Heat Shock Proteins as Immunomodulants


Abstract
:1. Introduction
1.1. Major Heat Shock Protein Families and Some of Their Roles in Immunomodulation
1.2. Heat Shock Proteins as ‘Chaperokines’
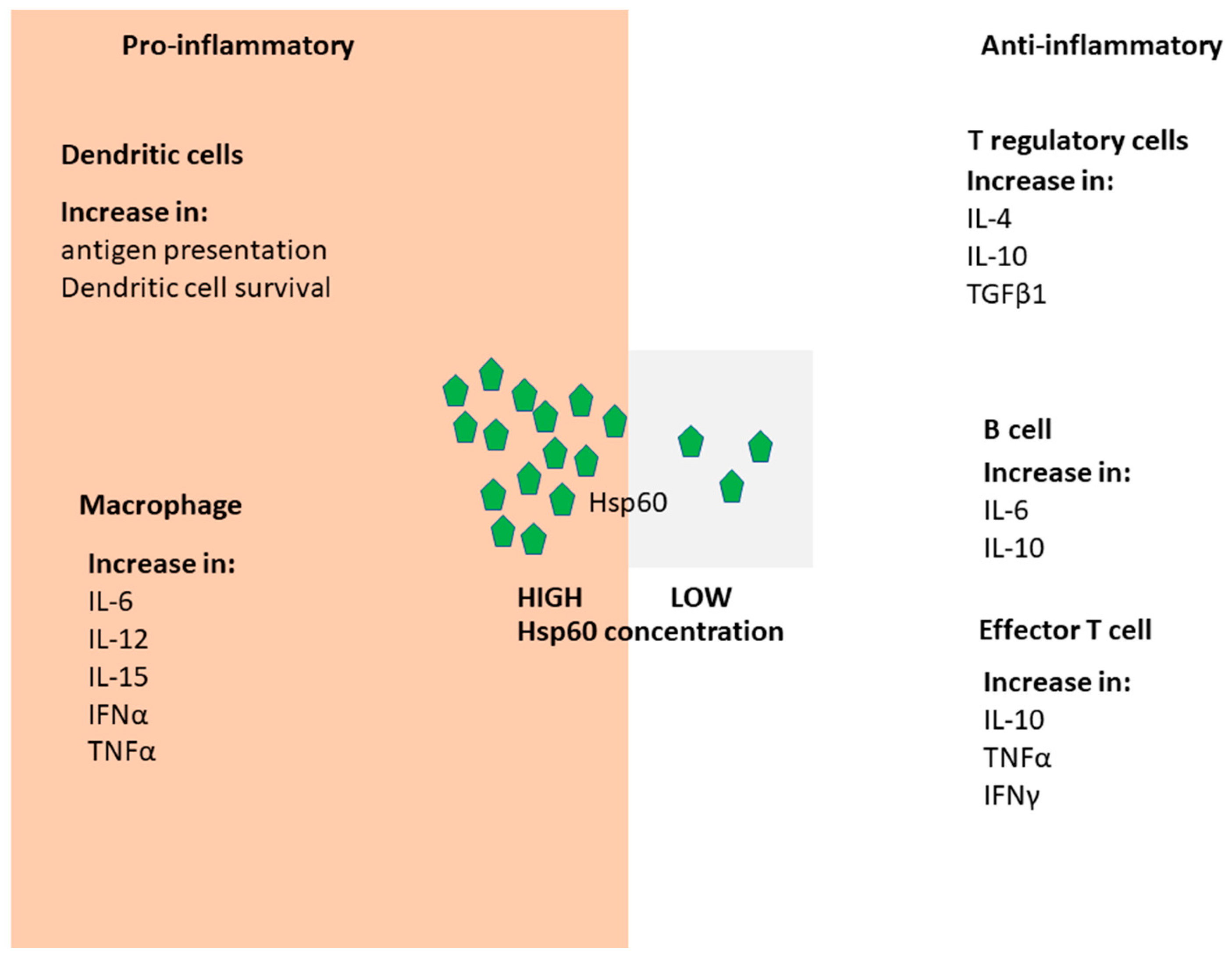
1.3. Hsp60
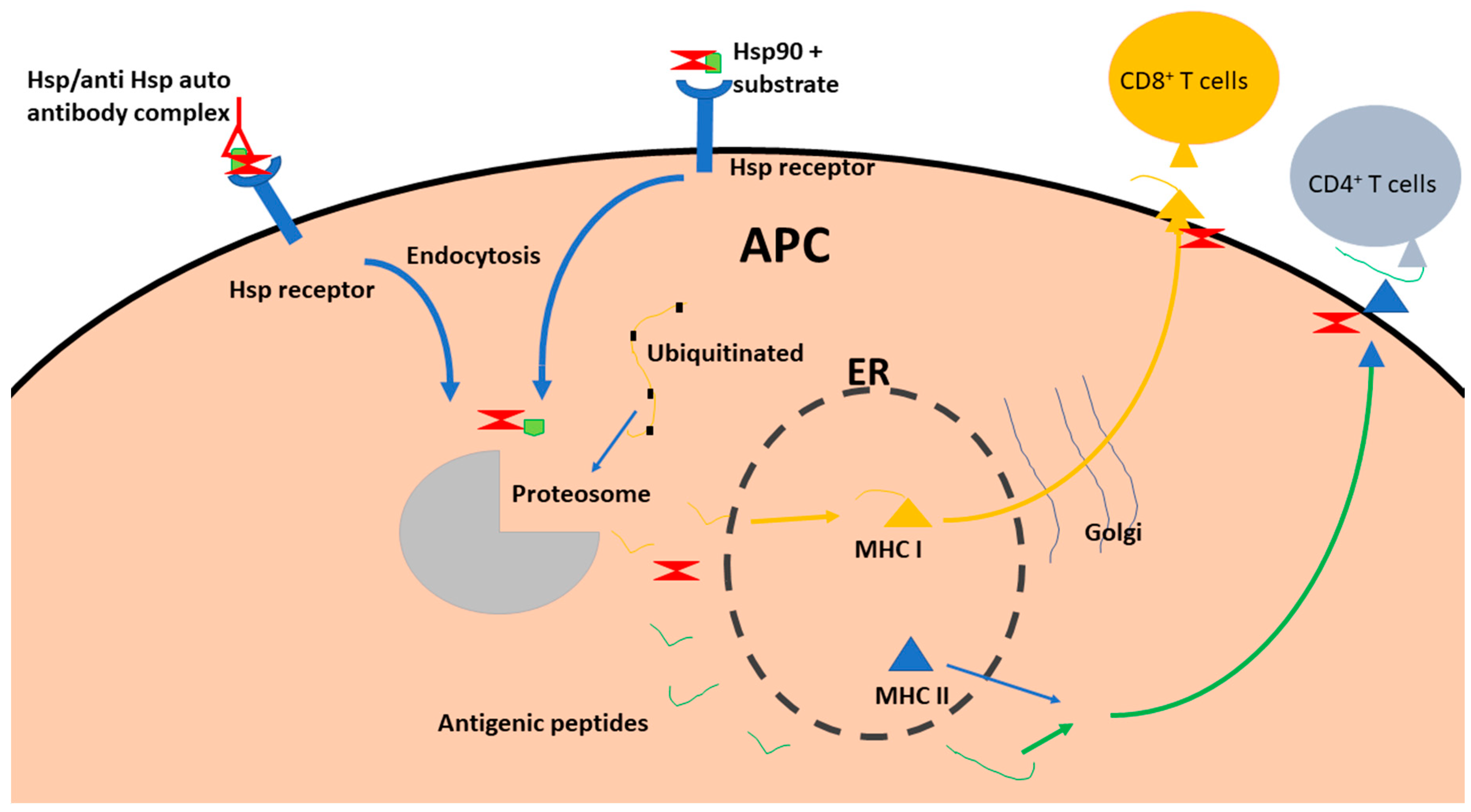
1.4. Hsp90
1.5. Hsp40
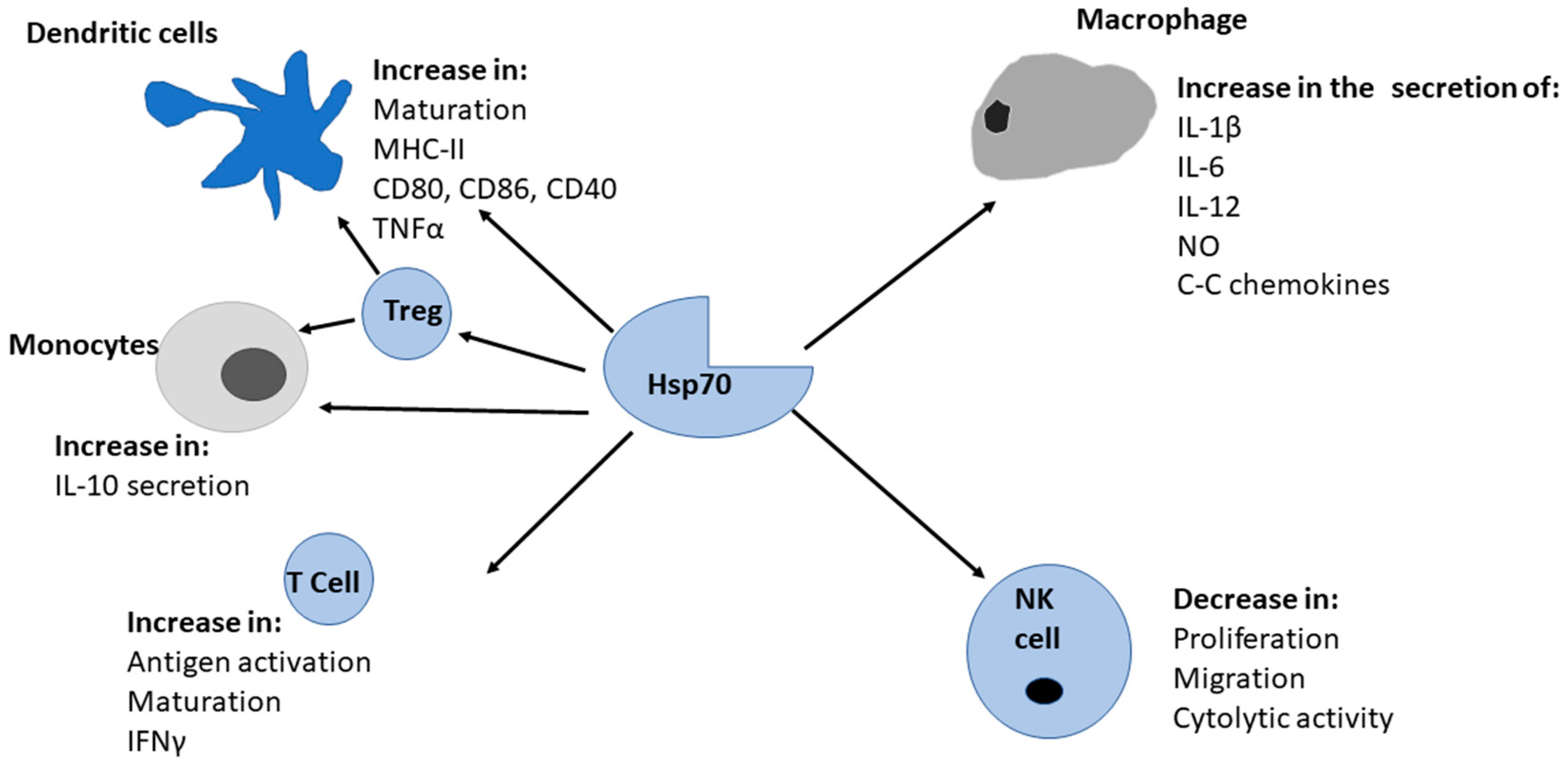
1.6. Hsp70
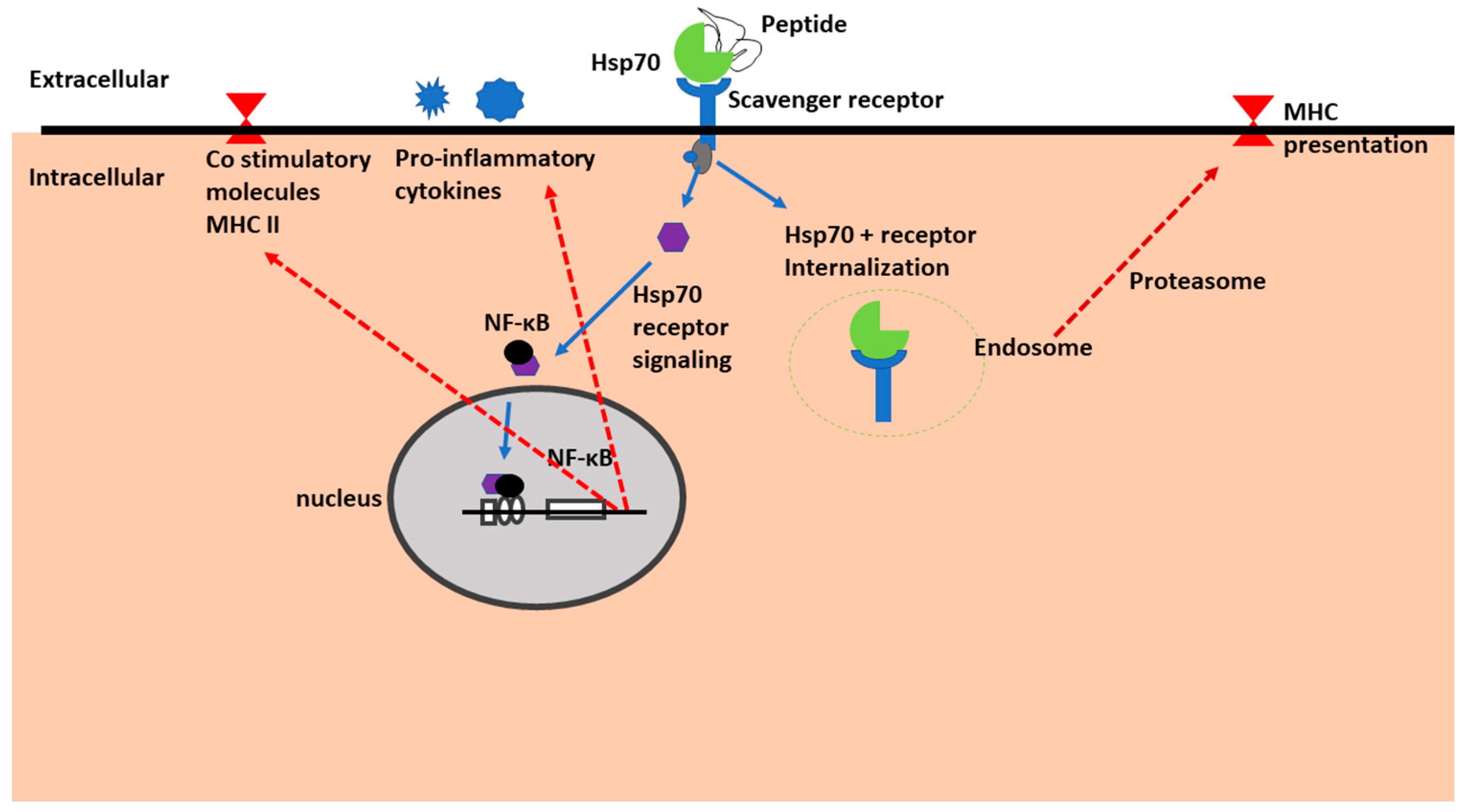
1.7. Hsp70 as Receptors
1.8. Small Hsps
1.9. Contaminating Constituents that Confound Immunomodulation Studies
2. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bukau, B.; Weissman, J.; Horwich, A. Molecular chaperones and protein quality control. Cell 2016, 125, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Kriegenburg, F.; Ellgaard, L.; Hartmann-Petersen, R. Molecular chaperones in targeting misfolded proteins for ubiquitin-dependent degradation. FEBS J. 2012, 279, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Calderwood, S.K.; Gong, J.; Murshid, A. Extracellular HSPs: The complicated roles of extracellular HSPs in immunity. Front. Immunol. 2016, 7, 159. [Google Scholar] [CrossRef] [PubMed]
- Multhoff, G.; Hightower, L.E. Distinguishing integral and receptor-bound heat shock protein 70 (Hsp70) on the cell surface by Hsp70-specific antibodies. Cell Stress Chaperones 2011, 16, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Pockley, A.G.; Multhoff, G. Cell stress proteins in extracellular fluids: Friend or foe? Novartis Found. Symp. 2008, 291, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Cabral, F.J.; Vianna, L.G.; Medeiros, M.M.; Carlos, B.C.; Martha, R.D.; Silva, N.M.; Da Silva, L.H.P.; Stabeli, R.G.; Wunderlich, G. Immunoproteomics of Plasmodium falciparum-infected red blood cell membrane fractions. Mem. Inst. Oswaldo Cruz 2017, 112, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Dreiza, C.M.; Komalavilas, P.; Furnish, E.J.; Flynn, C.R.; Sheller, M.R.; Smoke, C.C.; Lopes, L.B.; Brophy, C.M. The small heat shock protein, HSPB6, in muscle function and disease. Cell Stress Chaperones 2010, 15, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Murshid, A.; Gong, J.; Calderwood, S.K. The role of heat shock proteins in antigen cross presentation. Front. Immunol. 2012, 3, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rappa, F.; Farina, F.; Zummo, G.; David, S.; Campanella, C.; Carini, F.; Tomasello, G.; Damiani, P.; Cappello, F.; de Macario, E.C.; et al. HSP-molecular chaperones in cancer biogenesis and tumor therapy: An overview. Anticancer Res. 2012, 32, 5139–5150. [Google Scholar] [PubMed]
- Khalil, A.A.; Kabapy, N.F.; Deraz, S.F.; Smith, C. Heat shock proteins in oncology: Diagnostic biomarkers or therapeutic targets? Biochim. Biophys. Acta 2011, 1816, 89–104. [Google Scholar] [CrossRef] [PubMed]
- Caruso Bavisotto, C.; Cappello, F.; Macario, A.J.; Conway de Macario, E.; Logozzi, M.; Fais, S.; Campanella, C. Exosomal HSP60: A potentially useful biomarker for diagnosis, assessing prognosis, and monitoring response to treatment. Expert Rev. Mol. Diagn. 2017, 17, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Li, X.S.; Xu, Q.; Fu, X.Y.; Luo, W.S. Heat shock protein 60 overexpression is associated with the progression and prognosis in gastric cancer. PLoS ONE 2014, 9, 107507. [Google Scholar] [CrossRef] [PubMed]
- Bakthisaran, R.; Tangirala, R.; Rao, C.M. Small heat shock proteins: Role in cellular functions and pathology. Biochim. Biophys. Acta 2015, 1854, 291–319. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Cui, J.; Zhang, X.; Gao, S.; Ma, F.; Yao, H.; Sun, X.; He, Y.; Yin, Y.; Xu, W. Pneumococcal DnaJ modulates dendritic cell-mediated Th1 and Th17 immune responses through toll-like receptor 4 signaling pathway. Immunobiology 2017, 222, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Ma, C.; Ye, G.; Shi, Y.; Xu, W.; Zhong, L.; Wang, J.; Yin, Y.; Zhang, X.; Wang, H. DnaJ (hsp40) of Streptococcus pneumoniae is involved in bacterial virulence and elicits a strong natural immune reaction via PI3K/JNK. Mol. Immunol. 2017, 83, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Juwono, J.; Martinus, R.D. Does Hsp60 provide a link between mitochondrial stress and inflammation in diabetes mellitus? J. Diabetes Res. 2016, 2016, 8017571. [Google Scholar] [CrossRef] [PubMed]
- Borges, T.J.; Wieten, L.; Van Herwijnen, M.J.; Broere, F.; Van Der Zee, R.; Bonorino, C.; Van Eden, W. The anti-inflammatory mechanisms of Hsp70. Front. Immunol. 2012, 3, 95. [Google Scholar] [CrossRef] [PubMed]
