An Overview of LEDs’ Effects on the Production of Bioactive Compounds and Crop Quality



Abstract
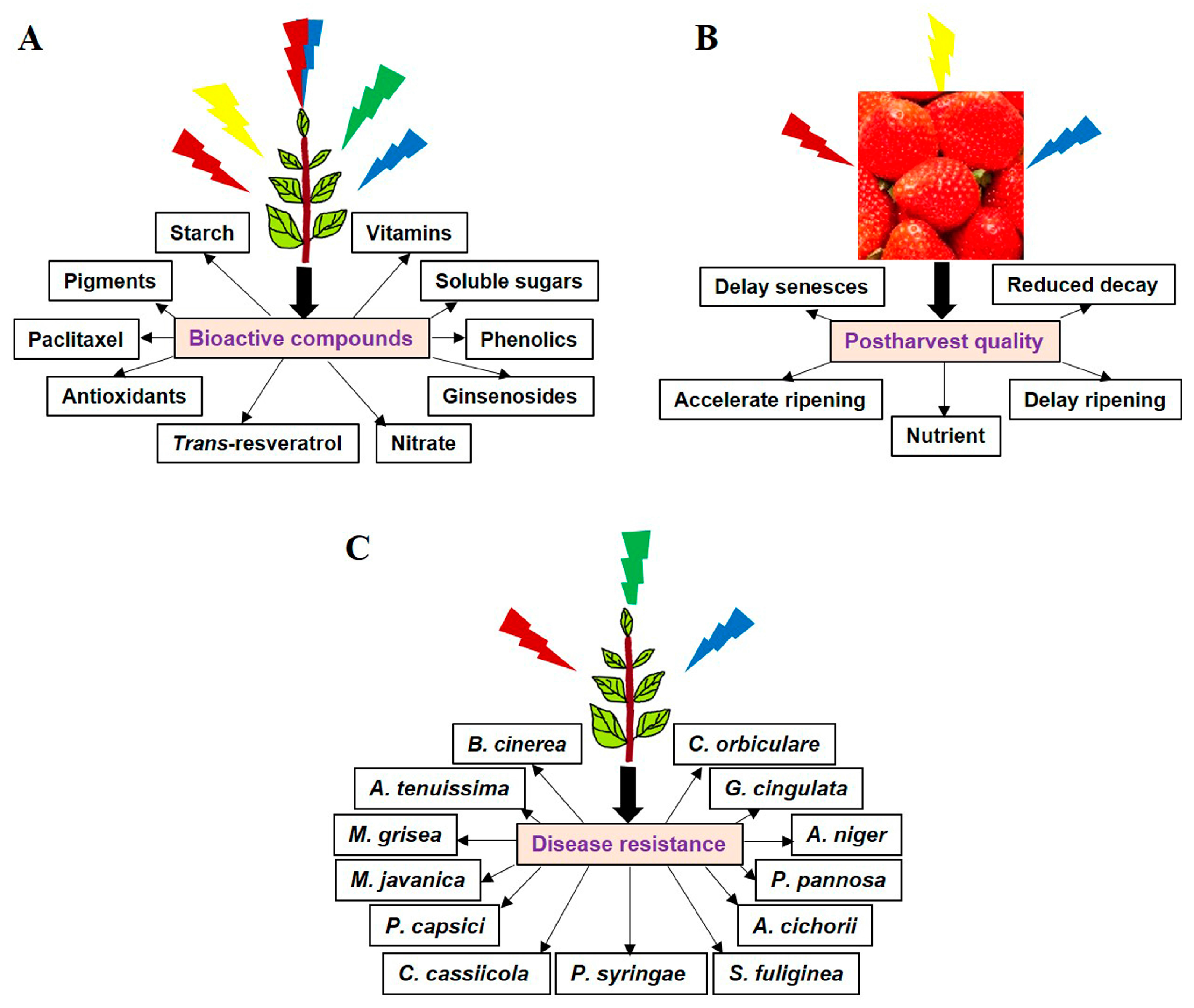
:1. Introduction
2. LEDs Induce Bioactive Compound Synthesis in Crops
3. LEDs Enhance Antioxidant Properties
4. LEDs Improve Nutritional Traits of the Postharvest Produce
5. LEDs Offer Protection against Food Spoilage and Crop Loss
6. Role of LEDs in Increasing Crop Yield
7. Conclusions and Future Prospects
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Quail, P.H.; Boylan, M.T.; Parks, B.M.; Short, T.W.; Xu, Y.; Wagner, D. Phytochromes: Photosensory perception and signal transduction. Science 1995, 268, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.W.; Quail, P.H. Signalling in light-controlled development. Semin. Cell Dev. Biol. 1999, 10, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Braidot, E.; Petrussa, E.; Peresson, C.; Patui, S.; Bertolini, A.; Tubaro, F.; Wahlby, U.; Coan, M.; Vianello, A.; Zancani, M. Low-intensity light cycles improve the quality of lamb’s lettuce (Valerianella olitoria [L.] Pollich) during storage at low temperature. Postharvest Biol. Technol. 2014, 90, 15–30. [Google Scholar] [CrossRef]
- Costa, L.; Montano, Y.M.; Carrion, C.; Rolny, N.; Guiamet, J.J. Application of low intensity light pulses to delay postharvest senescence of Ocimum basilicum leaves. Postharvest Biol. Technol. 2013, 86, 181–191. [Google Scholar] [CrossRef]
- Glowacz, M.; Mogren, L.M.; Reade, J.P.H.; Cobb, A.H.; Monaghan, J.M. High-but not low-intensity light leads to oxidative stress and quality loss of cold-stored baby leaf spinach. J. Sci. Food Agric. 2014, 95, 1821–1829. [Google Scholar] [CrossRef] [PubMed]
- Kozai, T.; Fujiwara, K.; Runkle, E.S. LED Lighting for Urban Agriculture; Springer Science+Business Media: Singapore, 2016. [Google Scholar]
- Pogson, B.J.; Morris, S.C. Postharvest senescence of vegetables and its regulation. In Plant Cell Death Processes; Nood’en, L.D., Ed.; Academic Press: San Diego, CA, USA, 2004; pp. 319–329. [Google Scholar]
- Hopkins, W.G.; Hüner, N.P.A. Secondary metabolites. In Introduction to Plant Physiology, 4th ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2008. [Google Scholar]
- Hasan, M.M.; Baek, K.H. Induction of resveratrol biosynthesis in grape skin and leaves by ultrasonication treatment. Korean J. Hortic. Sci. Technol. 2013, 31, 496–502. [Google Scholar] [CrossRef]
- Hasan, M.M.; Yun, H.K.; Kwak, E.J.; Baek, K.H. Preparation of resveratrol-enriched grape juice from ultrasonication treated grape fruits. Ultrason. Sonochem. 2014, 21, 729–734. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.M.; Bae, H. An Overview of stress-induced resveratrol synthesis in grapes: Perspectives for resveratrol-enriched grape products. Molecules 2017, 22, 294. [Google Scholar] [CrossRef]
- Hasan, M.M.; Bashir, T.; Bae, H. Use of ultrasonication technology for the increased production of plant secondary metabolites. Molecules 2017, 22, 1046. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Ghate, V.; Kim, M.J.; Zhou, W.B.; Khoo, G.H.; Yuk, H.G. Kinetics of bacterial inactivation by 405 nm and 520 nm light emitting diodes and the role of endogenous coproporphyrin on bacterial susceptibility. J. Photochem. Photobiol. B Biol. 2015, 149, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Lubart, R.; Lipovski, A.; Nitzan, Y.; Friedmann, H. A possible mechanism for the bactericidal effect of visible light. Laser Ther. 2011, 20, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.A.; Both, A.-J.; Bourget, C.M.; Burr, J.F.; Kubota, C.; Lopez, R.G.; Morrow, R.C.; Runkle, E.S. LEDs: The future of greenhouse lighting! Chron. Hortic. 2012, 52, 5–13. [Google Scholar]
