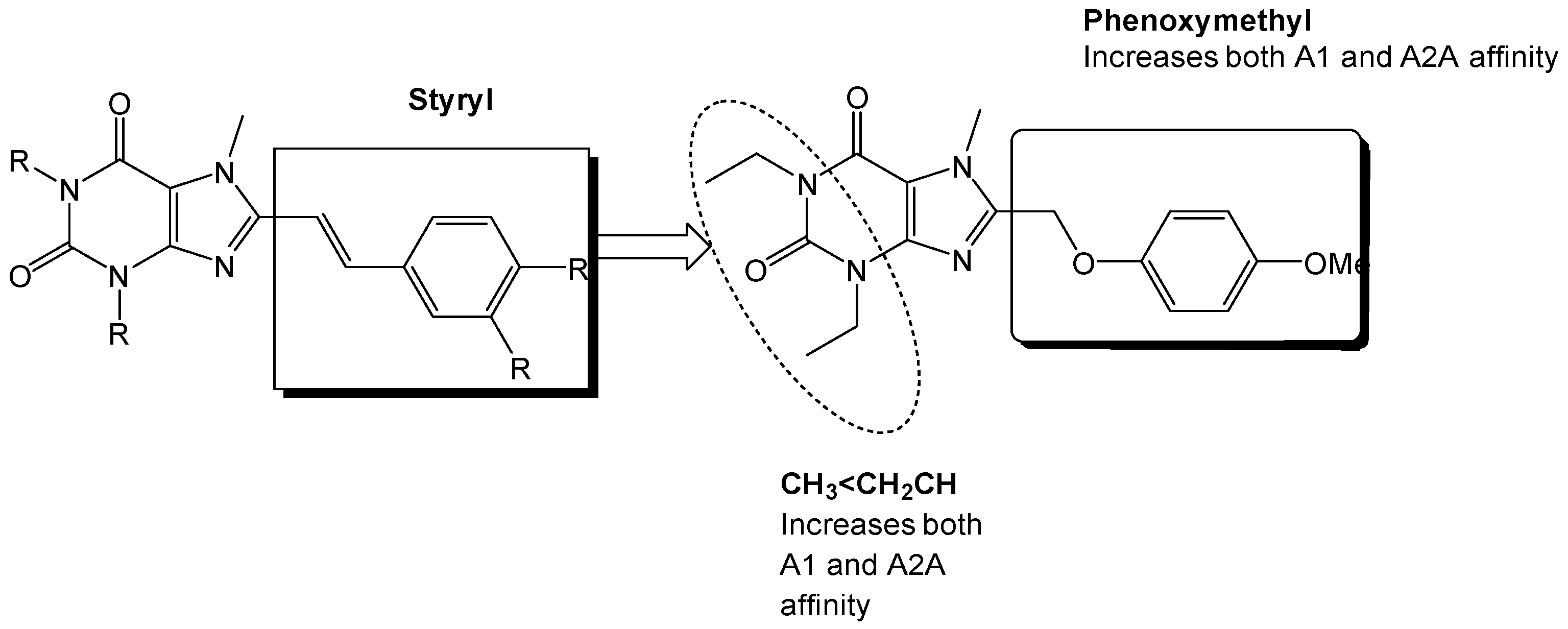
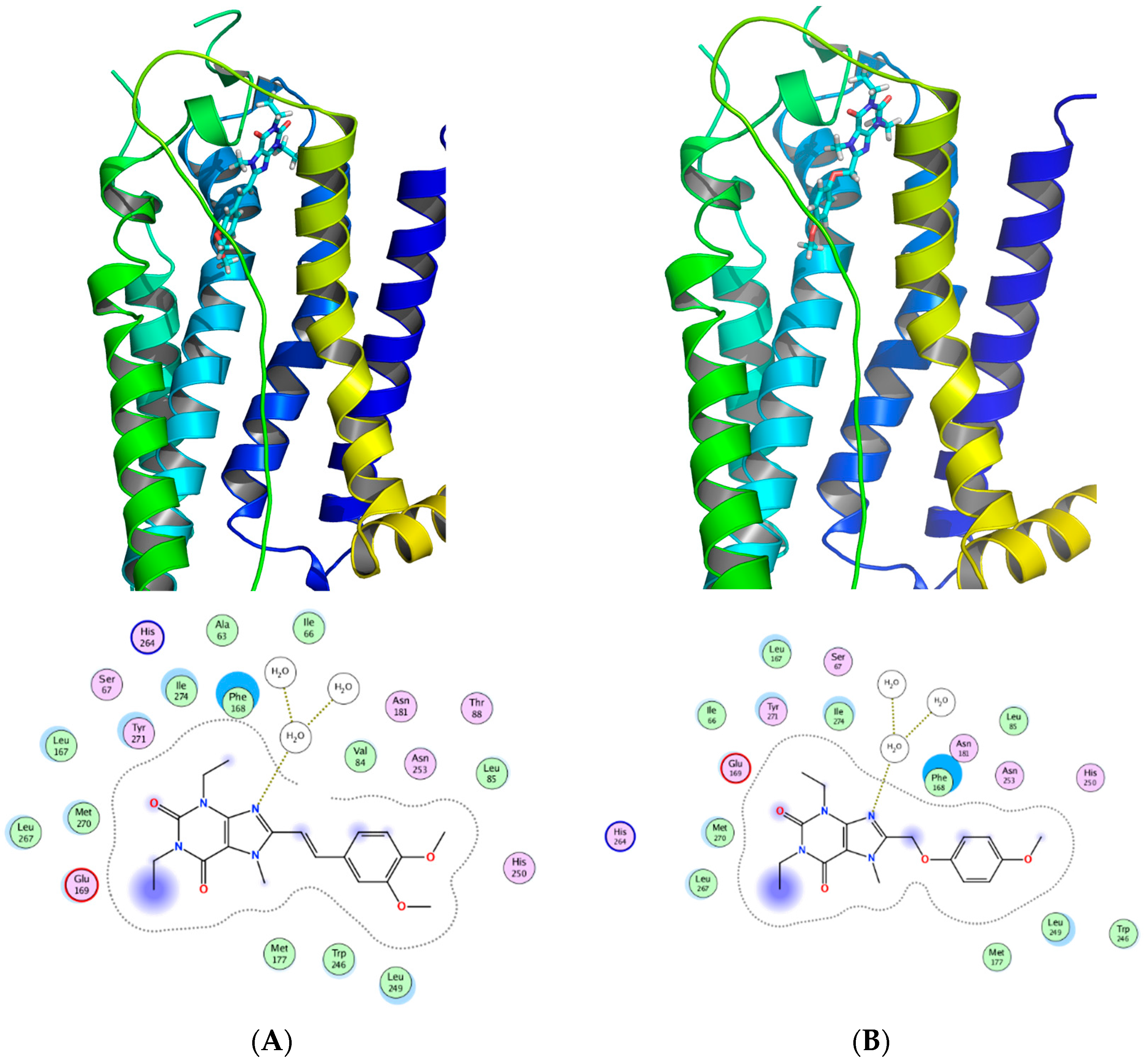
3. Medicinal Chemistry
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Gillespie, C.D.; Hurvitz, K.A. Prevalence of Uncontrolled Risk Factors for Cardiovascular Disease: United States, 1999–2010. NCHS Data Brief 2012, 103, 1–8. [Google Scholar]
- Mustafa, S.J. Cellular and molecular mechanism(s) of coronary flow regulation by adenosine. Mol. Cell. Biochem. 1980, 31, 67–87. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.F.; Eltzschig, H.K.; Fredholm, B.B. Adenosine receptors as drug targets—What are the challenges? Nat. Rev. Drug Discov. 2013, 12, 265–286. [Google Scholar] [CrossRef] [PubMed]
- Delacretaz, E. Clinical practice. Supraventricular tachycardia. N. Engl. J. Med. 2006, 354, 1039–1051. [Google Scholar] [CrossRef] [PubMed]
- Verzijl, D.; Ijzerman, A.P. Functional selectivity of adenosine receptor ligands. Purinergic Signal. 2011, 7, 171–192. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.R.; Nayeem, M.A.; Tilley, S.L.; Mustafa, S.J. Angiotensin II stimulation alters vasomotor response to adenosine in mouse mesenteric artery: Role for A1 and A2B adenosine receptors. Br. J. Pharmacol. 2015, 172, 4959–4969. [Google Scholar] [CrossRef] [PubMed]