- Gross, C.; Schmidt-Wolf, I.G.; Nagaraj, S.; Gastpar, R.; Ellwart, J.; Kunz-Schughart, L.A.; Multhoff, G. Heat shock protein 70-reactivity is associated with increased cell surface density of CD94/CD56 on primary natural killer cells. Cell Stress Chaperones 2003, 8, 348–360. [Google Scholar] [CrossRef]
- Mbofung, R.M.; McKenzie, J.A.; Malu, S.; Zhang, M.; Peng, W.; Liu, C.; Kuiatse, I.; Tieu, T.; Williams, L.; Devi, S.; et al. Hsp90 inhibition enhances cancer immunotherapy by upregulating interferon response genes. Nat. Commun. 2017, 8, 451. [Google Scholar] [CrossRef] [PubMed]
- Asea, A.; Kabingu, E.; Stevenson, M.A.; Calderwood, S.K. Hsp70 peptide-bearing and peptide-negative preparations act as chaperokines. Cell Stress Chaperones 2000, 5, 425–431. [Google Scholar] [CrossRef]
- Hulina, A.; Rajković, M.G.; Despot, D.J.; Jelić, D.; Dojder, A.; Čepelak, I.; Rumora, L. Extracellular Hsp70 induces inflammation and modulates LPS/LTA-stimulated inflammatory response in THP-1 cells. Cell Stress Chaperones 2018, 23, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Tsan, M.F.; Gao, B. Cytokine function of heat shock proteins. Am. J. Physiol. Cell Physiol. 2004, 286, 739–744. [Google Scholar] [CrossRef] [PubMed]
- Svensson, P.A.; Asea, A.; Englund, M.C.; Bausero, M.A.; Jernas, M.; Wiklund, O.; Ohlsson, B.G.; Carlsson, L.M.; Carlsson, B. Major role of Hsp70 as a paracrine inducer of cytokine production in human oxidized LDL treated macrophages. Atherosclerosis 2006, 185, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Ferat-Osorio, E.; Sánchez-Anaya, A.; Gutiérrez-Mendoza, M.; Boscó-Gárate, I.; Wong-Baeza, I.; Pastelin-Palacios, R.; Pedraza-Alva, G.; Bonifaz, L.C.; Cortés-Reynosa, P.; Pérez-Salazar, E.; et al. Heat shock protein 70 down-regulates the production of toll-like receptor-induced pro-inflammatory cytokines by a heat shock factor-1/constitutive heat shock element-binding factor-dependent mechanism. J. Inflamm. 2014, 11, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atre, N.; Thomas, L.; Mistry, R.; Pathak, K.; Chiplunkar, S. Role of nitric oxide in heat shock protein induced apoptosis of γδT cells. Int. J. Cancer 2006, 119, 1368–1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuppner, M.C.; Gastpar, R.; Gelwer, S.; Nössner, E.; Ochmann, O.; Scharner, A.; Issels, R.D. The role of heat shock protein (hsp70) in dendritic cell maturation: Hsp70 induces the maturation of immature dendritic cells but reduces DC differentiation from monocyte precursors. Eur J. Immunol. 2001, 31, 1602–1609. [Google Scholar] [CrossRef] [Green Version]
- Stocki, P.; Dickinson, A.M. The immunosuppressive activity of heat shock protein 70. Autoimmune Dis. 2012, 2012, 617213. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.S. Sequence and structural homology between a mouse T-complex protein TCP-1 and the ‘chaperonin’ family of bacterial (GroEL, 60–65 kDa heat shock antigen) and eukaryotic proteins. Biochem. Int. 1990, 20, 833–841. [Google Scholar] [PubMed]
- Quintana, F.J.; Cohen, I.R. The Hsp60 immune system network. Trends Immunol. 2010, 32, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Calderwood, S.K.; Mambula, S.S.; Gray, P.J. Extracellular heat shock proteins in cell signaling and immunity. Ann. N. Y. Acad. Sci. 2007, 1113, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Vilasi, S.; Bulone, D.; Caruso Bavisotto, C.; Campanella, C.; Marino Gammazza, A.; San Biagio, P.L.; Cappello, F.; Conway de Macario, E.; Macario, A. Chaperonin of group I: Oligomeric spectrum and biochemical and biological implications. Front. Mol. Biosci. 2018, 4, 99. [Google Scholar] [CrossRef] [PubMed]
- Vilasi, S.; Carrotta, R.; Mangione, M.R.; Campanella, C.; Librizzi, F.; Randazzo, L.; Martorana, V.; Gammazza, A.M.; Ortore, M.G.; Vilasi, A.; et al. Human Hsp60 with its mitochondrial import signal occurs in solution as heptamers and tetradecamers remarkably stable over a wide range of concentrations. PLoS ONE 2014, 9, 97657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riffo-Vasquez, Y.; Spina, D.; Page, C.; Tormay, P.; Singh, M.; Henderson, B.; Coates, A. Effect of Mycobacterium tuberculosis chaperonins on bronchial eosinophilia and hyper-responsiveness in a murine model of allergic inflammation. Clin. Exp. Allergy 2004, 34, 712–719. [Google Scholar] [CrossRef] [PubMed]