- Morrow, R.C. LED lighting in horticulture. Hortscience 2008, 43, 1947–1950. [Google Scholar]
- Branas, C.; Azcondo, F.J.; Alonso, J.M. Solid-state lighting a system review. IEEE Ind. Electron. Mag. 2013, 7, 6–14. [Google Scholar] [CrossRef]
- Yeh, N.C.; Chung, J.P. High-brightness LEDs-Energy efficient lighting sources and their potential in indoor plant cultivation. Renew. Sustain. Energy Rev. 2009, 13, 2175–2180. [Google Scholar] [CrossRef]
- Bian, Z.H.; Yang, Q.C.; Liu, W.K. Effects of light quality on the accumulation of phytochemicals in vegetables produced in controlled environments: A review. J. Sci. Food Agric. 2015, 95, 869–877. [Google Scholar] [CrossRef]
- Britz, S.J.; Sager, J.C. Photomorphogenesis and photoassimilation in soybean and sorghum grown under broad spectrum or blue-deficient light sources. Plant Physiol. 1990, 94, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Li, H.M.; Xu, Z.G.; Tang, C.M. Effect of light-emitting diodes on growth and morphogenesis of upland cotton (Gossypium hirsutum L.) plantlets in vitro. Plant Cell Tissue Org. Cult. 2010, 103, 155–163. [Google Scholar] [CrossRef]
- Soebo, A.; Krekling, T.; Applegren, M. Light quality affects photosynthesis and leaf anatomy of birch plantlets in vitro. Plant Cell Tissue Org. Cult. 1995, 41, 177–185. [Google Scholar] [CrossRef]
- Kim, K.; Kook, H.-S.; Jang, Y.-J.; Lee, W.-H.; Kamala-Kannan, S.; Chae, J.-C.; Lee, K.-J. The effect of blue-light emitting diodes on antioxidant properties and resistance to Botrytis cinerea in tomato. J. Plant Pathol. Microbiol. 2013, 4, 203. [Google Scholar]
- Johkan, M.; Shoji, K.; Goto, F.; Hashida, S.; Yoshihara, T. Blue light-emitting diode light irradiation of seedlings improves seedling quality and growth after transplanting in red leaf lettuce. Hortscience 2010, 45, 1809–1814. [Google Scholar]
- Li, H.M.; Tang, C.M.; Xu, Z.G.; Liu, X.Y.; Han, X.L. Effects of different light sources on the growth of non-heading Chinese cabbage (Brassica campestris L.). J. Agric. Sci. 2012, 4, 262–273. [Google Scholar] [CrossRef]
- Heo, J.W.; Kang, D.H.; Bang, H.S.; Hong, S.G.; Chun, C.; Kang, K.K. Early growth, pigmentation, protein content, and phenylalanine ammonia-lyase activity of red curled lettuces grown under different lighting conditions. Korean J. Hortic. Sci. Technol. 2012, 30, 6–12. [Google Scholar] [CrossRef]
- Heo, J.W.; Shin, K.S.; Kim, S.K.; Paek, K.Y. Light quality affects in vitro growth of grape ‘Teleki 5BB’. J. Plant Biol. 2006, 49, 276–280. [Google Scholar] [CrossRef]
- Shin, K.S.; Murthy, H.N.; Heo, J.W.; Hahn, E.J.; Paek, K.Y. The effect of light quality on the growth and development of in vitro cultured Doritaenopsis plants. Acta Physiol. Plant. 2008, 30, 339–343. [Google Scholar] [CrossRef]
- Lee, M.K.; Arasu, M.V.; Park, S.; Byeon, D.H.; Chung, S.O.; Park, S.U.; Lim, Y.P.; Kim, S.J. LED lights enhance metabolites and antioxidants in chinese cabbage and kale. Braz. Arch. Biol. Technol. 2016, 59, e16150546. [Google Scholar] [CrossRef]
