A Novel Method for the Quantification of White Wine Mannoproteins by a Competitive Indirect Enzyme-Linked Lectin Sorbent Assay (CI-ELLSA)

 ,
, 


Abstract
:
1. Introduction
2. Results and Discussion
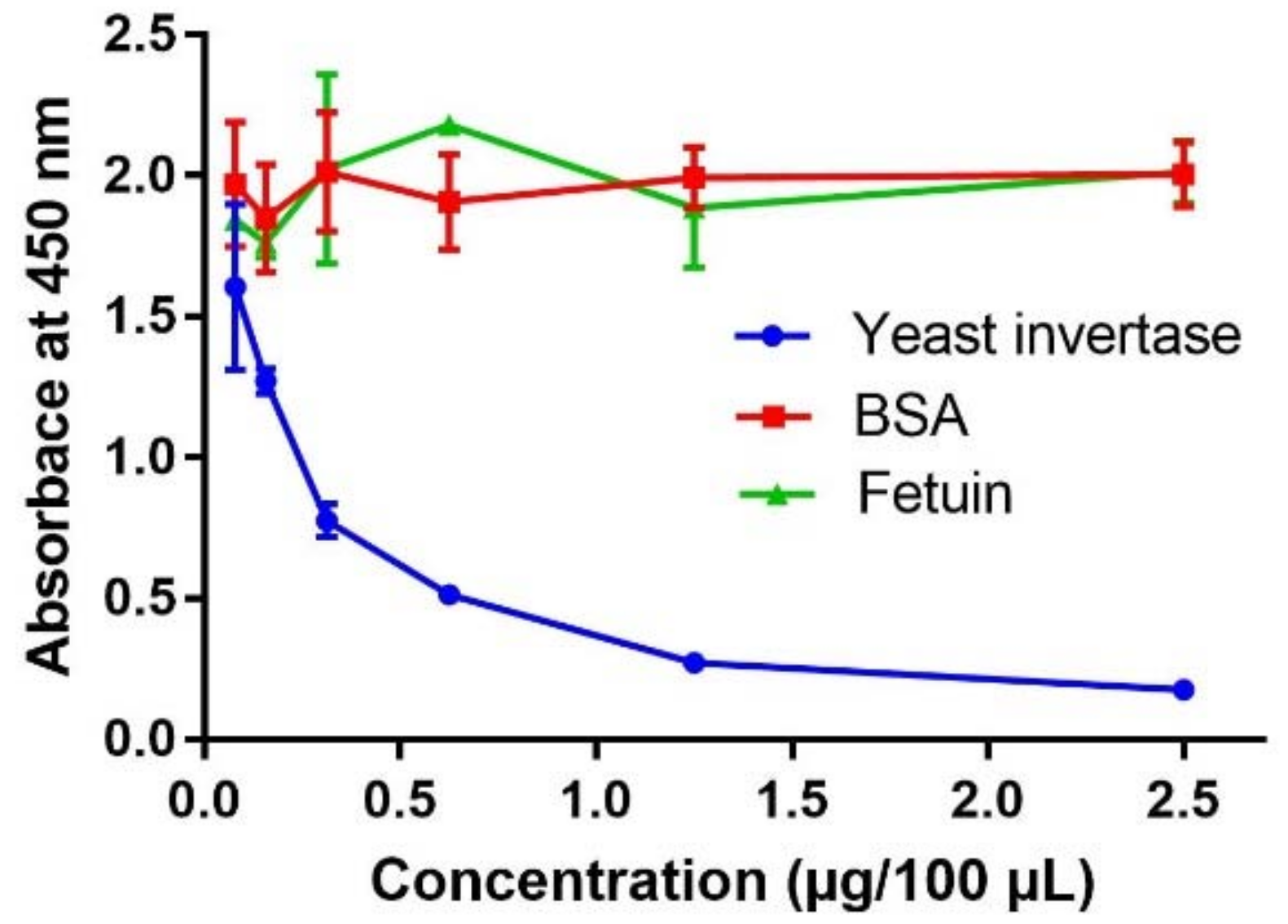
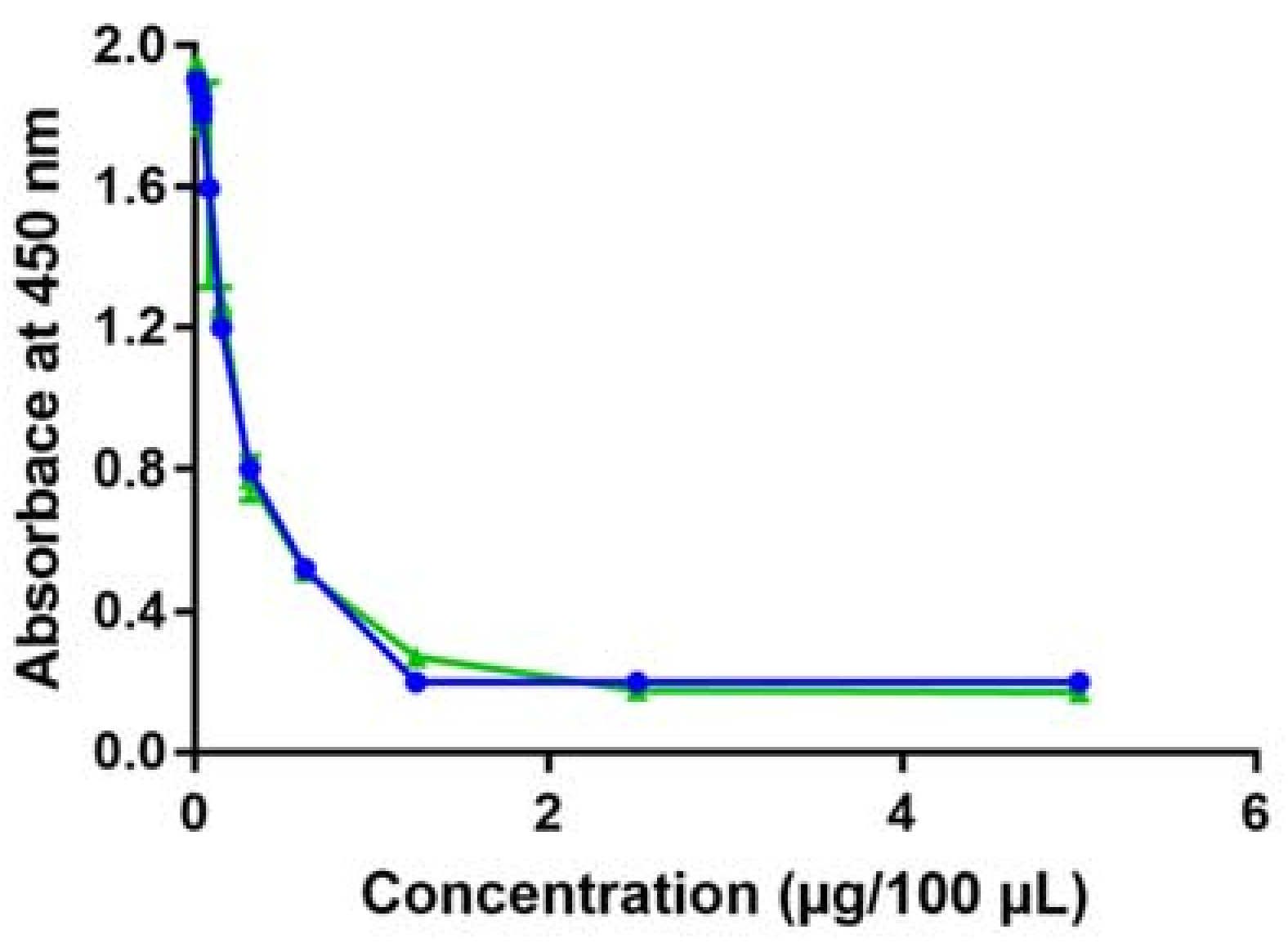
2.1. Determination of Optimal Conditions for the Competitive Indirect Enzyme-Linked Lectin Sorbent Assay (Method Development)
2.2. Recovery Test in Ultra-Filtered White Wine Spiked with Yeast Invertase
2.3. Mannoprotein Quantification in Different White Wine Samples
3. Materials and Methods
3.1. Materials and Sample Preparation
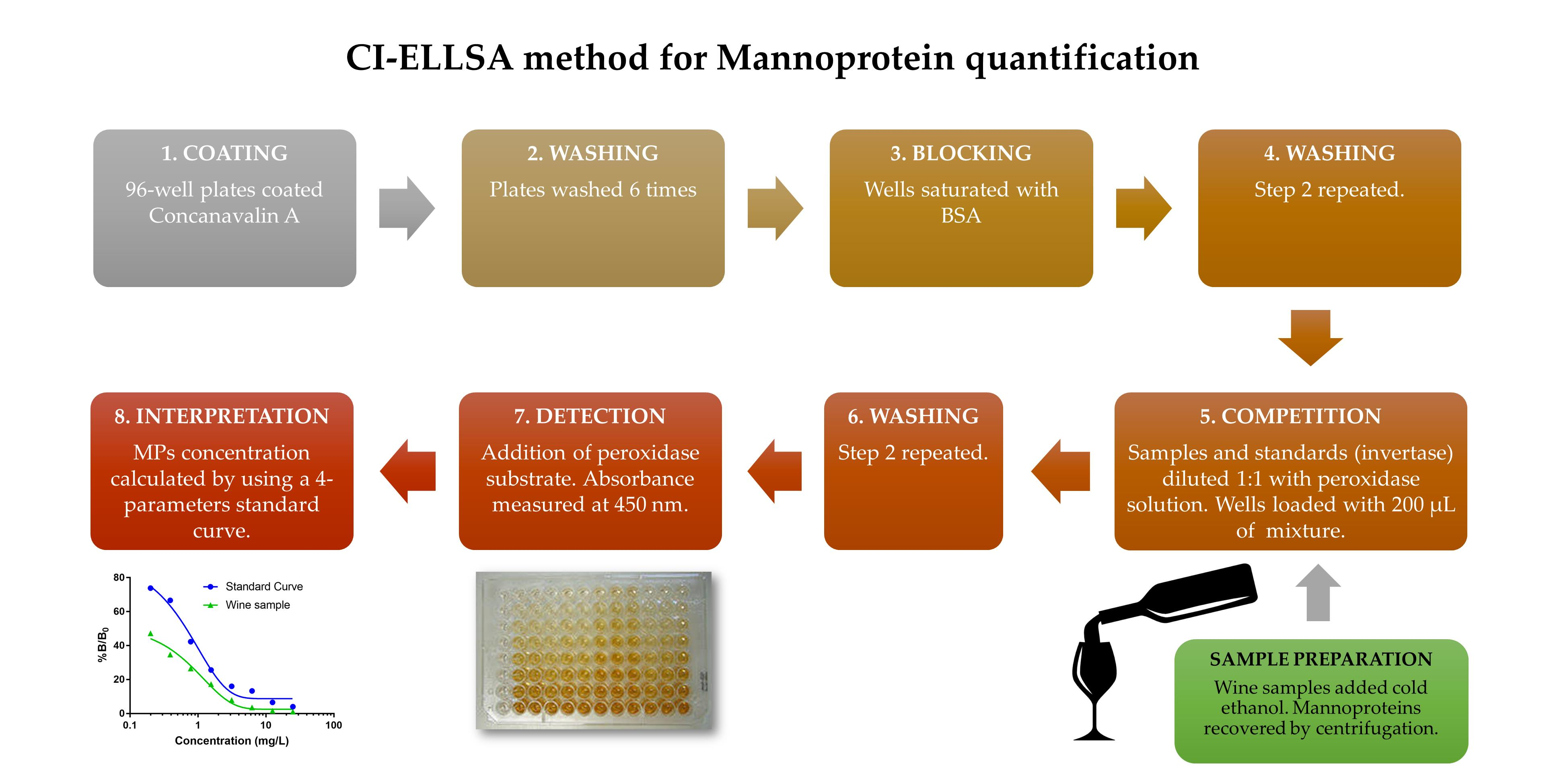
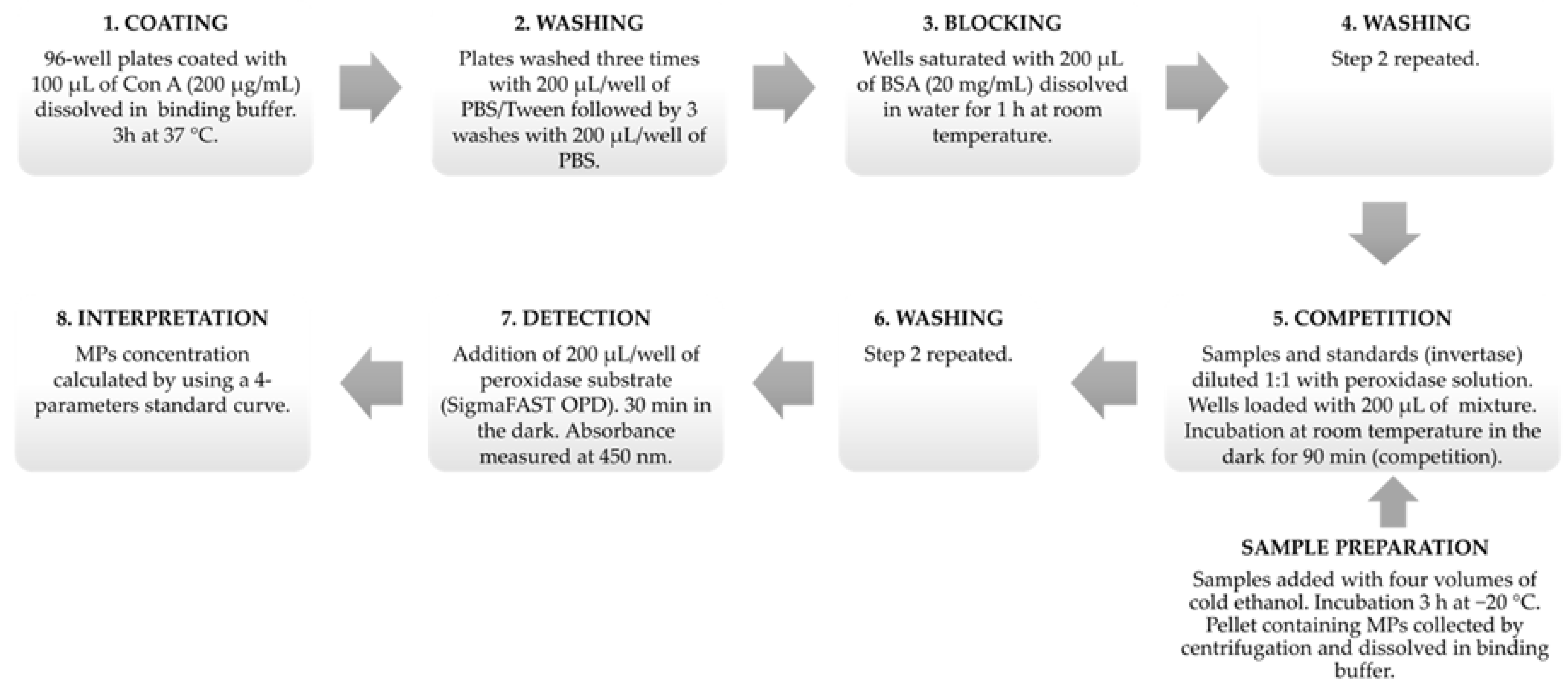
3.2. Competitive Indirect Enzyme-Linked Lectin Sorbent Assay (CI-ELLSA)
3.2.1. Preparation of Solutions
3.2.2. Protocol of Analysis
3.2.3. Four-Parameter Standard Curve
3.3. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vidal, S.; Williams, P.; Doco, T.; Moutounet, M.; Pellerin, P. The polysaccharides of red wine: Total fractionation and characterization. Carbohydr. Polym. 2003, 54, 439–447. [Google Scholar] [CrossRef]