- Henderson, B.; Pockley, A. Molecular chaperones and protein-folding catalysts as intercellular signaling regulators in immunity and inflammation. J. Leukoc. Biol. 2010, 88, 445–462. [Google Scholar] [CrossRef] [PubMed]
- Corrao, S.; Campanella, C.; Anzalone, R.; Farina, F.; Zummo, G.; de Macario, E.C.; Macario, A.J.; Cappello, F.; La Rocca, G. Human Hsp10 and Early Pregnancy Factor (EPF) and their relationship and involvement in cancer and immunity: Current knowledge and perspectives. Life Sci. 2010, 86, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Rolfe, B.E.; Cavanagh, A.C.; Quinn, K.A.; Morton, H. Identification of two suppressor factors induced by early pregnancy factor. Clin. Exp. Immunol. 1988, 73, 219–225. [Google Scholar] [PubMed]
- Dordea, A.C.; Sweeney, M.; Taggart, J.; Lartey, J.; Wessel, H.; Robson, S.C.; Taggart, M.J. Differential vasodilation of human placental and myometrial arteries related to myofilament Ca2+-desensitization and the expression of Hsp20 but not MYPT1. Mol. Hum. Reprod. 2013, 19, 727–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, A.J.; Paulson, H.L. Polyglutamine neurodegeneration: Protein misfolding revisited. Trends Neurosci. 2008, 31, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Verdegaal, M.E.; Zegveld, S.T.; van Furth, R. Heat shock protein 65 induces CD62e, CD106, and CD54 on cultured human endothelial cells and increases their adhesiveness for monocytes and granulocytes. J. Immunol. 1996, 157, 369–376. [Google Scholar] [PubMed]
- Pei, W.; Tanaka, K.; Huang, S.C.; Xu, L.; Liu, B.; Sinclair, J.; Idol, J.; Varshney, G.K.; Huang, H.; Lin, S.; et al. Extracellular Hsp60 triggers tissue regeneration and wound healing by regulating inflammation and cell proliferation. NPJ Regen. Med. 2016, 1, 16013. [Google Scholar] [CrossRef] [PubMed]
- Gobert, A.P.; Bambou, J.C.; Werts, C.; Balloy, V.; Chignard, M.; Moran, A.P.; Ferrero, R.L. Helicobacter pylori heat shock protein 60 mediates interleukin-6 production by macrophages via a toll-like receptor (TLR)-2-, TLR-4-, and myeloid differentiation factor 88-independent mechanism. J. Biol. Chem. 2004, 279, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Cehovin, A.; Coates, A.R.; Hu, Y.; Riffo-Vasquez, Y.; Tormay, P.; Botanch, C.; Altare, F.; Henderson, B. Comparison of the moonlighting actions of the two highly homologous chaperonin 60 proteins of Mycobacterium tuberculosis. Infect. Immun. 2010, 78, 3196–3206. [Google Scholar] [CrossRef] [PubMed]
- Rha, Y.H.; Taube, C.; Haczku, A.; Joetham, A.; Takeda, K.; Duez, C.; Siegel, M.; Aydintug, M.K.; Born, W.K.; Dakhama, A.; et al. Effect of microbial heat shock proteins on airway inflammation and hyperresponsiveness. J. Immunol. 2002, 169, 5300–5307. [Google Scholar] [CrossRef] [PubMed]
- Hickey, T.B.; Ziltener, H.J.; Speert, D.P.; Stokes, R.W. Mycobacterium tuberculosis employs Cpn60. 2 as an adhesin that binds CD43 on the macrophage surface. Cell Microbiol. 2010, 12, 1634–1647. [Google Scholar] [CrossRef] [PubMed]
- Hjelholt, A.; Carlsen, T.; Deleuran, B.; Jurik, A.G.; Schiøttz-Christensen, B.; Christiansen, G.; Birkelund, S. Increased levels of IgG antibodies against human HSP60 in patients with spondyloarthritis. PLoS ONE 2013, 8, 56210. [Google Scholar] [CrossRef] [PubMed]
- Marino Gammazza, A.M.; Rizzo, M.; Citarrella, R.; Rappa, F.; Campanella, C.; Bucchieri, F.; Patti, A.; Nikolic, D.; Cabibi, D.; Amico, G.; et al. Elevated blood Hsp60, its structural similarities and cross-reactivity with thyroid molecules, and its presence on the plasma membrane of oncocytes point to the chaperonin as an immunopathogenic factor in Hashimoto’s thyroiditis. Cell Stress Chaperones 2013, 19, 343–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marino Gammazza, A.M.; Bucchieri, F.; Grimaldi, L.M.; Benigno, A.; de Macario, E.C.; Macario, A.J.; Zummo, G.; Cappello, F. The molecular anatomy of human Hsp60 and its similarity with that of bacterial orthologs and acetylcholine receptor reveal a potential pathogenetic role of anti-chaperonin immunity in myasthenia gravis. Cell Mol. Neurobiol. 2012, 32, 943–947. [Google Scholar] [CrossRef] [PubMed]
- Cappello, F.; Marino Gammazza, A.; Zummo, L.; Conway de Macario, E.; Macario, A.J. Hsp60 and AChR cross-reactivity in myasthenia gravis: An update. J. Neurol. Sci. 2010, 292, 117–118. [Google Scholar] [CrossRef] [PubMed]
