Soluble Epoxide Hydrolase Inhibitory Activity of Components Isolated from Apios americana Medik



Abstract
:
1. Introduction
2. Results and Discussion
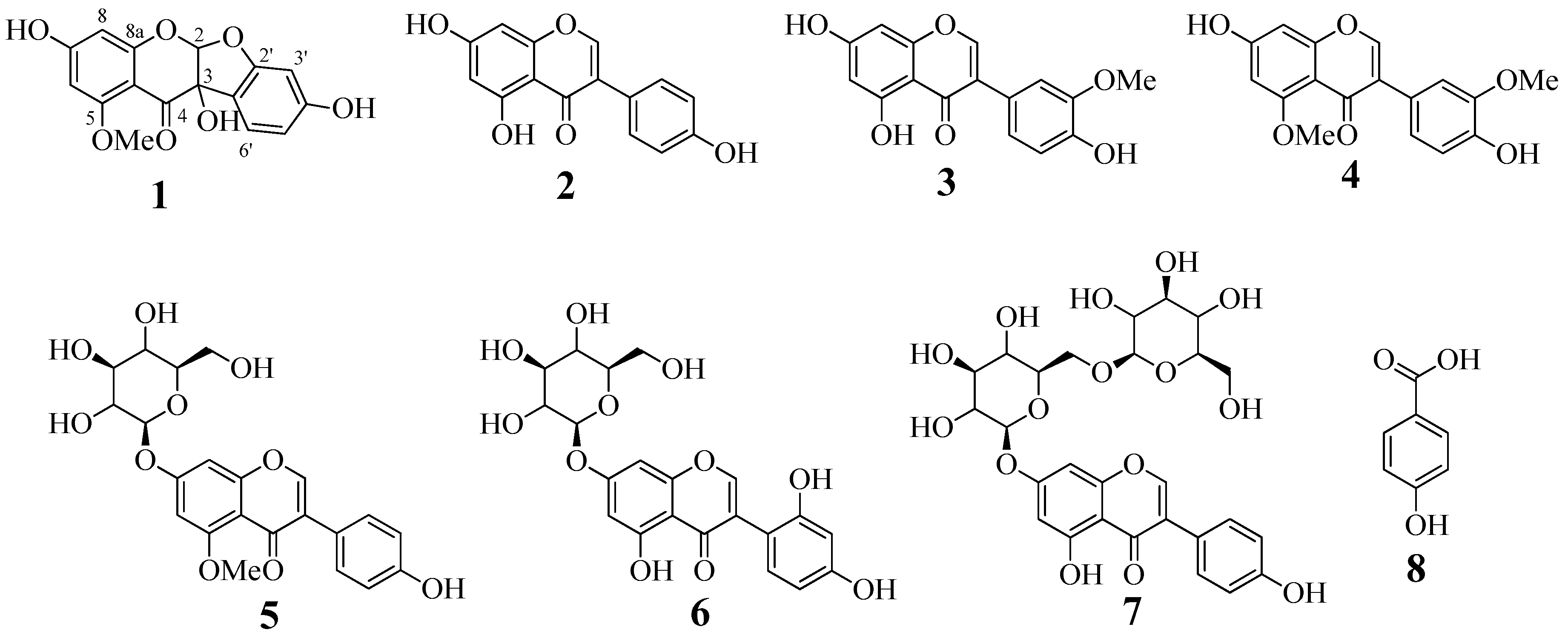
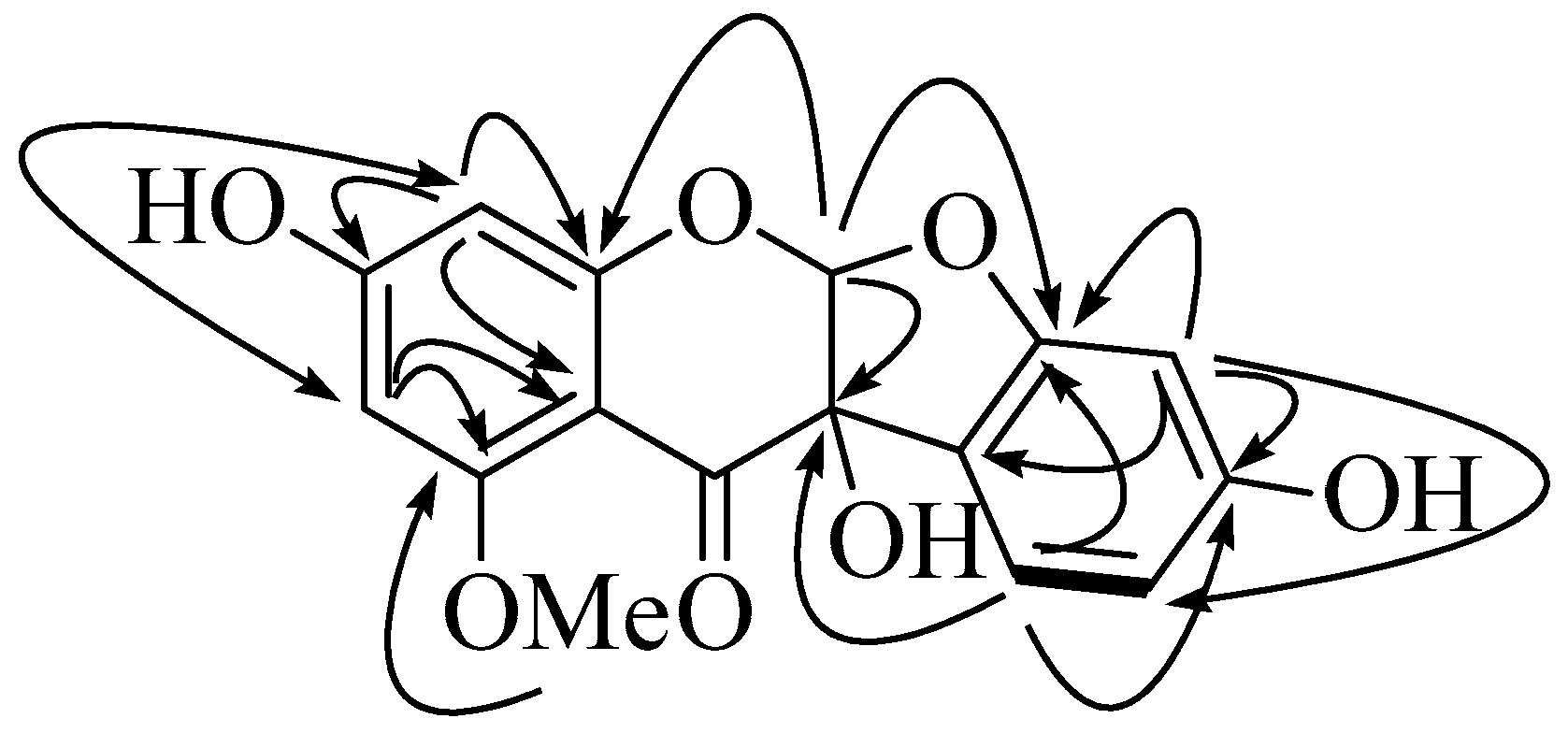
2.1. Isolation and Identification
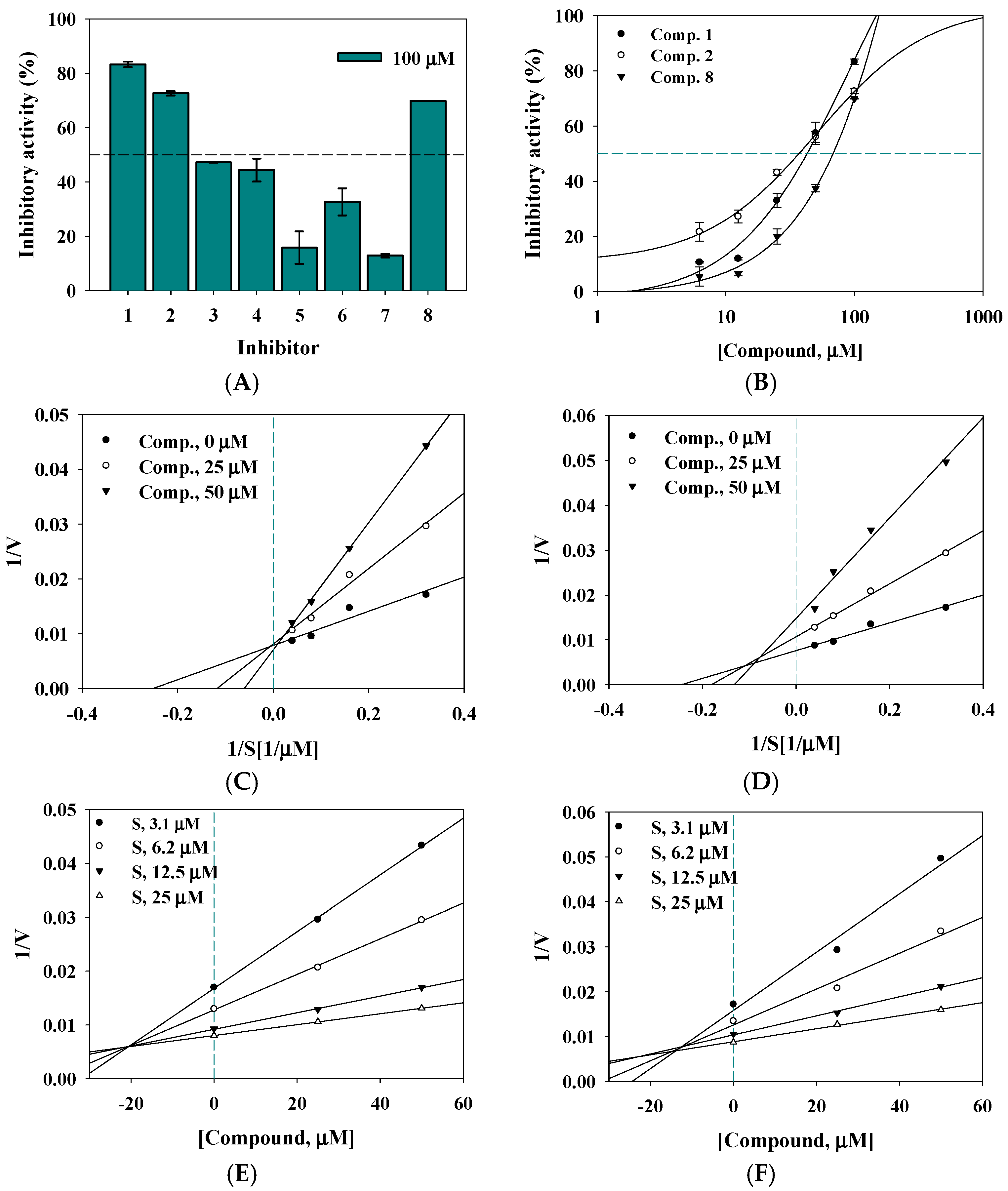
2.2. Enzyme Activity
2.3. Enzyme Kinetics
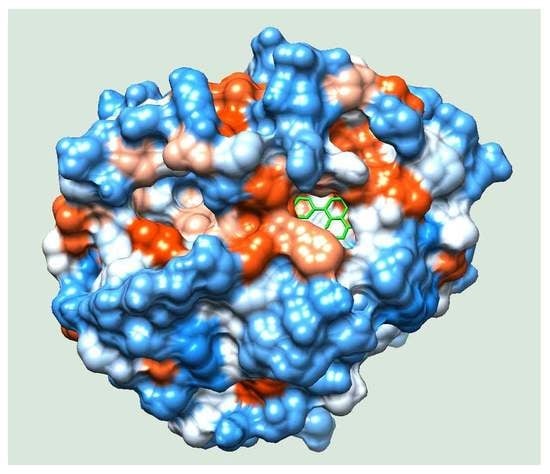
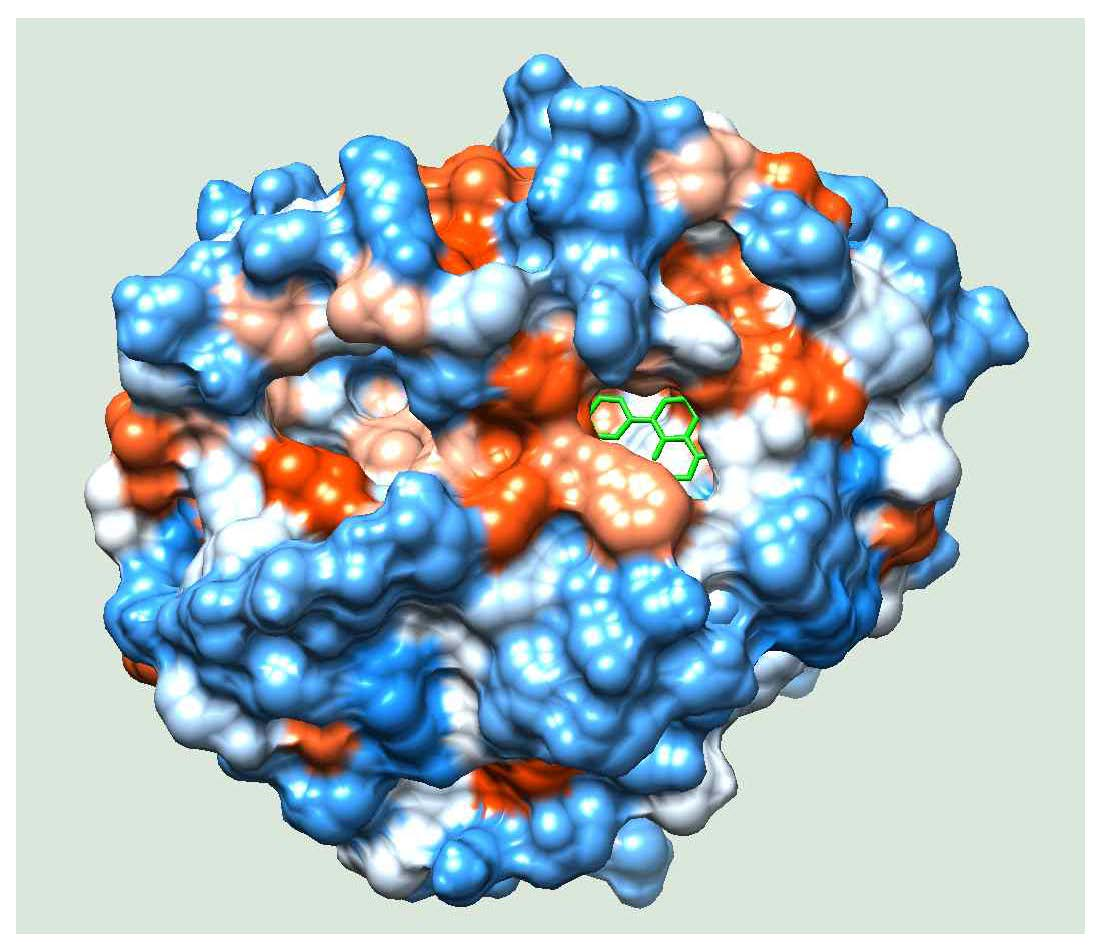
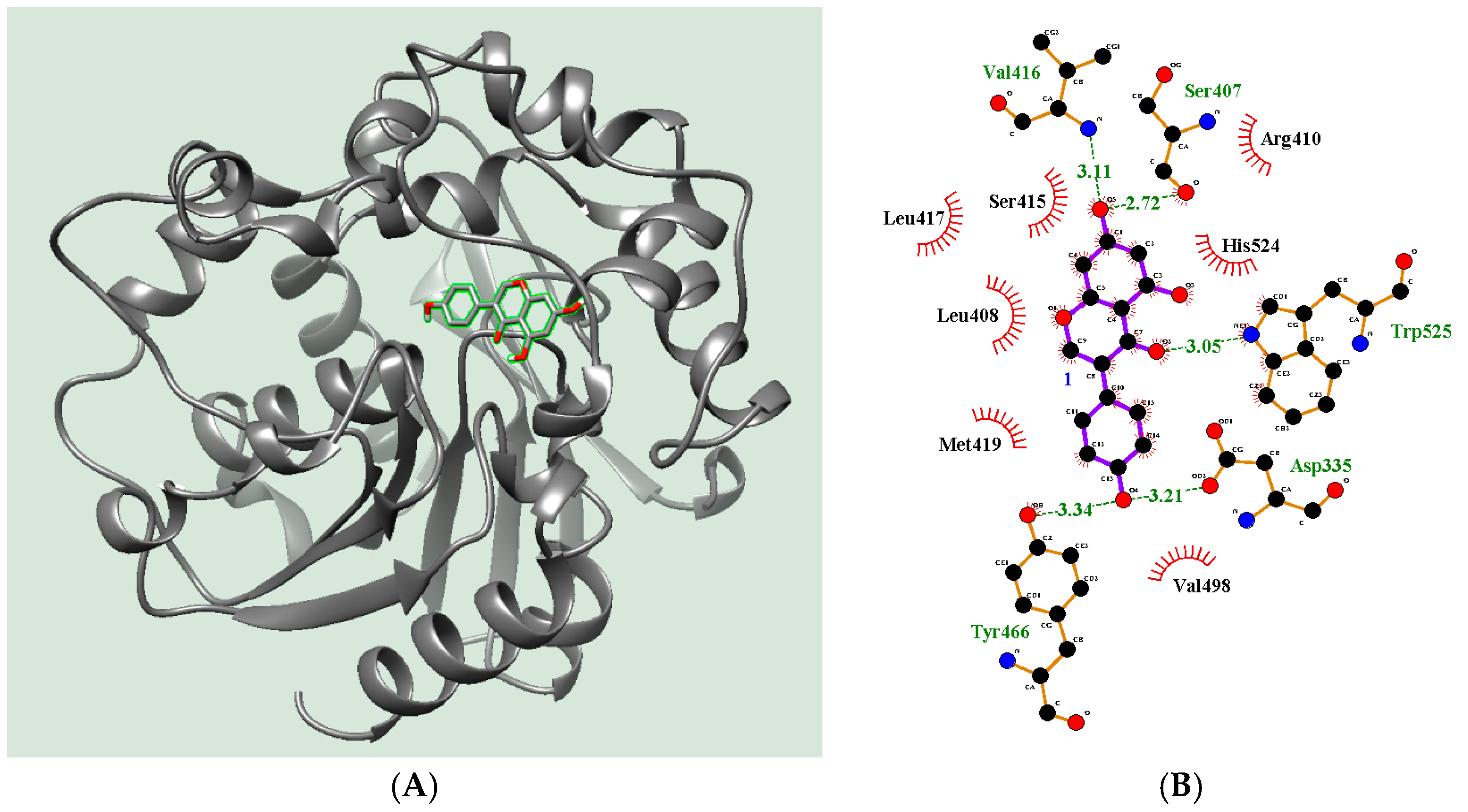
2.4. Molecular Docking
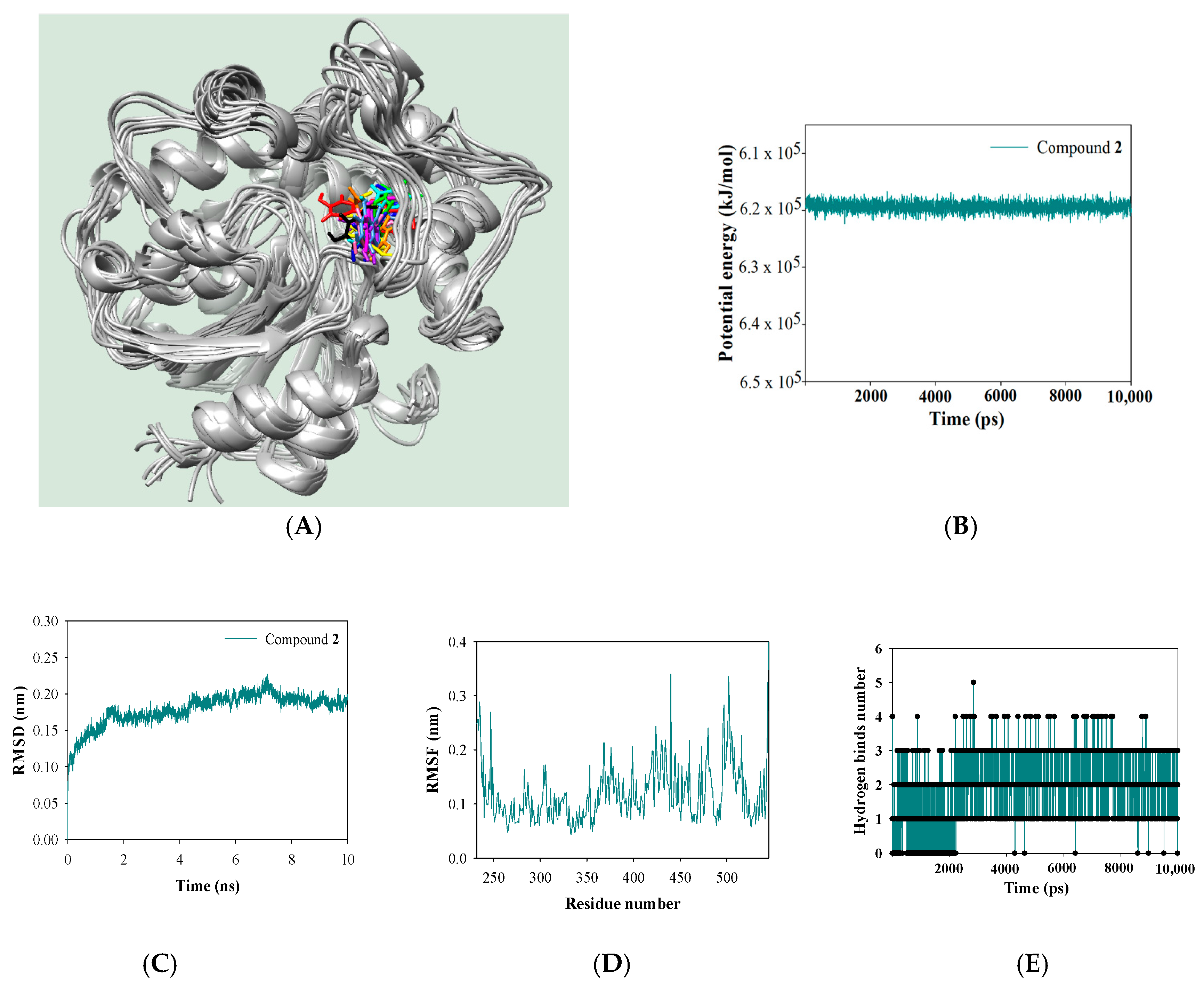
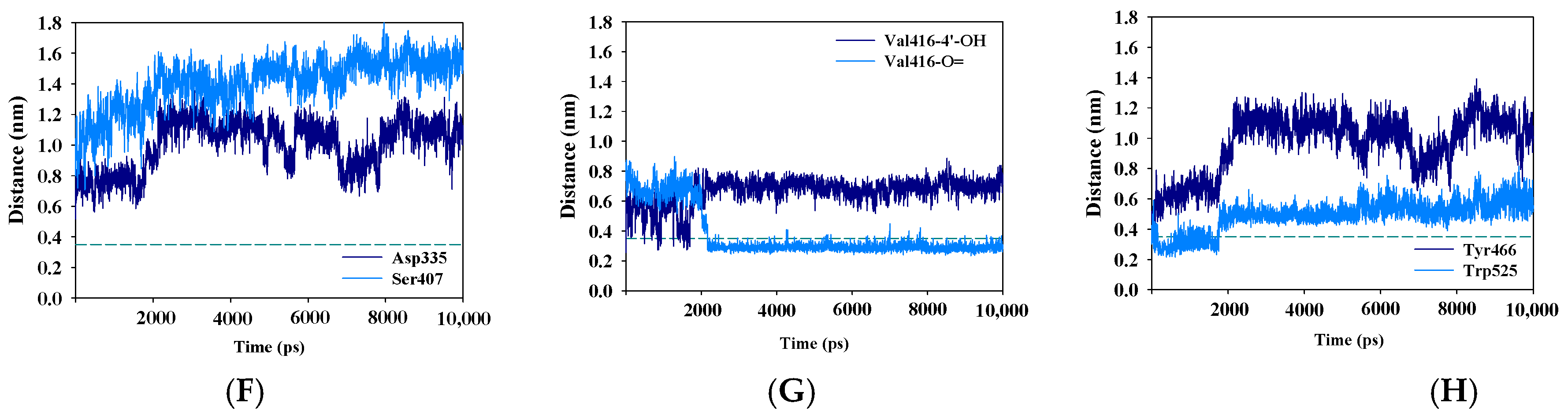
2.5. Molecular Simulation
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Plant Material
3.3. Extraction and Isolation
3.4. sEH Assay
3.5. Molecular Docking
3.6. Molecular Dynamics