- Guadalupe, Z.; Ayestaran, B. Polysaccharide profile and content during the vinification and aging of Tempranillo red wines. J. Agric. Food Chem. 2007, 55, 10720–10728. [Google Scholar] [CrossRef] [PubMed]
- Jégou, S.; Hoang, D.A.; Salmon, T.; Williams, P.; Oluwa, S.; Vrigneau, C.; Doco, T.; Marchal, R. Effect of grape juice press fractioning on polysaccharide and oligosaccharide compositions of Pinot meunier and Chardonnay Champagne base wines. Food Chem. 2017, 232, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Klis, F.M.; Boorsma, A.; De Groot, P.W.J. Cell wall construction in Saccharomyces cerevisiae. Yeast 2006, 23, 185–202. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, F.; Heyraud, A.; Norberta de Pinto, M.; Rinaudo, M. Characterization of white wine mannoproteins. J. Agric. Food Chem. 2002, 50, 6097–6101. [Google Scholar] [CrossRef] [PubMed]
- Doco, T.; Vuchot, P.; Cheynier, V.; Moutounet, M. Structural modification of wine arabinogalactans during aging on lees. Am. J. Enol. Vitic. 2003, 54, 150–157. [Google Scholar]
- Guadalupe, Z.; Ayestarán, B.N.; Williams, P.; Doco, T. Determination of must and wine polysaccharides by Gas Chromatography-Mass Spectrometry (GC-MS) and Size-Exclusion Chromatography (SEC). In Polysaccharides: Bioactivity and Biotechnology; Ramawat, K.M.J., Ed.; Springer International Publishing: Cham, Switzerland, 2014; pp. 1–29. ISBN 978-3-319-03751-6. [Google Scholar]