- Campanella, C.; Marino Gammazza, A.; Mularoni, L.; Cappello, F.; Zummo, G.; Di Felice, V. A comparative analysis of the products of GROEL-1 gene from Chlamydia trachomatis serovar D and the HSP60 var1 transcript from Homo sapiens suggests a possible autoimmune response. Int. J. Immunogenet. 2009, 36, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Yokota, S.I.; Hirata, D.; Minota, S.; Higashiyama, T.; Kurimoto, M.; Yanagi, H.; Yura, T.; Kubota, H. Autoantibodies against chaperonin CCT in human sera with rheumatic autoimmune diseases: Comparison with antibodies against other Hsp60 family proteins. Cell Stress Chaperones 2000, 5, 337–346. [Google Scholar] [CrossRef]
- Landstein, D.; Ulmansky, R.; Naparstek, Y. Hsp60—A double edge sword in autoimmunity. Oncotarget 2015, 6, 32299–32300. [Google Scholar] [CrossRef] [PubMed]
- Van Eden, W.; Jansen, M.A.; Ludwig, I.; van Kooten, P.; van Der Zee, R.; Broere, F. The enigma of heat shock proteins in immune tolerance. Front. Immunol. 2017, 8, 1599. [Google Scholar] [CrossRef] [PubMed]
- Zanin-Zhorov, A.; Cahalon, L.; Tal, G.; Margalit, R.; Lider, O.; Cohen, I.R. Heat shock protein 60 enhances CD4+ CD25+ regulatory T cell function via innate TLR2 signaling. J. Clin. Investig. 2006, 116, 2022–2032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Kleer, I.M.; Kamphuis, S.M.; Rijkers, G.T.; Scholtens, L.; Gordon, G.; de Jager, W.; Häfner, R.; van de Zee, R.; van Eden, W.; Kuis, W.; et al. The spontaneous remission of juvenile idiopathic arthritis is characterized by CD30+ T cells directed to human heat-shock protein 60 capable of producing the regulatory cytokine interleukin-10. Arthritis Rheum. 2003, 48, 2001–2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kol, A.; Lichtman, A.H.; Finberg, R.W.; Libby, P.; Kurt-Jones, E.A. Cutting edge: Heat shock protein (Hsp) 60 activates the innate immune response: CD14 is an essential receptor for Hsp60 activation of mononuclear cells. J. Immunol. 2000, 164, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.P.; Jagadeesan, B.; Burkholder, K.M.; Jaradat, Z.W.; Wampler, J.L.; Lathrop, A.A.; Morgan, M.T.; Bhunia, A.K. Adhesion characteristics of Listeria adhesion protein (LAP)-expressing Escherichia coli to Caco-2 cells and of recombinant LAP to eukaryotic receptor Hsp60 as examined in a surface plasmon resonance sensor. FEMS Microbiol. Lett. 2006, 256, 324–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sreedhar, A.S.; Kalmár, É.; Csermely, P.; Shen, Y.F. Hsp90 isoforms: Functions, expression and clinical importance. FEBS Lett. 2004, 562, 11–15. [Google Scholar] [CrossRef]
- Epple, L.M.; Griffiths, S.G.; Dechkovskaia, A.M.; Dusto, N.L.; White, J.; Ouellette, R.J.; Anchordoquy, T.J.; Bemis, L.T.; Graner, M.W. Medulloblastoma exosome proteomics yield functional roles for extracellular vesicles. PLoS ONE 2012, 7, 42064. [Google Scholar] [CrossRef] [PubMed]
- Campanella, C.; Bavisotto, C.C.; Gammazza, A.M.; Nikolic, D.; Rappa, F.; David, S.; Cappello, F.; Bucchieri, F.; Fais, S. Exosomal heat shock proteins as new players in tumour cell-to-cell communication. J. Circ. Biomark. 2014, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Cid, C.; Regidor, I.; Poveda, P.D.; Alcazar, A. Expression of heat shock protein 90 at the cell surface in human neuroblastoma cells. Cell Stress Chaperones 2009, 14, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.B.; Feng, L.X.; Yue, Q.X.; Wu, W.Y.; Guan, S.H.; Jiang, B.H.; Yang, M.; Liu, X.; Guo, D.A. Paraptosis accompanied by autophagy and apoptosis was induced by celastrol, a natural compound with influence on proteasome, ER stress and Hsp90. J. Cell Physiol. 2012, 227, 2196–2206. [Google Scholar] [CrossRef] [PubMed]
- Graner, M.W. Hsp90 and immune modulation in cancer. Adv. Cancer Res. 2016, 129, 191–224. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Sahu, D.; Tsen, F. Secreted heat shock protein-90 (Hsp90) in wound healing and cancer. Biochim. Biophys. Acta 2012, 1823, 730–741. [Google Scholar] [CrossRef] [PubMed]
- Kovalchin, J.T.; Wang, R.; Wagh, M.S.; Azoulay, J.; Sanders, M.; Chandawarkar, R.Y. In vivo delivery of heat shock protein 70 accelerates wound healing by up-regulating macrophage-mediated phagocytosis. Wound Repair Regen. 2006, 14, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Multhoff, G.; Pockley, A.G.; Streffer, C.; Gaipl, U.S. Dual role of heat shock proteins (Hsps) in anti-tumor immunity. Curr. Mol. Med. 2012, 12, 1174–1182. [Google Scholar] [CrossRef] [PubMed]