- Leiva, A.; Guzman-Gutierrez, E.; Contreras-Duarte, S.; Fuenzalida, B.; Cantin, C.; Carvajal, L.; Salsoso, R.; Gutierrez, J.; Pardo, F.; Sobrevia, L. Adenosine receptors: Modulators of lipid availability that are controlled by lipid levels. Mol. Asp. Med. 2017. [Google Scholar] [CrossRef] [PubMed]
- Peleli, M.; Carlstrom, M. Adenosine signaling in diabetes mellitus and associated cardiovascular and renal complications. Mol. Asp. Med. 2017. [Google Scholar] [CrossRef] [PubMed]
- Atef, R.M.; Agha, A.M.; Abdel-Rhaman, A.A.; Nassar, N.N. The Ying and Yang of Adenosine A1 and A2A Receptors on ERK1/2 Activation in a Rat Model of Global Cerebral Ischemia Reperfusion Injury. Mol. Neurobiol. 2017, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Langemeijer, E.V.; Verzijl, D.; Dekker, S.J.; Ijzerman, A.P. Functional selectivity of adenosine A1 receptor ligands? Purinergic Signal. 2013, 9, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Kazemi, M.H.; Raoufi, S.; Hojjat-Farsangi, M.; Anvari, E.; Ghalamfarsa, G.; Mohammadi, H.; Jadidi-Niaragh, F. Adenosine and Adenosine Receptors in the Immunopathogenesis and Treatment of Cancer. J. Cell. Physiol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Kull, B.; Svenningsson, P.; Fredholm, B.B. Adenosine A(2A) receptors are colocalized with and activate g(olf) in rat striatum. Mol. Pharmacol. 