- Lekkham, P.; Srilaong, V.; Pongprasert, N.; Kondo, S. Anthocyanin concentration and antioxidant activity in light-emitting diode (LED)-treated apples in a greenhouse environmental control system. Fruits 2016, 71, 269–274. [Google Scholar] [CrossRef]
- Choi, H.G.; Moon, B.Y.; Kang, N.J. Effects of LED light on the production of strawberry during cultivation in a plastic greenhouse and in a growth chamber. Sci. Hortic. 2015, 189, 22–31. [Google Scholar] [CrossRef]
- Samuoliene, G.; Brazaityte, A.; Sirtautas, R.; Virsile, A.; Sakalauskaite, J.; Sakalauskiene, S.; Duchovskis, P. LED illumination affects bioactive compounds in romaine baby leaf lettuce. J. Sci. Food Agric. 2013, 93, 3286–3291. [Google Scholar] [CrossRef] [PubMed]
- Samuoliene, G.; Sirtautas, R.; Brazaityte, A.; Duchovskis, P. LED lighting and seasonality effects antioxidant properties of baby leaf lettuce. Food Chem. 2012, 134, 1494–1499. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Fu, Y.; Liu, G.; Liu, H. Growth, photosynthetic characteristics, antioxidant capacity and biomass yield and quality of wheat (Triticum aestivum L.) Exposed to LED light sources with different spectra combinations. J. Agron. Crop Sci. 2014, 200, 219–230. [Google Scholar] [CrossRef]
- Lin, K.H.; Huang, M.Y.; Huang, W.D.; Hsu, M.H.; Yang, Z.W.; Yang, C.M. The effects of red, blue, and white light-emitting diodes on the growth, development, and edible quality of hydroponically grown lettuce (Lactuca sativa L. var. capitata). Sci. Hortic. 2013, 150, 86–91. [Google Scholar] [CrossRef]
- Bantis, F.; Ouzounis, T.; Radoglou, K. Artificial LED lighting enhances growth characteristics and total phenolic content of Ocimum basilicum, but variably affects transplant success. Sci. Hortic. 2016, 198, 277–283. [Google Scholar] [CrossRef]
- Ahn, S.Y.; Kim, S.A.; Yun, H.K. Inhibition of Botrytis cinerea and accumulation of stilbene compounds by light-emitting diodes of grapevine leaves and differential expression of defense-related genes. Eur. J. Plant Pathol. 2015, 143, 753–765. [Google Scholar] [CrossRef]
- Wu, M.C.; Hou, C.Y.; Jiang, C.M.; Wang, Y.T.; Wang, C.Y.; Chen, H.H.; Chang, H.M. A novel approach of LED light radiation improves the antioxidant activity of pea seedlings. Food Chem. 2007, 101, 1753–1758. [Google Scholar] [CrossRef]
- Ma, G.; Zhang, L.C.; Setiawan, C.K.; Yamawaki, K.; Asai, T.; Nishikawa, F.; Maezawa, S.; Sato, H.; Kanemitsu, N.; Kato, M. Effect of red and blue LED light irradiation on ascorbate content and expression of genes related to ascorbate metabolism in postharvest broccoli. Postharvest Biol. Technol. 2014, 94, 97–103. [Google Scholar] [CrossRef]
- Kook, H.S.; Park, S.H.; Jang, Y.J.; Lee, G.W.; Kim, J.S.; Kim, H.M.; Oh, B.T.; Chae, J.C.; Lee, K.J. Blue LED (light-emitting diodes)-mediated growth promotion and control of Botrytis disease in lettuce. Acta Agric. Scand. Sect. B Soil Plant Sci. 2013, 63, 271–277. [Google Scholar]
- Poudel, P.R.; Kataoka, I.; Mochioka, R. Effect of red- and blue-light-emitting diodes on growth and morphogenesis of grapes. Plant Cell Tissue Org. C. 2008, 92, 147–153. [Google Scholar] [CrossRef]