- Oura, J.; Tamura, Y.; Kamiguchi, K.; Kutomi, G.; Sahara, H.; Torigoe, T.; Himi, T.; Sato, N. Extracellular heat shock protein 90 plays a role in translocating chaperoned antigen from endosome to proteasome for generating antigenic peptide to be cross-presented by dendritic cells. Int. Immunol. 2011, 23, 223–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokota, S.I.; Fujii, N. Immunomodulatory activity of extracellular heat shock proteins and their autoantibodies. Microbiol. Immunol. 2010, 54, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Laufen, T.; Mayer, M.P.; Beisel, C.; Klostermeier, D.; Reinstein, J.; Bukau, B. Mechanism of regulation of Hsp70 chaperones by DnaJ co-chaperones. Proc. Natl. Acad. Sci. USA 1999, 96, 5452–5457. [Google Scholar] [CrossRef] [PubMed]
- Botha, M.; Pesce, E.R.; Blatch, G.L. The Hsp40 proteins of Plasmodium falciparum and other apicomplexa: Regulating chaperone power in the parasite and the host. Int. J. Biochem. Cell Biol. 2007, 39, 1781–1803. [Google Scholar] [CrossRef] [PubMed]
- Njunge, J.M.; Mandal, P.; Przyborski, J.M.; Boshoff, A.; Pesce, E.-R.; Blatch, G.L. PFB0595w is a Plasmodium falciparum J protein that co-localizes with PfHsp70-1 and can stimulate its in vitro ATP hydrolysis activity. Int. J. Biochem. Cell Biol. 2015, 62, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Bascos, N.A.D.; Landry, S.J. Structural rigidity regulates functional interactions in the Hsp40-Hsp70 molecular machine. Biophys. J. 2015, 108, 210. [Google Scholar] [CrossRef]
- Njunge, J.M.; Ludewig, M.H.; Boshoff, A.; Pesce, E.; Blatch, G.L. Hsp70s and J proteins of Plasmodium parasites infecting rodents and primates: Structure, function, clinical relevance, and drug targets. Curr. Pharm. Des. 2013, 19, 387–403. [Google Scholar] [CrossRef] [PubMed]
- Gomes, P.S.; Bhardwaj, J.; Rivera-Correa, J.; Freire-De-Lima, C.G.; Morrot, A. Immune escape strategies of malaria parasites. Front. Microbiol. 2016, 7, 1617. [Google Scholar] [CrossRef] [PubMed]
- Tukaj, S.; Kotlarz, A.; Jozwik, A.; Smolenska, Z.; Bryl, E.; Witkowski, J.M.; Lipinska, B. Hsp40 proteins modulate humoral and cellular immune response in rheumatoid arthritis patients. Cell Stress Chaperones 2010, 15, 555–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, M.P. Hsp70 chaperone dynamics and molecular mechanism. Trends Biochem. Sci. 2013, 38, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Masison, D.C. Hsp70 structure, function, regulation and influence on yeast prions. Protein Pept. Lett. 2009, 16, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Easton, D.P.; Kaneko, Y.; Subjeck, J.R. The Hsp110 and Grp170 stress proteins: Newly recognized relatives of the Hsp70s. Cell Stress Chaperones 2000, 5, 276–290. [Google Scholar] [CrossRef]
- Mogk, A.; Tomoyasu, T.; Goloubinoff, P.; Rüdiger, S.; Röder, D.; Langen, H.; Bukau, B. Identification of thermolabile Escherichia coli proteins: Prevention and reversion of aggregation by DnaK and ClpB. EMBO J. 1999, 18, 6934–6949. [Google Scholar] [CrossRef] [PubMed]
- Vega, V.L.; Rodríguez-Silva, M.; Frey, T.; Gehrmann, M.; Diaz, J.C.; Steinem, C.; Multhoff, G.; Arispe, N.; de Maio, A. Hsp70 translocates into the plasma membrane after stress and is released into the extracellular environment in a membrane-associated form that activates macrophages. J. Immunol. 2008, 180, 4299–4307. [Google Scholar] [CrossRef] [PubMed]
- Binder, R.J. Heat-shock protein-based vaccines for cancer and infectious disease. Expert Rev. Vaccines 2008, 7, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Shevtsov, M.; Multhoff, G. Heat shock protein–peptide and Hsp-based immunotherapies for the treatment of cancer. Front. Immunol. 2016, 7, 171. [Google Scholar] [CrossRef] [PubMed]
- Multhoff, G.; Mizzen, L.; Winchester, C.C.; Milner, C.M.; Wenk, S.; Eissner, G.; Kampinga, H.H.; Laumbacher, B.; Johnson, J. Heat shock protein 70 (Hsp70) stimulates proliferation and cytolytic activity of natural killer cells. Exp. Hematol. 1999, 27, 1627–1636. [Google Scholar] [CrossRef]
- Gehrmann, M.; Liebisch, G.; Schmitz, G.; Anderson, R.; Steinem, C.; de Maio, A.; Pockley, G.; Multhoff, G. Tumor-specific Hsp70 plasma membrane localization is enabled by the glycosphingolipid Gb3. PLoS ONE 2008, 3, 1925. [Google Scholar] [CrossRef] [PubMed]