2000, 58, 771–777. [Google Scholar] [PubMed]
- Nayeem, M.A.; Mustafa, S.J. Mechanisms of delayed preconditioning with A1 adenosine receptor activation in porcine coronary smooth muscle cells. Pol. J. Pharmacol. 2002, 54, 443–453. [Google Scholar] [PubMed]
- Nayeem, M.A.; Mustafa, S.J. Protein kinase C isoforms and A1 adenosine receptors in porcine coronary smooth muscle cells. Vasc. Pharmacol. 2002, 39, 47–54. [Google Scholar] [CrossRef]
- Nayeem, M.A.; Matherne, G.P.; Mustafa, S.J. Ischemic and pharmacological preconditioning induces further delayed protection in transgenic mouse cardiac myocytes over-expressing adenosine A1 receptors (A1AR): Role of A1AR, iNOS and K(ATP) channels. Naunyn Schmiedeberg Arch. Pharmacol. 2003, 367, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Nayeem, M.A.; Mustafa, S.J. Ischemic and pharmacological induction of delayed cellular protection in iNOS gene-disrupted mice myocytes. Pol. J. Pharmacol. 2003, 55, 73–79. [Google Scholar] [PubMed]
- Nayeem, M.A.; Poloyac, S.M.; Falck, J.R.; Zeldin, D.C.; Ledent, C.; Ponnoth, D.S.; Ansari, H.R.; Mustafa, S.J. Role of CYP epoxygenases in A2A AR-mediated relaxation using A2A AR-null and wild-type mice. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H2068–H2078. [Google Scholar] [CrossRef] [PubMed]
- Nayeem, M.A.; Ponnoth, D.S.; Boegehold, M.A.; Zeldin, D.C.; Falck, J.R.; Mustafa, S.J. High-salt diet enhances mouse aortic relaxation through adenosine A2A receptor via CYP epoxygenases. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R567–R574. [Google Scholar] [CrossRef] [PubMed]
- Nayeem, M.A.; Zeldin, D.C.; Boegehold, M.A.; Morisseau, C.; Marowsky, A.; Ponnoth, D.S.; Roush, K.P.; Falck, J.R. Modulation by salt intake of the vascular response mediated through adenosine A(2A) receptor: Role of CYP epoxygenase and soluble epoxide hydrolase. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, R325–R333. [Google Scholar] [CrossRef] [PubMed]
- Nayeem, M.A.; Zeldin, D.C.; Boegehold, M.A.; Falck, J.R. Salt modulates vascular response through adenosine A(2A) receptor in eNOS-null mice: Role of CYP450 epoxygenase and soluble epoxide hydrolase. Mol. Cell. Biochem. 2011, 350, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Ponnoth, D.S.; Nayeem, M.A.; Tilley, S.L.; Ledent, C.; Jamal Mustafa, S. CYP-epoxygenases contribute to A2A receptor-mediated aortic relaxation via sarcolemmal KATP channels. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 303, R1003–R1010. [Google Scholar] [CrossRef] [PubMed]
- Kunduri, S.S.; Mustafa, S.J.; Ponnoth, D.S.; Dick, G.M.; Nayeem, M.A. Adenosine A1 receptors link to smooth muscle contraction via CYP4a, protein kinase C-alpha, and ERK1/2. J. Cardiovasc. Pharmacol. 2013, 62, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Kunduri, S.; Dick, G.; Nayeem, M.; Mustafa, S. Adenosine A1 receptor signaling inhibits BK channels through a PKCalpha-dependent mechanism in mouse aortic smooth muscle. Physiol. Rep. 2013, 1, e00037. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.R.; Hong, K.L.; Zeldin, D.C.; Nayeem, M.A. Vascular endothelial over-expression of soluble epoxide hydrolase (Tie2-sEH) enhances adenosine A1 receptor-dependent contraction in mouse mesenteric arteries: Role of ATP-sensitive K+ channels. Mol. Cell. Biochem. 2016, 422, 197–206. [Google Scholar] [CrossRef] [PubMed]
- De Lera Ruiz, M.; Lim, Y.H.; Zheng, J. Adenosine A2A receptor as a drug discovery target. J. Med. Chem. 2014, 57, 3623–3650. [Google Scholar] [CrossRef] [PubMed]