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nara, K.; Nihei, K.-I.; Ogasawara, Y.; Koga, H.; Kato, Y. Novel isoflavone diglycoside in groundnut (Apios. americana Medik). Food Chem. 2011, 124, 703–710. [Google Scholar] [CrossRef]
- Nakahashi, H.; Nishino, Y.; Nakagawa, H.; Hara, N.; Usami, A.; Miyazawa, M. Evaluation of the key odorants in volatile oils from tubers of Apios. americana Medikus. J. Oleo Sci. 2015, 64, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Ichige, M.; Fukuda, E.; Miida, S.; Hattan, J.-I.; Misawa, N.; Saito, S.; Fujimaki, T.; Masaya, I.; Shindo, K. Novel isoflavone glucosides in groundnut (Apios. americana Medik) and their antiandrogenic activities. J. Agric. Food Chem. 2013, 61, 2183–2187. [Google Scholar] [CrossRef] [PubMed]
- Kenmochi, E.; Kabir, S.R.; Ogawa, T.; Naude, R.; Tateno, H.; Hirabayashi, J.; Muramoto, K. Isolation and biochemical characterization of Apios. tuber lectin. Molecules 2015, 20, 987–1002. [Google Scholar] [CrossRef] [PubMed]
- Kaneta, H.; Koda, M.; Saito, S.; Imoto, M.; Kawada, M.; Yamazaki, Y.; Momose, I.; Shindo, K. Biological activities of unique isoflavones prepared from Apios. americana Medik. Biosci. Biotechnol. Biochem. 2016, 80, 774–778. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Tai, B.H.; Yang, S.Y.; Kim, J.E.; Kim, S.K.; Kim, Y.H. Soluble epoxide hydrolase inhibitory constituents from Selaginella. tamariscina. Bull. Korean Chem. Soc. 2015, 36, 300–304. [Google Scholar] [CrossRef]
- Kim, J.H.; Cho, C.W.; Tai, B.H.; Yang, S.Y.; Choi, G.-S.; Kang, J.S.; Kim, Y.H. Soluble epoxide hydrolase inhibitory activity of selaginellin from Selaginella. tamariscina. Molecules 2015, 20, 21405–21414. [Google Scholar] [CrossRef] [PubMed]