- Multhoff, G.; Pockley, A.G.; Schmid, T.E.; Schilling, D. The role of heat shock protein 70 (Hsp70) in radiation-induced immunomodulation. Cancer Lett. 2015, 368, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, R.A.; Williamson, P. Phosphatidylserine, a death knell. Cell Death Differ. 2001, 8, 551–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyurina, Y.Y.; Poloyac, S.M.; Tyurin, V.A.; Kapralov, A.A.; Jiang, J.; Anthonymuthu, T.S.; Kapralova, V.I.; Vikulina, A.S.; Jung, M.Y.; Epperly, M.W.; et al. A mitochondrial pathway for biosynthesis of lipid mediators. Nat. Chem. 2014, 6, 542. [Google Scholar] [CrossRef] [PubMed]
- de Maio, A. Extracellular heat shock proteins, cellular export vesicles, and the stress observation system: A form of communication during injury, infection, and cell damage. Cell Stress Chaperones 2011, 16, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Specht, H.M.; Ahrens, N.; Blankenstein, C.; Duell, T.; Fietkau, R.; Gaipl, U.S.; Günther, C.; Gunther, S.; Habl, G.; Hautmann, H.; et al. Heat shock protein 70 (Hsp70) peptide activated natural killer (NK) cells for the treatment of patients with non-small cell lung cancer (NSCLC) after radiochemotherapy (RCTx)–from preclinical studies to a clinical phase II trial. Front. Immunol. 2015, 6, 162. [Google Scholar] [CrossRef] [PubMed]
- Thériault, J.R.; Adachi, H.; Calderwood, S.K. Role of scavenger receptors in the binding and internalization of heat shock protein 70. J. Immunol. 2006, 177, 8604–8611. [Google Scholar] [CrossRef] [PubMed]
- Quintana, F.J.; Cohen, I.R. Heat shock proteins as endogenous adjuvants in sterile and septic inflammation. J. Immunol. 2005, 175, 2777–2782. [Google Scholar] [CrossRef] [PubMed]
- Asea, A.; Rehli, M.; Kabingu, E.; Boch, J.A.; Baré, O.; Auron, P.E.; Stevenson, M.A.; Calderwood, S.K. Novel signal transduction pathway utilized by extracellular HSP70: Role of TLR2 and TLR4. J. Biol. Chem. 2002, 277, 15028–15034. [Google Scholar] [CrossRef] [PubMed]
- Shonhai, A. The role of Hsp70s in the development and pathogenicity of Plasmodium species. Heat Shock Proteins Malar. 2014, 47–69. [Google Scholar] [CrossRef]
- Mabate, B.; Zininga, T.; Ramatsui, L.; Makumire, S.; Achilonu, I.; Dirr, H.W.; Shonhai, A. Structural and biochemical characterization of Plasmodium falciparum Hsp70-x reveals functional versatility of its C-terminal EEVN motif. Proteins 2018. [Google Scholar] [CrossRef] [PubMed]
- Pooe, O.J.; Kollisch, G.; Heine, H.; Shonhai, A. Plasmodium falciparum heat shock protein 70 lacks immune modulatory activity. Protein Pept. Lett. 2017, 24, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Moretta, L.; Bottino, C.; Pende, D.; Vitale, M.; Mingari, M.C.; Moretta, A. Human natural killer cells: Molecular mechanisms controlling NK cell activation and tumor cell lysis. Immunol. Lett. 2005, 100, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Multhoff, G.; Botzler, C.; Issels, R. The role of heat shock proteins in the stimulation of an immune response. Biol. Chem. 1998, 379, 295–300. [Google Scholar] [PubMed]
- Gastpar, R.; Gehrmann, M.; Bausero, M.A.; Asea, A.; Gross, C.; Schroeder, J.A.; Multhoff, G. Heat shock protein 70 surface-positive tumor exosomes stimulate migratory and cytolytic activity of natural killer cells. Cancer Res. 2005, 65, 5238–5247. [Google Scholar] [CrossRef] [PubMed]
- Gehrmann, M.; Doss, B.T.; Wagner, M.; Zettlitz, K.A.; Kontermann, R.E.; Foulds, G.; Pockley, A.G.; Multhoff, G. A novel expression and purification system for the production of enzymatic and biologically active human granzyme B. J. Immunol. Methods 2011, 371, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Gehrmann, M.; Stangl, S.; Kirschner, A.; Foulds, G.A.; Sievert, W.; Doss, B.T.; Walch, A.; Pockley, A.G.; Multhoff, G. Immunotherapeutic targeting of membrane Hsp70-expressing tumors using recombinant human granzyme B. PLoS ONE 2012, 7, 41341. [Google Scholar] [CrossRef] [PubMed]
- Kapelski, S.; de Almeida, M.; Fischer, R.; Barth, S.; Fendel, R. Antimalarial activity of Granzyme B and its targeted delivery by a Granzyme B–single-chain Fv fusion protein. Antimicrob. Agents Chemother. 2015, 59, 669–672. [Google Scholar] [CrossRef] [PubMed]
- Bulut, Y.; Michelsen, K.S.; Hayrapetian, L.; Naiki, Y.; Spallek, R.; Singh, M.; Arditi, M. Mycobacterium tuberculosis heat shock proteins use diverse Toll-like receptor pathways to activate pro-inflammatory signals. J. Biol. Chem. 2005, 280, 20961–20967. [Google Scholar] [CrossRef] [PubMed]