- Jaakola, V.P.; Griffith, M.T.; Hanson, M.A.; Cherezov, V.; Chien, E.Y.; Lane, J.R.; Ijzerman, A.P.; Stevens, R.C. The 2.6 angstrom crystal structure of a human A2A adenosine receptor bound to an antagonist. Science 2008, 322, 1211–1217. [Google Scholar] [CrossRef] [PubMed]
- Glukhova, A.; Thal, D.M.; Nguyen, A.T.; Vecchio, E.A.; Jorg, M.; Scammells, P.J.; May, L.T.; Sexton, P.M.; Christopoulos, A. Structure of the Adenosine A1 Receptor Reveals the Basis for Subtype Selectivity. Cell 2017, 168, 867–877. [Google Scholar] [CrossRef] [PubMed]
- Almerico, A.M.; Tutone, M.; Pantano, L.; Lauria, A. A3 adenosine receptor: Homology modeling and 3D-QSAR studies. J. Mol. Graph. Model. 2013, 42, 60–72. [Google Scholar] [CrossRef] [PubMed]
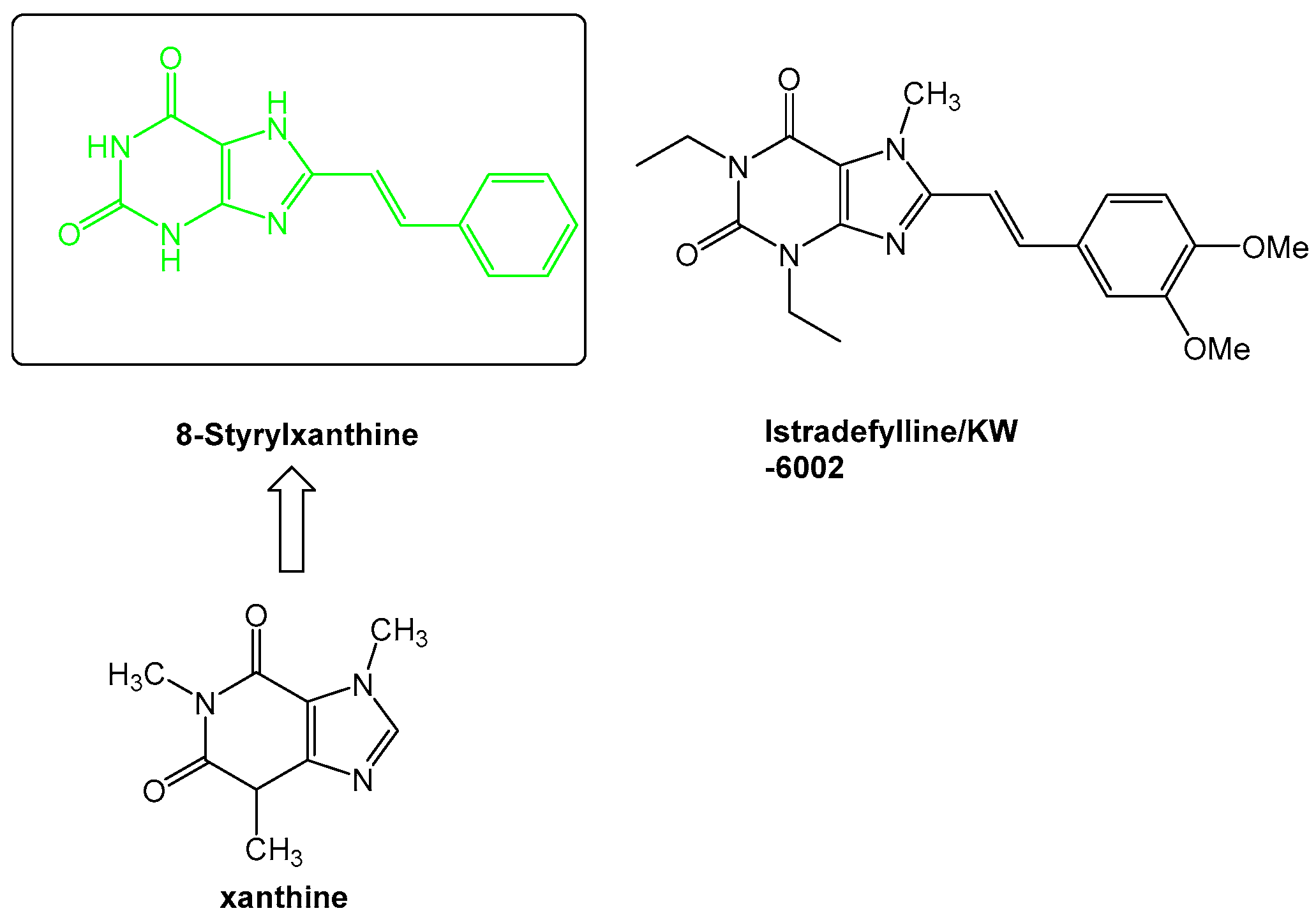
- Dungo, R.; Deeks, E.D. Istradefylline: First global approval. Drugs 2013, 73, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Petzer, J.P.; Steyn, S.; Castagnoli, K.P.; Chen, J.F.; Schwarzschild, M.A.; Van der Schyf, C.J.; Castagnoli, N. Inhibition of monoamine oxidase B by selective adenosine A2A receptor antagonists. Bioorg. Med. Chem. 2003, 11, 1299–1310. [Google Scholar] [CrossRef]
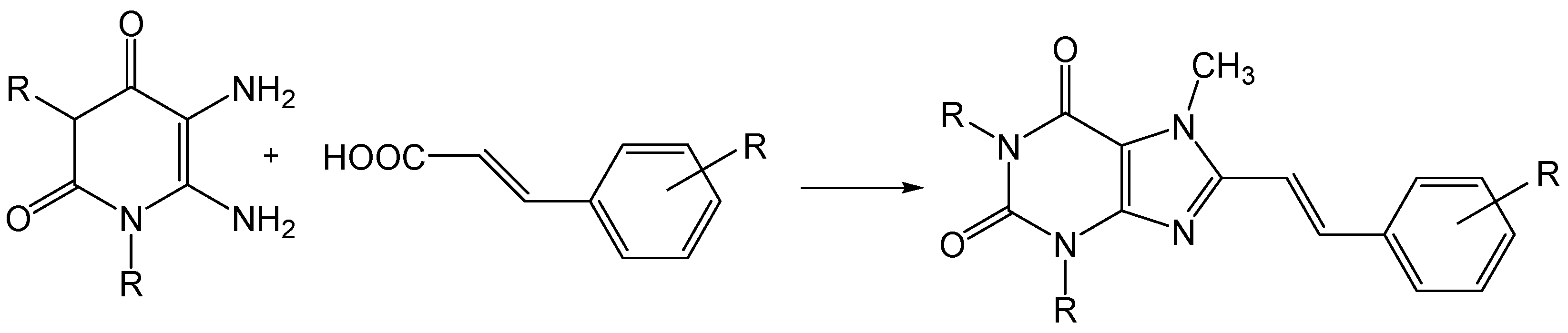
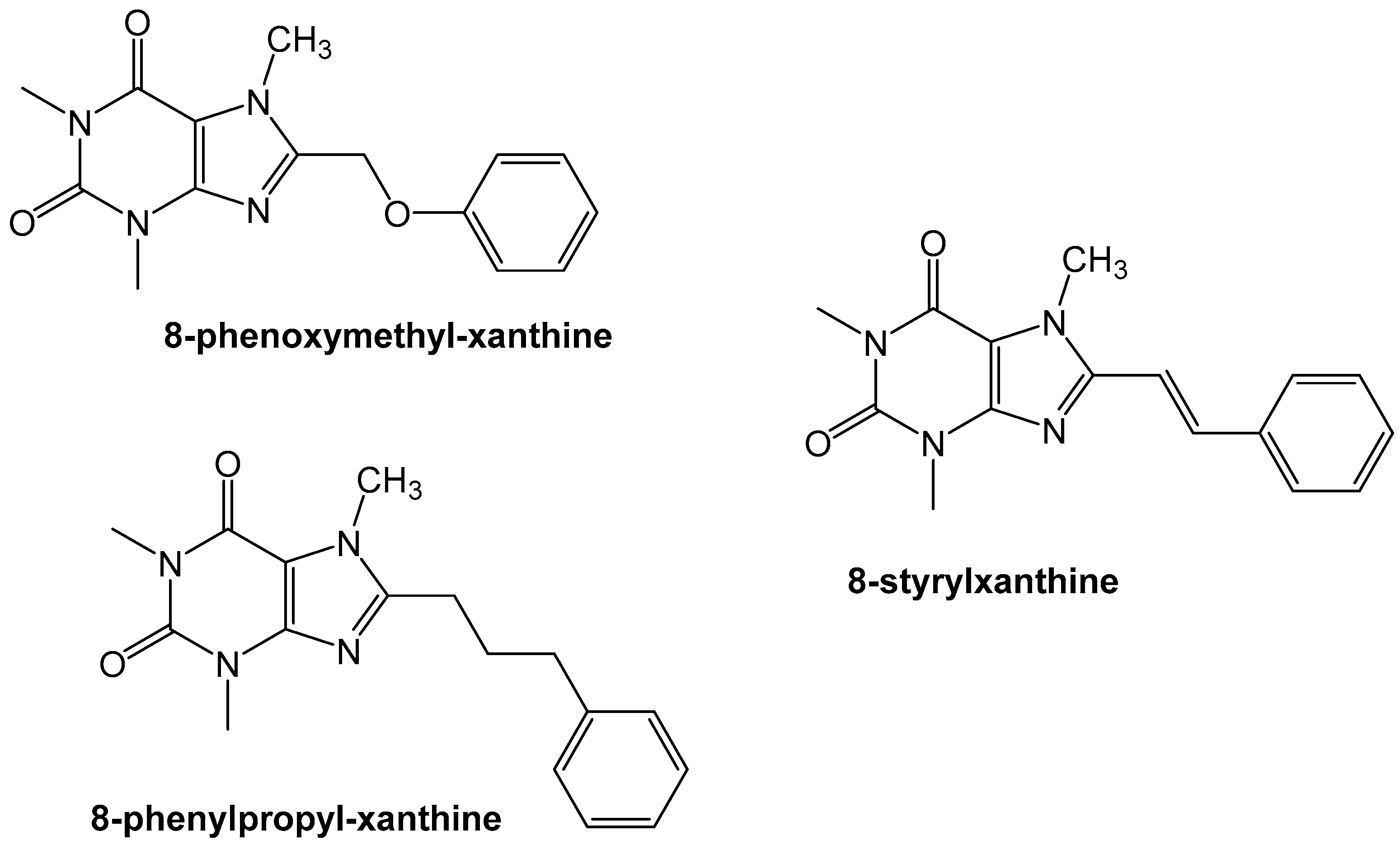