- Xu, F.; Cao, S.F.; Shi, L.Y.; Chen, W.; Su, X.G.; Yang, Z.F. Blue light irradiation affects anthocyanin content and enzyme activities involved in postharvest strawberry fruit. J. Agric. Food Chem. 2014, 62, 4778–4783. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Ha, J.Y.; Oh, J.E.; Cho, M.S. The effect of LED irradiation on the quality of cabbage stored at a low temperature. Food Sci. Biotechnol. 2014, 23, 1087–1093. [Google Scholar] [CrossRef]
- Mizuno, T.; Amaki, W.; Watanabe, H. Effects of monochromatic light irradiation by LED on the growth and anthocyanin contents in leaves of cabbage seedlings. In VI International Symposium on Light in Horticulture 907; Goto, E., Hikosaka, S., Eds.; ISHS: Leuven, Belgium, 2009; pp. 179–184. [Google Scholar]
- Dhakal, R.; Baek, K.-H. Metabolic alternation in the accumulation of free amino acids and γ-aminobutyric acid in postharvest mature green tomatoes following irradiation with blue light. Hortic. Environ. Biotechnol. 2014, 55, 36–41. [Google Scholar] [CrossRef]
- Shi, L.Y.; Cao, S.F.; Chen, W.; Yang, Z.F. Blue light induced anthocyanin accumulation and expression of associated genes in Chinese bayberry fruit. Sci. Hortic. 2014, 179, 98–102. [Google Scholar] [CrossRef]
- Gong, D.D.; Cao, S.F.; Sheng, T.; Shao, J.R.; Song, C.B.; Wo, F.C.; Chen, W.; Yang, Z.F. Effect of blue light on ethylene biosynthesis, signalling and fruit ripening in postharvest peaches. Sci. Hortic. 2015, 197, 657–664. [Google Scholar] [CrossRef]
- Liao, H.L.; Alferez, F.; Burns, J.K. Assessment of blue light treatments on citrus postharvest diseases. Postharvest Biol. Technol. 2013, 81, 81–88. [Google Scholar] [CrossRef]
- Alferez, F.; Liao, H.L.; Burns, J.K. Blue light alters infection by Penicillium digitatum in tangerines. Postharvest Biol. Technol. 2012, 63, 11–15. [Google Scholar] [CrossRef]
- Ballester, A.R.; Lafuente, M.T. LED Blue Light-induced changes in phenolics and ethylene in citrus fruit: Implication in elicited resistance against Penicillium digitatum infection. Food Chem. 2017, 218, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Park, S.U.; Ahn, D.J.; Jeon, H.J.; Kwon, T.R.; Lim, H.S.; Choi, B.S.; Baek, K.-H.; Bae, H. Increase in the contents of ginsenosides in raw ginseng roots in response to exposure to 450 and 470 nm light from light-emitting diodes. J. Ginseng Res. 2012, 36, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Nhut, D.T.; Nguyen, P.L.H.; Don, N.T.; Hien, N.T.T.; Huy, N.P.; Nam, N.B.; Vinh, B.T.; Luan, T.C. Induction, growth and paclitaxel content of needle-and petiole-derived calli in himalayan yew (Taxus Wallichiana Zucc.) under light-emitting diodes. Acta Biol. Crac. Ser. Bot. 2014, 56, 107–114. [Google Scholar] [CrossRef]
- Ahn, S.Y.; Kim, S.A.; Choi, S.J.; Yun, H.K. Comparison of accumulation of stilbene compounds and stilbene related gene expression in two grape berries irradiated with different light sources. Hortic. Environ. Biotechnol. 2015, 56, 36–43. [Google Scholar] [CrossRef]
- Kanazawa, K.; Hashimoto, T.; Yoshida, S.; Sungwon, P.; Fukuda, S. Short photo irradiation induces flavonoid synthesis and increases its production in postharvest vegetables. J. Agric. Food Chem. 2012, 60, 4359–4368. [Google Scholar] [CrossRef] [PubMed]
- Samuoliene, G.; Urbonaviciute, A.; Brazaityte, A.; Sabajeviene, G.; Sakalauskaite, J.; Duchovskis, P. The impact of LED illumination on antioxidant properties of sprouted seeds. Cent. Eur. J. Biol. 2011, 6, 68–74. [Google Scholar] [CrossRef]