- Borges, T.J.; Murakami, N.; Machado, F.D.; Murshid, A.; Lang, B.J.; Lopes, R.L.; Bellan, L.M.; Uehara, M.; Antunes, K.H.; Pérez-Saéz, M.J.; et al. March1-dependent modulation of donor MHC II on CD103+ dendritic cells mitigates alloimmunity. Nat. Commun. 2018, 9, 3482. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Whittall, T.; McGowan, E.; Younson, J.; Kelly, C.; Bergmeier, L.A.; Singh, M.; Lehner, T. Identification of stimulating and inhibitory epitopes within the heat shock protein 70 molecule that modulate cytokine production and maturation of dendritic cells. J. Immunol. 2005, 174, 3306–3316. [Google Scholar] [CrossRef] [PubMed]
- Floto, R.A.; MacAry, P.A.; Boname, J.M.; Mien, T.S.; Kampmann, B.; Hair, J.R.; Huey, O.S.; Houben, E.N.; Pieters, J.; Day, C.; et al. Dendritic cell stimulation by mycobacterial Hsp70 is mediated through CCR5. Science 2006, 314, 454–458. [Google Scholar] [CrossRef] [PubMed]
- Babaahmady, K.; Oehlmann, W.; Singh, M.; Lehner, T. Inhibition of human immunodeficiency virus type 1 infection of human CD4+ T cells by microbial HSP70 and the peptide epitope 407-426. J. Virol. 2007, 81, 3354–3360. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Lin, Z.; Zhao, Z.J.; Yang, Y.; Niu, H.; Shen, X. MK2-mediated formation and phosphorylation-regulated dissociation of the signal complex consisting of p38, MK2, Akt and Hsp27. J. Biol. Chem. 2006, 281, 37215–37226. [Google Scholar] [CrossRef] [PubMed]
- Rane, M.J.; Pan, Y.; Singh, S.; Powell, D.W.; Wu, R.; Cummins, T.; Chen, Q.; McLeish, K.R.; Klein, J.B. Heat shock protein 27 controls apoptosis by regulating Akt activation. J. Biol. Chem. 2003, 278, 27828–27835. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Kausar, H.; Johnson, P.; Montoya-Durango, D.E.; Merchant, M.; Rane, M.J. Hsp27 regulates Akt activation and PMN apoptosis by scaffolding MK2 to Akt signal complex. J. Biol. Chem. 2007, 282, 21598–21608. [Google Scholar] [CrossRef] [PubMed]
- Rayner, K.; Chen, Y.X.; Siebert, T.; O’Brien, E.R. Heat shock protein 27: Clue to understanding estrogen-mediated atheroprotection? Trends Cardiovasc. Med. 2010, 20, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Behr, C.; Sarthou, J.L.; Rogier, C.; Trape, J.F.; Dat, M.H.; Michel, J.C.; Aribot, G.; Dieye, A.; Claverie, J.M.; Druihle, P. Antibodies and reactive T cells against the malaria heat-shock protein Pf72/Hsp70-1 and derived peptides in individuals continuously exposed to Plasmodium falciparum. J. Immunol. 1992, 149, 3321–3330. [Google Scholar] [PubMed]
- Marth, J.D.; Grewal, P.K. Mammalian glycosylation in immunity. Nat. Rev. Immunol. 2008, 8, 874–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsan, M.F.; Gao, B. Pathogen-associated molecular pattern contamination as putative endogenous ligands of Toll-like receptors. J. Endotoxin Res. 2007, 13, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Planesse, C.; Nativel, B.; Iwema, T.; Gasque, P.; Robert-Da Silva, C.; Viranaïcken, W. Recombinant human Hsp60 produced in ClearColi™ BL21 (DE3) does not activate the NFκB pathway. Cytokine 2015, 73, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Tsan, M.F. Recombinant human heat shock protein 60 does not induce the release of tumor necrosis factor α from murine macrophages. J. Biol. Chem. 2003, 278, 22523–22529. [Google Scholar] [CrossRef] [PubMed]
- Bausinger, H.; Lipsker, D.; Ziylan, U.; Manié, S.; Briand, J.P.; Cazenave, J.P.; Muller, S.; Haeuw, J.F.; Ravanat, C.; Salle, H.D.L.; et al. Endotoxin-free heat-shock protein 70 fails to induce APC activation. Eur. J Immunol. 2002, 32, 3708–3713. [Google Scholar] [CrossRef] [Green Version]
- Ye, Z.; Gan, Y.H. Flagellin contamination of recombinant heat shock protein 70 is responsible for its activity on T cells. J. Biol. Chem. 2007, 282, 4479–4484. [Google Scholar] [CrossRef] [PubMed]
- Bendz, H.; Marincek, B.C.; Momburg, F.; Ellwart, J.W.; Issels, R.D.; Nelson, P.J.; Noessner, E. Calcium signaling in dendritic cells by human or mycobacterial Hsp70 is caused by contamination and is not required for Hsp70-mediated enhancement of cross-presentation. J. Biol. Chem. 2008, 283, 26477–26483. [Google Scholar] [CrossRef] [PubMed]
- Wallin, R.P.; Lundqvist, A.; Moré, S.H.; von Bonin, A.; Kiessling, R.; Ljunggren, H.G. Heat-shock proteins as activators of the innate immune system. Trends Immunol. 2002, 23, 130–135. [Google Scholar] [CrossRef]
- Reed, R.C.; Berwin, B.; Baker, J.P.; Nicchitta, C.V. GRP94/gp96 elicits ERK activation in Murine macrophages a role for endotoxin contamination in NF-κB activation and nitric oxide production. J. Biol. Chem. 2003, 278, 31853–31860. [Google Scholar] [CrossRef] [PubMed]