- Charpentier, C.; Dos Santos, A.M.; Feuillat, M. Release of macromolecules by Saccharomyces cerevisiae during ageing of French flor sherry wine “Vin jaune”. Int. J. Food Microbiol. 2004, 96, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Kemp, B.; Alexandre, H.; Robillard, B.; Marchal, R. Effect of production phase on bottle-fermented sparkling wine quality. J. Agric. Food Chem. 2015, 63, 19–38. [Google Scholar] [CrossRef] [PubMed]
- Alexandre, H.; Guilloux-Benatier, M. Yeast autolysis in sparkling wine—A review. Aust. J. Grape Wine Res. 2006, 12, 119–127. [Google Scholar] [CrossRef]
- Vincenzi, S.; Marangon, M.; Tolin, S.; Curioni, A. Protein evolution during the early stages of white winemaking and its relations with wine stability. Aust. J. Grape Wine Res. 2011, 17, 20–27. [Google Scholar] [CrossRef]
- Caridi, A. Enological functions of parietal yeast mannoproteins. Antonie Van Leeuwenhoek 2006, 89, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Waters, E.J.; Pellerin, P.; Brillouet, J.-M. A Saccharomyces mannoprotein that protects wine from protein haze. Carbohydr. Polym. 1994, 23, 185–191. [Google Scholar] [CrossRef]