- Van der Walt, M.M.; Terre’Blanche, G.; Petzer, A.; Lourens, A.C.; Petzer, J.P. The adenosine A(2A) antagonistic properties of selected C8-substituted xanthines. Bioorg. Chem. 2013, 49, 49–58. [Google Scholar] [CrossRef] [PubMed]
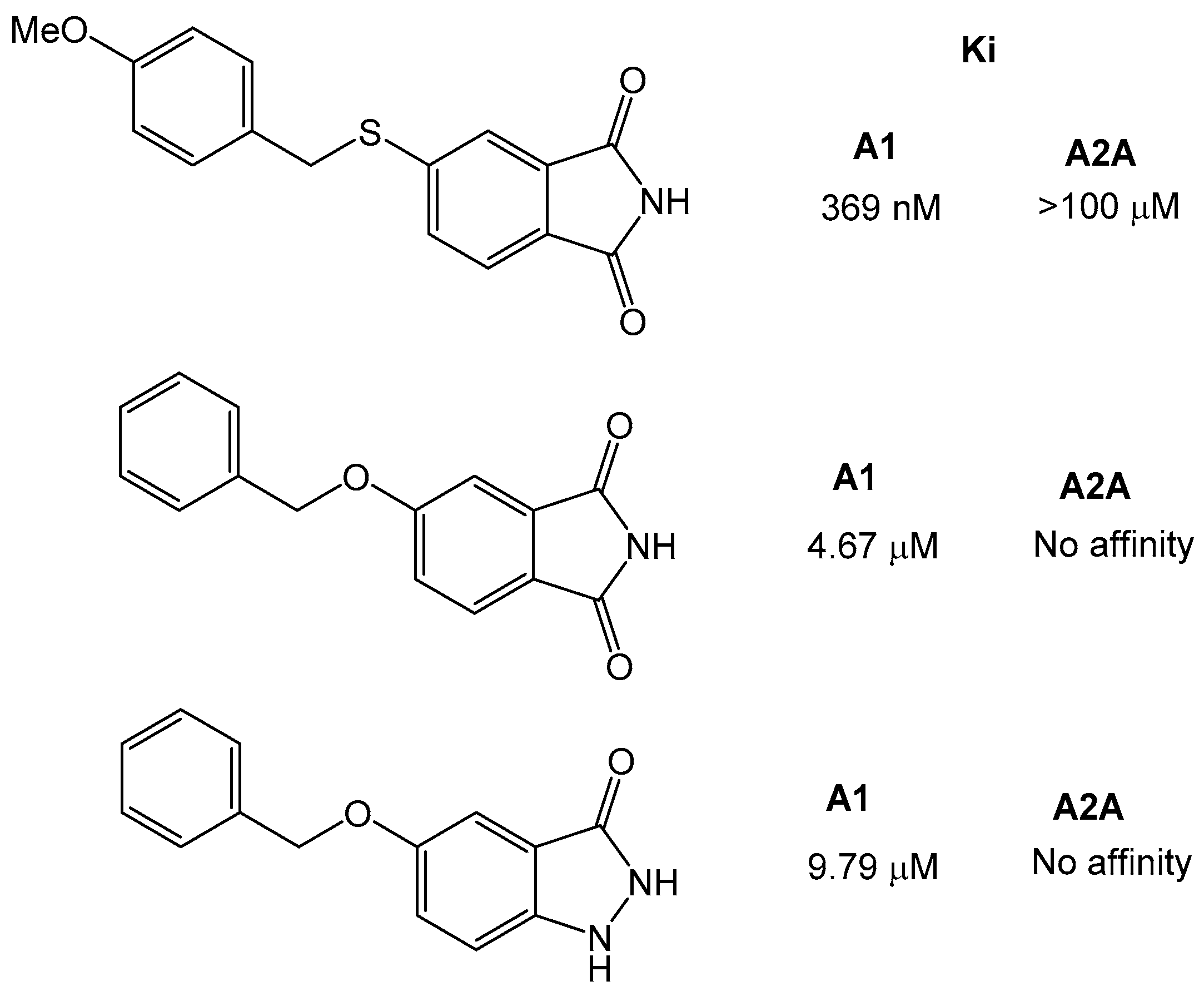
- Van der Walt, M.M.; Terre’Blanche, G.; Petzer, A.; Petzer, J.P. The adenosine receptor affinities and monoamine oxidase B inhibitory properties of sulfanylphthalimide analogues. Bioorg. Chem. 2015, 59, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.J.; Petzer, J.P.; Terre’Blanche, G.; Petzer, A.; van der Walt, M.M.; Bergh, J.J.; Lourens, A.C. 2-Aminopyrimidines as dual adenosine A1/A2A antagonists. Eur. J. Med. Chem. 2015, 104, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.J.; Petzer, J.P.; Rousseau, A.L.; Terre’Blanche, G.; Petzer, A.; Lourens, A.C. Carbamate substituted 2-amino-4,6-diphenylpyrimidines as adenosine receptor antagonists. Bioorg. Med. Chem. Lett. 2016, 26, 734–738. [Google Scholar] [CrossRef] [PubMed]
- Harmse, R.; van der Walt, M.M.; Petzer, J.P.; Terre’Blanche, G. Discovery of 1,3-diethyl-7-methyl-8-(phenoxymethyl)-xanthine derivatives as novel adenosine A1 and A2A receptor antagonists. Bioorg. Med. Chem. Lett. 2016, 26, 5951–5955. [Google Scholar] [CrossRef] [PubMed]