- Fong, J.J.; Sreedhara, K.; Deng, L.; Varki, N.M.; Angata, T.; Liu, Q.; Nizet, V.; Varki, A. Immunomodulatory activity of extracellular Hsp70 mediated via paired receptors Siglec-5 and Siglec-14. EMBO J. 2015, 34, 2775–2788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zininga, T.; Pooe, O.J.; Makhado, P.B.; Ramatsui, L.; Prinsloo, E.; Achilonu, I.; Dirr, H.; Shonhai, A. Polymyxin B inhibits the chaperone activity of Plasmodium falciparum Hsp70. Cell Stress Chaperones 2017, 22, 707–715. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, M.A.; Troyanova, N.I.; Servuli, E.A.; Bolkhovitina, E.L.; Fedorina, A.S.; Sapozhnikov, A.M. Study of immunomodulatory effects of extracellular Hsp70 in a mouse model of allergic airway inflammation. Biochemistry 2016, 81, 1384–1395. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Lu, Y.; Tian, Q.Y.; Zhang, Y.; Guo, F.J.; Liu, G.Y.; Syed, N.M.; Lai, Y.; Lin, E.A.; Kong, L.; et al. The growth factor progranulin binds to TNF receptors and is therapeutic against inflammatory arthritis in mice. Science 2011, 332, 478–484. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proteins | Associated Pathology | Immunomodulatory Function | Ref |
|---|---|---|---|
| sHsps | Cancer | Phosphorylation of Hsp27 by mitogen-activated protein 2 (MK2) is implicated in signal transduction. Hsp27 regulates Akt (protein kinase B) activation. | [13] |
| Hsp40 | Pneumococcal infection | Stimulates Th1 and Th17 immune response against Streptococcus pneumoniae infection in mice. Activates BMDCs through recognition of TLR4 causing activation of MAPKs, NF-κB and PI3K-Akt pathways resulting in secretion of IFN-γ and IL-17A. | [14] |
| Induces pro-inflammatory cytokine production in macrophages. Activates PI3K and JNK signal pathways resulting in secretion of IL6. | [15] | ||
| Hsp60 | Type 1 diabetes mellitus | Induces both pro-inflammatory and anti-inflammatory cytokines. Binds multiple allelic variants of HLA-DR, this results in the release of IL-10, an anti-inflammatory cytokine, and IFN-γ. | [3] |
| Type 2 diabetes | Interaction of Hsp60 with TLR2 and TLR4 results in release of pro-inflammatory cytokines (IL-1β, IL-6, IL-8, MCP-1 and TNF-α). | [16] | |
| Hsp70 | Chronic inflammatory diseases | Promotes the production of anti-inflammatory cytokines. Interact with DCs, MDSCs, and monocytes, by binding to their endocytic receptors resulting in the release of anti-inflammatory cytokine IL-10 and inevitable immunosuppression. | [17] |
| Cancer | Acts as extracellular localized recognition site for NK cells. Interaction with NK cells through the TKD motif results in cytolytic attack mediated by NK cells. | [18] | |
| Hsp90 | Cancer | Hsp90 is implicated in T-cell mediated antitumor responses. Hsp90 inhibition up-regulates expression of interferon response genes, which promotes killing of melanoma cells by T cells. | [19] |
| Heat Shock Protein Implicated in Immunomodulation E | Contaminants Implicated in Study | References |
|---|---|---|
| Hsp60 induced pro-inflammatory cytokine production by macrophages | LPS | [20,112] |
| Hsp70 induced pro-inflammatory cytokine production by macrophages | LPS | [94,113] |
| Hsp70 induced activation and maturation of dendritic cells | LPS | [114] |
| Hsp70 and anti-CD-3 co-stimulation of IL-2 production by Jurkat T cells | Flagellin | [115] |
| Hsp70 induced, CCR5 mediated calcium signaling by dendritic cells | Nucleotides (ATP and ADP) | [116] |
| Hsp90 induced pro-inflammatory cytokine production by macrophages | LPS | [117] |
| Gp96 induced activation of NF-κB and production of NO by macrophages | LPS | [118] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zininga, T.; Ramatsui, L.; Shonhai, A. Heat Shock Proteins as Immunomodulants. Molecules 2018, 23, 2846. https://doi.org/10.3390/molecules23112846
Zininga T, Ramatsui L, Shonhai A. Heat Shock Proteins as Immunomodulants. Molecules. 2018; 23(11):2846. https://doi.org/10.3390/molecules23112846
Chicago/Turabian StyleZininga, Tawanda, Lebogang Ramatsui, and Addmore Shonhai. 2018. "Heat Shock Proteins as Immunomodulants" Molecules 23, no. 11: 2846. https://doi.org/10.3390/molecules23112846
APA StyleZininga, T., Ramatsui, L., & Shonhai, A. (2018). Heat Shock Proteins as Immunomodulants. Molecules, 23(11), 2846. https://doi.org/10.3390/molecules23112846