Isolation, Characterization and Antitumor Effect on DU145 Cells of a Main Polysaccharide in Pollen of Chinese Wolfberry

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Extraction, Isolation and Purification of WPPs
2.2. Further Purification by Sephadex G-100 Column
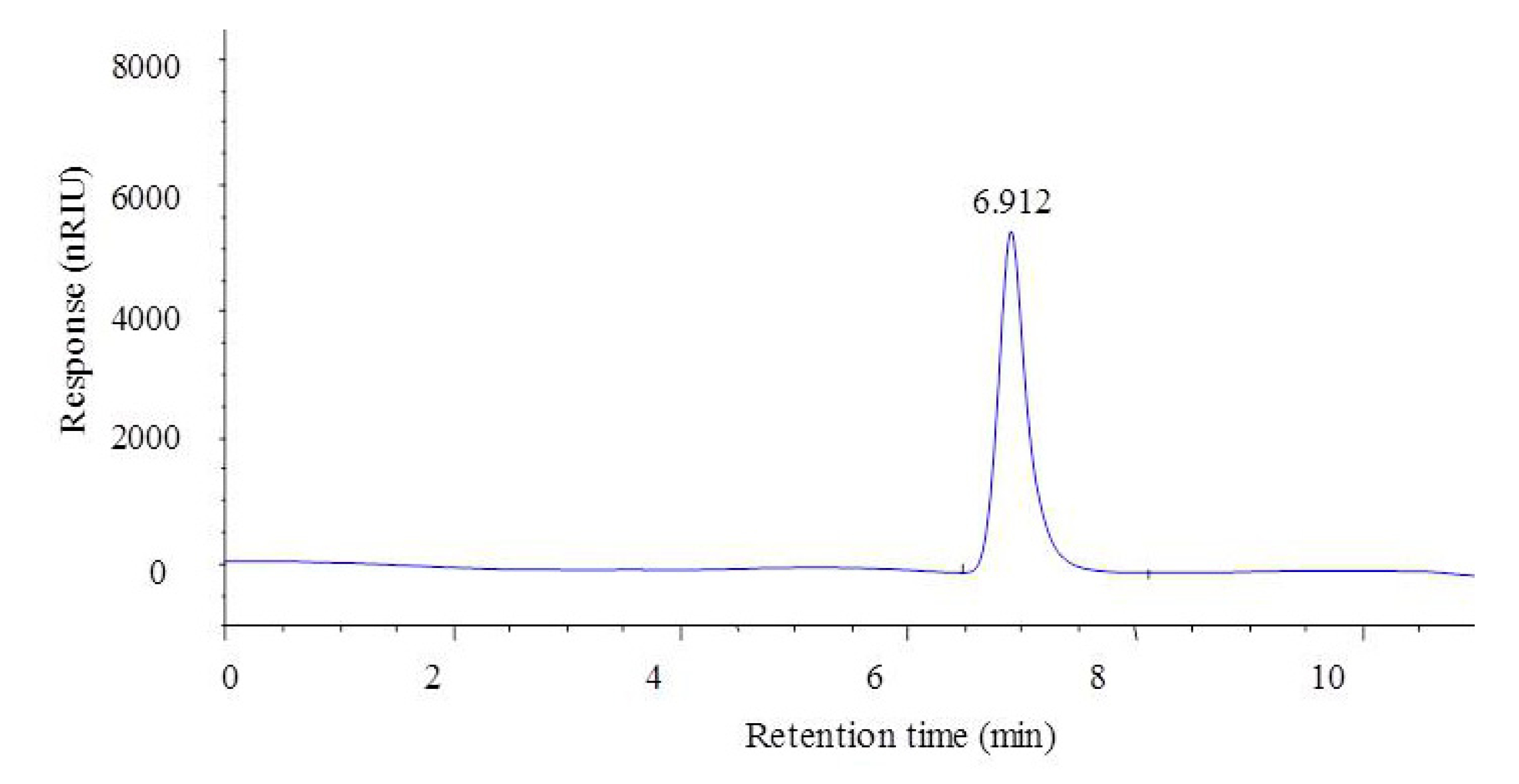
2.3. Analysis of Molecular Weight and Monosaccharide Composition of CF1 of WPPs
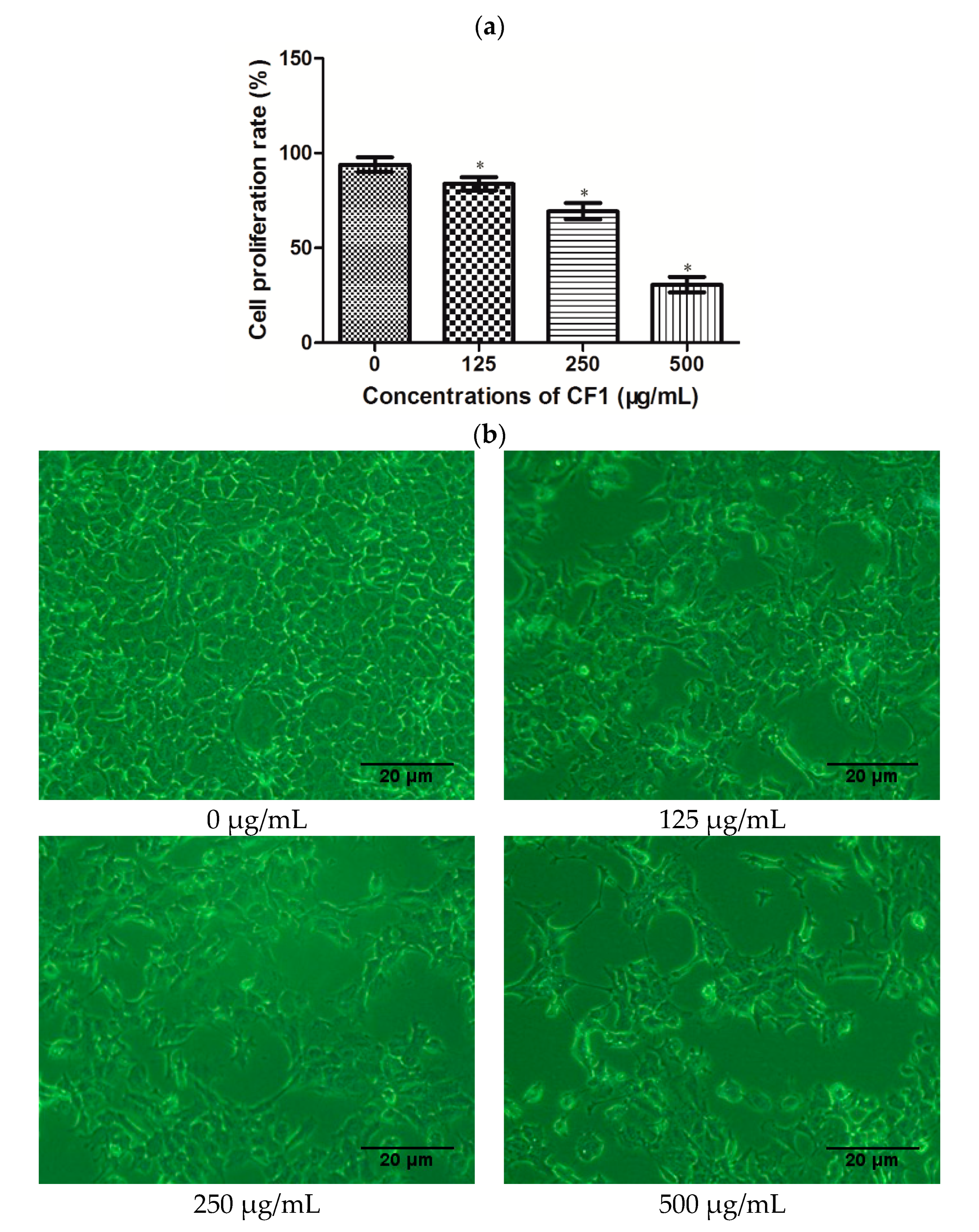
2.4. Effects of CF1 of WPPs on the Proliferation of DU145 Cells
2.5. Effects of the CF1 of WPPs on the Apoptosis Induction of DU145 Cells
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Extraction of Crude WPPs
4.3. Purification of Crude WPPs by DEAE Cellulose-52 Column
4.4. Further Purification by Sephadex G-100 Column
4.5. Analysis of Monosaccharide Composition of CF1 of WPPs
4.6. Determination of the MolecularWeight of CF1 of WPPs
4.7. Antitumor Activities of the CF1 of WPPs In Vitro
4.7.1. Cell Culture
4.7.2. Cell Proliferation Assay
4.7.3. Tunel Assay
4.7.4. Flow Cytometry Assay
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Toren, P.; Zoubeidi, A. Targeting the PI3K/Akt pathway in prostate cancer: Challenges and opportunities (review). Int. J. Oncol. 2014, 45, 1793–1801. [Google Scholar] [CrossRef] [PubMed]
- Žilić, S.; Vančetović, J.; Janković, M.; Maksimović, V. Chemical composition, bioactive compounds, antioxidant capacity and stability of floral maize (Zea mays L.) pollen. J. Funct. Foods. 2014, 10, 65–74. [Google Scholar] [CrossRef]
- Habib, F.K.; Ross, M.; Buck, A.C.; Ebeling, L.; Lewenstein, A. In vitro evaluation of the pollen extract cernitin T60, in the regulation of prostrate cell growth. Br. J. Urol. 1990, 66, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.F.; Tang, L.P.; He, R.R.; Xu, Z.; Xiang, F.J.; Su, W.W.; Hiroshi, K. Anthocyanins extract from bilberry enhances the therapeutic effect of pollen of Brassica napus L. on stress-provoked benign prostatic hyperplasia in restrained mice. J. Funct. Foods 2013, 5, 1357–1365. [Google Scholar] [CrossRef]
- Monden, K.; Tsugawa, M.; Ninomiya, Y.; Ando, E.; Kumon, H.A. Japanese version of the National Institutes of Health Chronic Prostatitis Symptom Index (NIH-CPSI, Okayama version) and the clinical evaluation of cernitin pollen extract for chronic non-bacterial prostatitis. Nihon Hinyokika Gakkai Zasshi 2002, 93, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Brecker, L.; Wicklein, D.; Moll, H.; Fuchs, E.C.; Becker, W.M.; Petersen, A. Structural and immunological properties of arabinogalactan polysaccharides from pollen of timothy grass (Phleumpratense L.). Carbohydr. Res. 2005, 340, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Guo, D.; Zhang, J.; Wu, M. Characterization and antitumor activity of pollen polysaccharide. Int. Immunopharmacol. 