- Dupin, I.V.; McKinnon, B.M.; Ryan, C.; Boulay, M.; Markides, A.J.; Jones, G.P.; Williams, P.J.; Waters, E.J. Saccharomyces cerevisiae mannoproteins that protect wine from protein haze: Their release during fermentation and lees contact and a proposal for their mechanism of action. J. Agric. Food Chem. 2000, 48, 3098–3105. [Google Scholar] [CrossRef] [PubMed]
- Van Sluyter, S.C.; McRae, J.M.; Falconer, R.J.; Smith, P.A.; Bacic, A.; Waters, E.J.; Marangon, M. Wine protein haze: Mechanisms of formation and advances in prevention. J. Agric. Food Chem. 2015, 63, 4020–4030. [Google Scholar] [CrossRef] [PubMed]
- Lubbers, S.; Leger, B.; Charpentier, C.; Feuillat, M. Effect des colloïdes-protecteurs d’extraits de parois de levures sur la stabilité tartrique d’un vin modèle. J. Int. Sci. Vigne Vin. 1993, 27, 13–22. [Google Scholar]
- Guilloux-Benatier, M.; Chassagne, D. Comparison of components released by fermented or active dried yeasts after aging on lees in a model wine. J. Agric. Food Chem. 2003, 51, 746–751. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Magariño, S.; Martínez-Lapuente, L.; Bueno-Herrera, M.; Ortega-Heras, M.; Guadalupe, Z.; Ayestarán, B. Use of commercial dry yeast products rich in mannoproteins for white and rosé sparkling wine elaboration. J. Agric. Food Chem. 2015, 63, 5670–5681. [Google Scholar] [CrossRef] [PubMed]