- Kokalj, D.; Hribar, J.; Cigic, B.; Zlatic, E.; Demsar, L.; Sinkovic, L.; Sircelj, H.; Bizjak, G.; Vidrih, R. Influence of yellow light-emitting diodes at 590 nm on storage of apple, tomato and bell pepper fruit. Food Technol. Biotechnol. 2016, 54, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Colquhoun, T.A.; Schwieterman, M.L.; Gilbert, J.L.; Jaworski, E.A.; Langer, K.M.; Jones, C.R.; Rushing, G.V.; Hunter, T.M.; Olmstead, J.; Clark, D.G.; et al. Light modulation of volatile organic compounds from petunia flowers and select fruits. Postharvest Biol. Technol. 2013, 86, 37–44. [Google Scholar] [CrossRef]
- Muneer, S.; Kim, E.J.; Park, J.S.; Lee, J.H. Influence of green, red and blue light emitting diodes on multiprotein complex proteins and photosynthetic activity under different light intensities in lettuce leaves (Lactuca sativa L.). Int. J. Mol. Sci. 2014, 15, 4657–4670. [Google Scholar] [CrossRef] [PubMed]
- Massa, G.D.; Kim, H.H.; Wheeler, R.M.; Mitchell, C.A. Plant productivity in response to LED lighting. Hortscience 2008, 43, 1951–1956. [Google Scholar]
- D’Souza, C.; Yuk, H.-G.; Khoo, G.H.; Zhou, W. Application of light-emitting diodes in food production, postharvest preservation, and microbiological food safety. Compr. Rev. Food Sci. Food Saf. 2015, 14, 719–740. [Google Scholar] [CrossRef]
- Suthaparan, A.; Torre, S.; Stensvand, A.; Herrero, M.L.; Pettersen, R.I.; Gadoury, D.M.; Gislerod, H.R. Specific light-emitting diodes can suppress sporulation of Podosphaera pannosa on greenhouse roses. Plant Dis. 2010, 94, 1105–1110. [Google Scholar] [CrossRef]
- Vakalounakis, D.J.; Christias, C. Sporulation in Alternaria cichorii is controlled by a blue and near ultraviolet reversible photoreaction. Can. J. Bot. 1981, 59, 626–628. [Google Scholar] [CrossRef]
- Murdoch, L.E.; Mckenzie, K.; Maclean, M.; Macgregor, S.J.; Anderson, J.G. Lethal effects of high-intensity violet 405-nm light on Saccharomyces cerevisiae, Candida albicans, and on dormant and germinating spores of Aspergillus niger. Fungal Biol. 2013, 117, 519–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, S.Z.; Honda, Y.; Arase, S. Light-induced resistance of broad bean against Botrytis cinerea. J. Phytopathol. 1998, 146, 479–485. [Google Scholar] [CrossRef]
- Arase, S.; Fujita, K.; Uehara, T.; Honda, Y.; Isota, J. Light-enhanced resistance to Magnaporthe grisea infection in the rice Sekiguchi lesion mutants. J. Phytopathol. 2000, 148, 197–203. [Google Scholar] [CrossRef]
- Islam, S.Z.; Babadoost, M.; Bekal, S.; Lambert, K. Red Light-induced systemic disease resistance against root-knot nematode Meloidogyne javanica and Pseudomonas syringae pv. tomato DC 3000. J. Phytopathol. 2008, 56, 708–714. [Google Scholar] [CrossRef]
- Islam, S.Z.; Honda, Y.; Sawa, Y.; Babadoost, M. Characterization of antifungal glycoprotein in red-light irradiated broad bean leaflets. Mycoscience 2002, 43, 471–473. [Google Scholar] [CrossRef]
- Rahman, M.Z.; Khanam, H.; Ueno, M.; Kihara, J.; Honda, Y.; Arase, S. Suppression by red light irradiation of corynespora leaf spot of cucumber caused by Corynespora cassiicola. J. Phytopathol. 2010, 158, 378–381. [Google Scholar] [CrossRef]