2007, 7, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Diao, Q.; Zhang, Z.; Liu, Y.; Gao, Q.; Zhou, Y.; Li, S. Antitumor activity of bee pollen polysaccharides from Rosa rugosa. Mol. Med. Rep. 2013, 7, 1555–1558. [Google Scholar] [CrossRef] [PubMed]
- Tyliszczak, B.; Drabczyk, A.; Kudłacik-kramarczyk, S.; Grabowska, B.; Kędzierska, M. Physicochemical properties and cytotoxicity of hydrogels based on Beetosan® containing sage and bee pollen. Acta Biochim. Pol. 2017, 64, 709–712. [Google Scholar] [CrossRef] [PubMed]
- Yue, G.; Li, X.; Sun, M.; Su, F. Immunomodulatory effects of sulfated polysaccharides of pine pollen on mouse macrophages. Int. J. Biol. Macromol. 2016, 91, 846–855. [Google Scholar] [CrossRef]
- Luo, Q.; Li, J.; Cui, X.; Yan, J.; Zhao, Q.; Xiang, C. The effect of Lycium barbarum, polysaccharides on the male rats reproductive system and spermatogenic cell apoptosis exposed to low-dose ionizing irradiation. J. Ethnopharmacol. 2014, 154, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Gong, G.P.; Fan, J.B.; Sun, Y.J.; Wu, Y.M.; Liu, Y.; Sun, W.; Zhang, Y.; Wang, Z.F. Isolation, structural characterization, and antioxidativity of polysaccharide LBLP5-A from Lycium barbarum, leaves. Process. Biochem. 2015, 51, 314–324. [Google Scholar] [CrossRef]
- Mao, F.; Xiao, B.; Jiang, Z.; Zhao, J.; Huang, X.; Guo, J. Anticancer effect of Lycium barbarum, polysaccharides on colon cancer cells involves G0/G1 phase arrest. Med. Oncol. 2011, 28, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Liang, L.; Wang, Y.; Diao, J.; Zhao, C.; Chen, G.; He, Y.; Luo, C.; Wu, X.; Zhang, Y. Synergistic immunotherapeutic effects of Lycium barbarum polysaccharide and interferon-α2b on the murine Renca renal cell carcinoma cell line in vitro and in vivo. Mol. Med. Rep. 2015, 12, 6727–6737. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.; Lv, X.; Tao, W.; Ma, Q.; Teng, A.; Zhang, Y.; Zhang, M. Composition of Lycium barbarum polysaccharides and their apoptosis-inducing effect on human hepatoma SMMC-7721 cells. Food Nutr. Res. 2015, 59, 28696. [Google Scholar] [CrossRef]