- Jo, A.R.; Kim, J.H.; Yan, X.-T.; Yang, S.Y.; Kim, Y.H. Soluble epoxide hydrolase inhibitory components from Rheum undulatum and in silico approach. J. Enzym. Inhib. Med. Chem. 2016, 31, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Borhan, B.; Jones, A.D.; Pinot, F.; Grant, D.F.; Kurth, M.J.; Hammock, B.D. Mechanism of Soluble Epoxide Hydrolase formation of an α-hydroxy ester-enzyme intermediate through Asp-333. J. Biol. Chem. 1995, 270, 26923–26930. [Google Scholar] [CrossRef] [PubMed]
- Fretland, A.J.; Omiecinski, C.J. Epoxide hydrolases: Biochemistry and molecular biology. Chem. Biol. Interact. 2000, 129, 41–59. [Google Scholar] [CrossRef]
- Lee, G.Y.; Kim, J.H.; Choi, S.-K.; Kim, Y.H. Constituents of the seeds of Cassia tora with inhibitory activity on soluble expoxide hydrolease. Bioorg. Med. Chem. Lett. 2015, 25, 5097–5101. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Morgan, A.M.A.; Tai, B.H.; Van, D.T.; Cuong, N.M.; Kim, Y.H. Inhibition of soluble epoxide hydrolase activity by compounds isolated from the aerial parts of Glycosmis stenocarpa. J. Enzym. Inhib. Med. Chem. 2016, 31, 640–644. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Ryu, Y.B.; Lee, W.S.; Kim, Y.H. Neuraminidase inhibitory activities of quaternary isoquinoline alkaloids from Corydalis turtschaninovii rhizome. Bioorg. Med. Chem. 2014, 22, 6047–6052. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, S.-H.; Lee, H.W.; Sun, Y.N.; Jang, W.-H.; Yang, S.-Y.; Jang, H.-D.; Kim, Y.H. (−)-Epicatechin derivate from Orostachys japonicus as potential inhibitor of the human butyrylcholinesterase. Int. J. Biol. Macromolec. 2016, 91, 1033–1039. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Li, Q.; Xu, J. Chemical composition of roots flemingia philippinensis and their inhibitory kinetics on aromatase. Chem. Biodivers. 2017, 14, e1600193. [Google Scholar] [CrossRef] [PubMed]