- Wang, H.; Jiang, Y.P.; Yu, H.J.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Yu, J.Q. Light quality affects incidence of powdery mildew, expression of defence-related genes and associated metabolism in cucumber plants. Eur. J. Plant Pathol. 2010, 127, 125–135. [Google Scholar] [CrossRef]
- Ahn, S.Y.; Kim, S.A.; Baek, K.H.; Yun, H.K. Inhibiting wildfire and inducing defense-related gene expression by led treatment on Nicotiana benthamiana. J. Plant Pathol. 2013, 95, 477–483. [Google Scholar]
- Imada, K.; Tanaka, S.; Ibaraki, Y.; Yoshimura, K.; Ito, S. Antifungal effect of 405-nm light on Botrytis cinerea. Lett. Appl. Microbiol. 2014, 59, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Kudo, R.; Ishida, Y.; Yamamoto, K. Effects of green light irradiation on induction of disease resistance in plants. Acta Hortic. 2011, 907, 251–254. [Google Scholar] [CrossRef]
- Kudo, R.; Yamamoto, K.; Suekane, A.; Ishida, Y. Development of green light pest control systems in plants. I. Studies on effects of green light irradiation on induction of disease resistance. SRI Res. Rep. 2009, 93, 31–35. [Google Scholar]
- Khanam, N.N.; Ueno, M.; Kihara, J.; Honda, Y.; Arase, S. Suppression of red light-induced resistance in broad beans to Botrytis cinerea by salicylic acid. Physiol. Mol. Plant Pathol. 2005, 66, 20–29. [Google Scholar] [CrossRef]
- Mutar, S.S.; Fattah, F.A. Red light-induced systemic resistance to root-knot nematodes in tomato. J. Biol. Agric. Healthc. 2013, 3, 110–114. [Google Scholar]
- Rahman, M.Z.; Honda, Y.; Arase, S. Red-light-induced resistance in broad bean (Vicia faba L.) to leaf spot disease caused by Alternaria tenuissima. J. Phytopathol. Phytopathol. Z. 2003, 151, 86–91. [Google Scholar] [CrossRef]
- Jeandet, P.; Douillt-Breuil, A.C.; Bessis, R.; Debord, S.; Sbaghi, M.; Adrian, M. Phytoalexins from the vitaceae: Biosynthesis, phytoalexin gene expression in transgenic plants, antifungal activity, and metabolism. J. Agric. Food Chem. 2002, 50, 2731–2741. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.B.; Yu, K.W.; Hahn, E.J.; Paek, K.Y. Methyl jasmonate and salicylic acid elicitation induces ginsenosides accumulation, enzymatic and non-enzymatic antioxidant in suspension culture Panax ginseng roots in bioreactors. Plant Cell Rep. 2006, 25, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Genoud, T.; Buchala, A.J.; Chua, N.H.; Metraux, J.P. Phytochrome signalling modulates the SA-perceptive pathway in Arabidopsis. Plant J. 2002, 31, 87–95. [Google Scholar] [CrossRef] [PubMed]
- de Wit, M.; Spoel, S.H.; Sanchez-Perez, G.F.; Gommers, C.M.M.; Pieterse, C.M.J.; Voesenek, L.A.C.J.; Pierik, R. Perception of low red: Far-red ratio compromises both salicylic acid- and jasmonic acid-dependent pathogen defences in Arabidopsis. Plant J. 2013, 75, 90–103. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.E.; Tao, Y.; Chory, J.; Ballare, C.L. Ecological modulation of plant defense via phytochrome control of jasmonate sensitivity. Proc. Natl. Acad. Sci. USA 2009, 106, 4935–4940. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Zheng, J.M.; Little, C.; Khosla, S. LED inter-lighting in year-round greenhouse mini-cucumber production. Acta Hortic. 2012, 956, 335–340. [Google Scholar] [CrossRef]
- Li, H.-M.; Lu, X.-M.; Gao, Q.-H. Effects of different light qualities on the growth, photosynthetic pigments and stomatal characteristics of okra (Abelmoschus esculentus) seedlings. Acta Pratac. Sin. 2016, 25, 26–70. [Google Scholar]