- Miao, Y.; Xiao, B.; Jiang, Z.; Guo, Y.; Mao, F.; Zhao, J.; Huang, X.; Guo, J. Growth inhibition and cell-cycle arrest of human gastric cancer cells by Lycium barbarum polysaccharide. Med. Oncol. 2010, 27, 785–790. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wei, K.; Yang, S.; Yang, Y.; Zhang, Y.; Zhu, F.; Wang, D.; Zhu, R. Immunomodulatory effects of Taishan Pinus massoniana pollen polysaccharide and propolis on immunosuppressed chickens. Microb. Pathog. 2015, 78, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Gong, H.; Yang, S.; Yang, L.; Fan, Y.; Zhou, Y. Pectic bee pollen polysaccharide from rosarugosa alleviates diet-induced hepatic steatosis and insulin resistance via induction of AMPK/mTOR-mediated autophagy. Molecules 2017, 22, E699. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Liu, Y.; Yuan, G.; Guan, M. The contribution of side chains to antitumor activity of a polysaccharide from Codonopsispilosula. Int. J. Biol. Macromol. 2012, 50, 891–894. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.L.; Yuan, H.H.; Wang, C.Z.; Fan, W.; Lan, M.B. Polysaccharides from Cymbopogoncitratus with antitumor and immunomodulatory activity. Pharm. Biol. 2015, 53, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Liu, N.; Wu, Z.; Feng, Y.; Meng, X. Anti-tumor activity of a polysaccharide from blueberry. Molecules 2015, 20, 3841–3853. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Lin, M.; Luo, A.; Chun, Z.; Luo, A. Characterization and antitumor activity of a polysaccharide from sarcodiaceylonensis. Molecules 2014, 19, 10863–10876. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, S.; Sun, L.; Ji, L.; Zhu, J.; Fan, Y.; Tai, G.; Zhou, Y. Further analysis of the structure and immunological activity of an RG-I type pectin from Panax ginseng. Carbohydr. Polym. 2012, 89, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Luo, X.; Xu, X.; Wei, W.; Yu, M.; Jiang, N.; Ye, L.; Yang, Z.; Fei, X. Antioxidant and immunomodulatory activities of a polysaccharide from Flammulinavelutipes. J. Tradit. Chin. Med. 2014, 34, 733–740. [Google Scholar] [CrossRef]
- Suabjakyong, P.; Nishimura, K.; Toida, T.; Van Griensven, L.J. Structural characterization and immunomodulatory effects of polysaccharides from Phellinuslinteus and Phellinusigniarius on the IL-6/IL-10 cytokine balance of the mouse macrophage cell lines (RAW 264.7). Food Funct. 2015, 6, 2834–2844. [Google Scholar] [CrossRef] [PubMed]
- Kouakou, K.; Schepetkin, I.A.; Yapi, A.; Kirpotina, L.N.; Jutila, M.A.; Quinn, M.T. Immunomodulatory activity of polysaccharides isolated from Alchorneacordifolia. J. Ethnopharmacol. 2013, 146, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Cör, D.; Knez, Ž.; Knez Hrnčič, M. Antitumour, antimicrobial, antioxidant and antiacetylcholinesterase effect of Ganoderma Lucidum terpenoids and polysaccharides: A review. Molecules 2018, 23, 649. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.Y.; Dong, F.; Liu, X.; Qian, L.; Yang, Y.; Fei, L.; Chen, L.; Wang, T.; Wang, Z.; Zhang, Y. Effects of extraction methods on the yield, chemical structure and anti-tumor activity of polysaccharides from Cordyceps gunnii, mycelia. Carbohydr. Polym. 2016, 140, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jiao, L.L.; Zhang, X.; Tian, W.M.; Chen, S.; Zhang, L.P. Anti-tumor and immunomodulating activities of proteoglycans from mycelium of Phellinusnigricans and culture medium. Int. Immunopharmacol. 2008, 8, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Bao, H.H.; Choi, W.S.; You, S. Effect of sulfated modification on the molecular characteristics and biological activities of polysaccharides from Hypsizigusmarmoreus. Biosci. Biotechnol. Biochem. 2010, 74, 1408–1414. [Google Scholar] [CrossRef] [PubMed]
- Lombard, Y.; Giaimis, J.; Makaya-Kumba, M.; Fonteneau, P.; Poindron, P. A new method for studying the binding and ingestion of zymosan particles by macrophages. J. Immunol. Methods 1994, 174, 155–165. [Google Scholar] [CrossRef]
- Zheng, Y.; Hao, M.; Nan, H.; Jeff, I.; Zhou, Y.; Gao, Y. Relationship of chemical composition and cytotoxicity of water-soluble polysaccharides from lentinus edodes fruiting bodies. Pak. J. Pharm. Sci. 2015, 28, 1069–1074. [Google Scholar] [PubMed]
- Zong, A.; Cao, H.; Wang, F. Anticancer polysaccharides from natural resources: A review of recent research. Carbohydr. Polym. 2012, 90, 1395–1410. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhu, J.Y.; Lei, Z.M.; Wan, L.J.; Zhu, X.W.; Ye, F.; Tong, Y.Y. Anti-proliferative effects of paeonol on human prostate cancer cell lines DU145 and PC3. J. Physiol. Biochem. 2017, 73, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Choo, G.S.; Lee, H.N.; Shin, S.A.; Kim, H.J.; Jung, J.Y. Anticancer effect of fucoidan on DU-145 prostate cancer cells through inhibition of PI3K/Akt and MAPK pathway expression. Mar. Drugs 2016, 14, E126. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Li, Z.N.; Yan, J.; Zhu, F.; Xu, R.J.; Cai, Y.Z. Lycium barbarum polysaccharides induce apoptosis in human prostate cancer cells and inhibits prostate cancer growth in a xenograft mouse model of human prostate cancer. J. Med. Food 2009, 12, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Chen, H.; Huang, J.; Li, Z.; Zhu, C.; Zhang, S. Effect of Lycium barbarum polysaccharide on human hepatoma QGY7703 cells: Inhibition of proliferation and induction of apoptosis. Life Sci. 2005, 76, 2115–2124. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhao, X.Z.; Qi, Q.; Tao, L.; Zhao, Q.; Mu, R.; Gu, H.Y.; Wang, M.; Feng, X.; Guo, Q.L. Macranthoside B, a hederageninsaponin extracted from Loniceramacranthoides and its anti-tumor activities in vitro and in vivo. Food Chem. Toxicol. 