- Chemical Computing Group. Available online: www.chemcomp.com (accessed on 25 May 2017).














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A1 | A2A |
|---|---|
| 5UEN | 3EML |
| 3RFM | |
| 2YDO | |
| 2YDV |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geldenhuys, W.J.; Hanif, A.; Yun, J.; Nayeem, M.A. Exploring Adenosine Receptor Ligands: Potential Role in the Treatment of Cardiovascular Diseases. Molecules 2017, 22, 917. https://doi.org/10.3390/molecules22060917
Geldenhuys WJ, Hanif A, Yun J, Nayeem MA. Exploring Adenosine Receptor Ligands: Potential Role in the Treatment of Cardiovascular Diseases. Molecules. 2017; 22(6):917. https://doi.org/10.3390/molecules22060917
Chicago/Turabian StyleGeldenhuys, Werner J., Ahmad Hanif, June Yun, and Mohammed A. Nayeem. 2017. "Exploring Adenosine Receptor Ligands: Potential Role in the Treatment of Cardiovascular Diseases" Molecules 22, no. 6: 917. https://doi.org/10.3390/molecules22060917
APA StyleGeldenhuys, W. J., Hanif, A., Yun, J., & Nayeem, M. A. (2017). Exploring Adenosine Receptor Ligands: Potential Role in the Treatment of Cardiovascular Diseases. Molecules, 22(6), 917. https://doi.org/10.3390/molecules22060917