- Jokinen, K.; Sarakka, L.E.; Nakkila, J. Improving sweet pepper productivity by LED interlighting. Acta Hortic. 2012, 956, 59–66. [Google Scholar] [CrossRef]
- Sabzalian, M.R.; Heydarizadeh, P.; Zahedi, M.; Boroomand, A.; Agharokh, M.; Sahba, M.R.; Schoefs, B. High performance of vegetables, flowers, and medicinal plants in a red-blue LED incubator for indoor plant production. Agron. Sustain. Dev. 2014, 34, 879–886. [Google Scholar] [CrossRef]
- Shimazaki, K.; Doi, M.; Assmann, S.M.; Kinoshita, T. Light regulation of stomatal movement. Annu. Rev. Plant Biol. 2007, 58, 219–247. [Google Scholar] [CrossRef] [PubMed]
- Astolfi, S.; Marianello, C.; Grego, S.; Bellarosa, R. Preliminary investigation of LED lighting as growth light for seedlings from different tree species in growth chambers. Not. Bot. Horti Agrobot. Cluj-Napoca 2012, 40, 31–38. [Google Scholar]
- Nhut, D.T.; Nam, N.B. Light emitting diodes (LEDs): An artificial lighting source for biological studies. In Proceedings of the 3rd International Conference of the Development of BME in Vietnam, Ho Chi Minh, Vietnam, 11–14 January 2010. [Google Scholar]
- Ghate, V.S.; Ng, K.S.; Zhou, W.B.; Yang, H.; Khoo, G.H.; Yoon, W.B.; Yuk, H.G. Antibacterial effect of light emitting diodes of visible wavelengths on selected foodborne pathogens at different illumination temperatures. Int. J. Food Microbiol. 2013, 166, 399–406. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |


{kind=link}
| LED Light | Light Intensity | Crops | Synthesis of Bioactive Compounds and Crop Traits | References |
|---|---|---|---|---|
| Red | 50 μmol m−2 s−1 | Gossypium hirsutum | Sucrose, starch, soluble sugar | [21] |
| 50 μmol m−2 s−1 | Vitis root-stock | Sugar, starch | [27] | |
| 80 μmol m−2 s−1 | Brassica campestris L. | Starch | [25] | |
| 500 μmol m−2 s−1 | Glycine, Sorghum | Starch | [20] | |
| 30 μmol m−2 s−1 | Betula pendula Roth | Starch | [22] | |
| 50–80 μmol m−2 s−1 | Vitis vinifera | Stilbene | [27,37] | |
| 50 μmol m−2 s−1 | Malus domestica Borkh | Anthocyanin | [30] | |
| 500 μmol m−2 s−1 | Triticum aestivum L. | Lignin | [34] | |
| 128 μmol m−2 s−1 | Pisum sativum | β-Carotene | [38] | |
| 50 μmol m−2 s−1 | B. oleracea var. italic | Delayed senescence | [39] | |
| Blue | 100–200 μmol m−2 s−1 | Lactuca sativa | Phenolic content, Vit-C, tocopherol, carotenoid | [24,32,36,40] |
| 50 μmol m−2 s−1 | Vitis root-stock | Sugar, starch | [27,41] | |
| 80 μmol m−2 s−1 | Brassica campestris L. | Vit. C | [25] | |
| >20–40 μmol m−2 s−1 | Fragaria×ananassa | Organic acids, anthocyanin, ripening | [31,42] | |
| 50–80 μmol m−2 s−1 | B. rapa, B. oleracea var. capitata | Vit. C, polyphenolic content | [25,43,44] | |
| 85–150 μmol m−2 s−1 | Solanum lycopersicum | Proline, Reactive Oxygen Species, scavenger activities, polyphenolic compounds, γ-aminobutyric acid, shelf-life | [23,45] | |
| Blue | 40 μmol m−2 s−1 | Myrica rubra Sieb. and Zucc. | Anthocyanin | [46] |
| 40 μmol m−2 s−1 | Prunus persica | Ripening | [47] | |
| 40 μmol m−2 s−1 | Citrus reticulate | Reduced postharvest decay | [48] | |
| 40–630 μmol m−2 s−1 | Citrus hybrid | Reduced pathogen infection | [49,50] | |
| - | Panax ginseng | Ginsenosides | [51] | |
| 60 μmol m−2 s−1 | Taxus wallichina Zucc | Paclitaxel | [52] | |
| 80 μmol m−2 s−1 | Vitis vinifera | Trans-resveratrol | [53] | |