2009, 47, 1716–1721. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.Y.; Li, L.Z.; Xue, K.C.; Hu, D.D.; Gao, Y.J. Antitumor activity of fucoidan in anaplastic thyroid cancer via apoptosis and anti-angiogenesis. Mol. Med. Rep. 2017, 15, 2620–2624. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.S.; Kuo, H.P.; Chang, K.L.; Kong, Z.L. Apoptosis of hepatocellular carcinoma cells induced by nanoencapsulated polysaccharides extracted from antrodia camphorata. PLoS ONE 2015, 10, e0136782. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Sun, K.; Zhu, Q.; Song, T.; Liu, Y. Ginseng polysaccharide serves as a potential radiosensitizer through inducing apoptosis and autophagy in the treatment of osteosarcoma. Kaohsiung J. Med. Sci. 2017, 33, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.H.; Liu, Y.; Wu, X.L.; Liu, L.Z.; Fu, W.; Song, D.D. Isolation, purification, characterization and immunostimulatory activity of polysaccharides derived from American ginseng. Carbohydr. Polym. 2017, 156, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Xie, Y.; Wang, W.; Yan, Y.; Ye, H.; Jabbar, S.; Zeng, X. Extraction optimization, characterization and antioxidant activity in vitro of polysaccharides from mulberry (Morus alba L.) leaves. Carbohydr. Polym. 2015, 128, 52–62. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compound CF1 is available from the authors. |







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, F.; Ran, L.; Mi, J.; Yan, Y.; Lu, L.; Jin, B.; Li, X.; Cao, Y. Isolation, Characterization and Antitumor Effect on DU145 Cells of a Main Polysaccharide in Pollen of Chinese Wolfberry. Molecules 2018, 23, 2430. https://doi.org/10.3390/molecules23102430
Chen F, Ran L, Mi J, Yan Y, Lu L, Jin B, Li X, Cao Y. Isolation, Characterization and Antitumor Effect on DU145 Cells of a Main Polysaccharide in Pollen of Chinese Wolfberry. Molecules. 2018; 23(10):2430. https://doi.org/10.3390/molecules23102430
Chicago/Turabian StyleChen, Fei, Linwu Ran, Jia Mi, Yamei Yan, Lu Lu, Bo Jin, Xiaoying Li, and Youlong Cao. 2018. "Isolation, Characterization and Antitumor Effect on DU145 Cells of a Main Polysaccharide in Pollen of Chinese Wolfberry" Molecules 23, no. 10: 2430. https://doi.org/10.3390/molecules23102430
APA StyleChen, F., Ran, L., Mi, J., Yan, Y., Lu, L., Jin, B., Li, X., & Cao, Y. (2018). Isolation, Characterization and Antitumor Effect on DU145 Cells of a Main Polysaccharide in Pollen of Chinese Wolfberry. Molecules, 23(10), 2430. https://doi.org/10.3390/molecules23102430