- Sircar, D.; Roychowdhury, A.; Mitra, A.J. Accumulation of p-hydroxybenzoic acid in hairy roots of Daucus carota. J. Plant. Physiol. 2007, 164, 1358–1366. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Duan, J.-A.; Che, C.-T. Isoflavanones and their O-glycosides from Desmodium styracifolium. Phytochemistry 2007, 68, 1471–1479. [Google Scholar] [CrossRef] [PubMed]
- Thao, N.P.; Luyen, B.T.T.; Kim, J.H.; Jo, A.R.; Dat, N.T.; Kiem, P.V.; Minh, C.V.; Kim, Y.H. Identification, characterization, kinetics, and molecular docking of flavonoid constituents from Archidendron clypearia (Jack.) Nielsen leaves and twigs. Bioorg. Med. Chem. 2016, 24, 3125–3132. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | δH a,b | δC a,c |
|---|---|---|
| 2 | 6.14 (1H, s) | 122.5 |
| 3 | 82.2 | |
| 4 | 189.0 | |
| 4a | 104.9 | |
| 5 | 163.8 | |
| 6 | 6.03 (1H, d, J = 2.2 Hz) | 98.4 |
| 7 | 167.3 | |
| 8 | 6.11 (1H, d, J = 2.2 Hz) | 95.8 |
| 8a | 161.9 | |
| 1′ | 119.2 | |
| 2′ | 163.0 | |
| 3′ | 6.28 (1H, d, J = 1.5 Hz) | 98.7 |
| 4′ | 162.4 | |
| 5′ | 6.36 (1H, dd, J = 8.4, 1.5 Hz) | 110.6 |
| 6′ | 7.08 (1H, d, J = 8.4 Hz) | 126.5 |
| 5-OMe | 3.76 (3H, s) | 56.8 |
| The Inhibitory Activity on sEH | ||
|---|---|---|
| IC50 (μM) a | Binding mode (Ki, μM) a | |
| 1 | 43.2 ± 0.4 | Competitive (21.0 ± 0.8) |
| 2 | 33.5 ± 0.8 | Mixed (12.4 ± 1.5) |
| 8 | 69.3 ± 1.9 | N.T c |
| AUDA b | 7.6 ± 2.5 nM | |
| Hydrogen Bonds (Å) | Binding Energy (kcal/mol) | |
|---|---|---|
| 2 | Asp335 (3.21), Ser407 (2.72), Val416 (3.11), Tyr466 (3.34), Trp525 (3.05) | −8.02 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.H.; Kim, H.Y.; Kang, S.Y.; Kim, Y.H.; Jin, C.H. Soluble Epoxide Hydrolase Inhibitory Activity of Components Isolated from Apios americana Medik. Molecules 2017, 22, 1432. https://doi.org/10.3390/molecules22091432
Kim JH, Kim HY, Kang SY, Kim YH, Jin CH. Soluble Epoxide Hydrolase Inhibitory Activity of Components Isolated from Apios americana Medik. Molecules. 2017; 22(9):1432. https://doi.org/10.3390/molecules22091432
Chicago/Turabian StyleKim, Jang Hoon, Hyo Young Kim, Si Yong Kang, Young Ho Kim, and Chang Hyun Jin. 2017. "Soluble Epoxide Hydrolase Inhibitory Activity of Components Isolated from Apios americana Medik" Molecules 22, no. 9: 1432. https://doi.org/10.3390/molecules22091432
APA StyleKim, J. H., Kim, H. Y., Kang, S. Y., Kim, Y. H., & Jin, C. H. (2017). Soluble Epoxide Hydrolase Inhibitory Activity of Components Isolated from Apios americana Medik. Molecules, 22(9), 1432. https://doi.org/10.3390/molecules22091432