| Green | ~200 μmol m−2 s−1 | Lactuca sativa, Lens culinaris, Triticum aestivum L., B. oleracea var. capitata, Fragaria×ananassa | Phenolic content, Vit-C, α-tocopherol, anthocyanin | [32,36,40,43,54,55] |
| Yellow | ~100 μmol m−2 s−1 | Raphanus sativus, Malus sp., S. lycopersicum, C. annuum | Vit-C, α-tocopherol, γ-tocopherol, lutein | [55,56] |
| Red+Blue | 70 μmol m−2 s−1 | Doritaenopsis hort | Carotenoids, starch, sucrose, glucose, fructose | [28] |
| >20 μmol m−2 s−1 | Fragaria×ananassa | Organic acids | [31] | |
| 90 μmol m−2 s−1 | Lactuca sativa | Anthocyanin | [26] | |
| - | B. rapa, B. alboglabra | Polyphenol, flavonoids, glucosinolates | [29] | |
| Red + Blue + White | 210 μmol m−2 s−1 | Lactuca sativa | Soluble sugar, nitrate contents | [35] |
| Red + far - red | 50–200 μmol m−2 s−1 | Lactuca sativa, Petunia | Phenolic content, volatile compounds | [36,57] |
| LED Light | Light Intensity | Crops | Effect on Disease | References |
|---|---|---|---|---|
| Red | 261–550 μW/cm2 | Vicia faba | Induces resistance against B. cinerea, Alternaria tenuissima | [64] |
| 250–287 μW/cm2 | Rice sl mutants cultivar (Sekiguchi-asahi and Sekiguchi-himenomochi) | Induced resistance against Magnaporthe grisea | [65] | |
| 287 μW/cm2 | Arabidopsis | Induced resistance against M. javanica, P. syringae pv. tomato DC 3000 | [66] | |
| 287 μW/cm2 | Piper nigrum, Cucurbita, Solanum lycopersicum | Induced resistance against P. capsici | [67] | |
| 137 μW/cm2; 350 μmol m−2 s−1 | Cucumis sativus | Induced resistance against C. cassiicola and S. fuliginea | [68,69] | |
| 80 μmol m−2 s−1 | Vitis vinifera | Induced resistance against B. cinerea | [37] | |
| Nicotiana benthamiana | Induced resistance against P. syringae pv. tabaci | [70] | ||
| Blue | 200 μmol m−2 s−1 | Lactuca sativa | Induced resistance against grey mold by B. cinerea | [40] |
| 50–150 μmol m−2 s−1 | Solanum lycopersicum | Induced resistance against gray mold disease by B. cinerea | [23,71] | |
| 150 μmol m−2 s−1 | Suppression of sporulation of A. cichorii, P. pannosa | [61,62] | ||
| 3.4 μW/cm2 | Reduced spore germination of A. niger | [63] | ||
| Nicotiana benthamiana | Induced resistance against P. syringae pv. tabaci | [70] | ||
| Green | 80 μmol m−2 s−1 | Fragaria×ananassa | Glomerella cingulate | [72] |
| Cucumis sativus | C. orbiculare, B. cinerea | [73] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hasan, M.M.; Bashir, T.; Ghosh, R.; Lee, S.K.; Bae, H. An Overview of LEDs’ Effects on the Production of Bioactive Compounds and Crop Quality. Molecules 2017, 22, 1420. https://doi.org/10.3390/molecules22091420
Hasan MM, Bashir T, Ghosh R, Lee SK, Bae H. An Overview of LEDs’ Effects on the Production of Bioactive Compounds and Crop Quality. Molecules. 2017; 22(9):1420. https://doi.org/10.3390/molecules22091420
Chicago/Turabian StyleHasan, Md. Mohidul, Tufail Bashir, Ritesh Ghosh, Sun Keun Lee, and Hanhong Bae. 2017. "An Overview of LEDs’ Effects on the Production of Bioactive Compounds and Crop Quality" Molecules 22, no. 9: 1420. https://doi.org/10.3390/molecules22091420
APA StyleHasan, M. M., Bashir, T., Ghosh, R., Lee, S. K., & Bae, H. (2017). An Overview of LEDs’ Effects on the Production of Bioactive Compounds and Crop Quality. Molecules, 22(9), 1420. https://doi.org/10.3390/molecules22091420