- Dufour, C.; Bayonove, C.L. Influence of wine structurally different polysaccharides on the volatility of aroma substances in a model system. J. Agric. Food Chem. 1999, 47, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Gawel, R.; Smith, P.A.; Waters, E.J. Influence of polysaccharides on the taste and mouthfeel of white wine. Aust. J. Grape Wine Res. 2016, 22, 350–357. [Google Scholar] [CrossRef]
- Vidal, S.; Francis, L.; Williams, P.; Kwiatkowski, M.; Gawel, R.; Cheynier, V.; Waters, E. The mouth-feel properties of polysaccharides and anthocyanins in a wine like medium. Food Chem. 2004, 85, 519–525. [Google Scholar] [CrossRef]
- Jones, P.R.; Gawel, R.; Francis, I.L.; Waters, E.J. The influence of interactions between major white wine components on the aroma, flavour and texture of model white wine. Food Qual. Prefer. 2008, 19, 596–607. [Google Scholar] [CrossRef]
- Vincenzi, S.; Crapisi, A.; Curioni, A. Foamability of Prosecco wine: Cooperative effects of high molecular weight glycocompounds and wine PR-proteins. Food Hydrocoll. 2014, 34, 202–207. [Google Scholar] [CrossRef]
- Rodrigues, A.; Ricardo-Da-Silva, J.M.; Lucas, C.; Laureano, O. Characterization of mannoproteins during white wine (Vitis vinifera L. cv. Encruzado) ageing on lees with stirring in oak wood barrels and in a stainless steel tank with oak staves. OENO One 2012, 46, 321–329. [Google Scholar] [CrossRef]
- Doco, T.; O’Neill, M.; Pellerin, P. Determination of the neutral and acidic glycosyl-residue compositions of plant polysaccharides by GC-EI-MS analysis of the trimethylsilyl methyl glycoside derivatives. Carbohydr. Polym. 2001, 46, 249–259. [Google Scholar] [CrossRef]
- Ayestarán, B.; Guadalupe, Z.; León, D. Quantification of major grape polysaccharides (Tempranillo v.) released by maceration enzymes during the fermentation process. Anal. Chim. Acta 2004, 513, 29–39. [Google Scholar] [CrossRef]
- Guadalupe, Z.; Martínez-Pinilla, O.; Garrido, Á.; Carrillo, J.D.; Ayestarán, B. Quantitative determination of wine polysaccharides by gas chromatography–mass spectrometry (GC–MS) and size exclusion chromatography (SEC). Food Chem. 2012, 131, 367–374. [Google Scholar] [CrossRef] [Green Version]
- Coimbra, M.A.; Barros, A.S.; Coelho, E.; Gonçalves, F.; Rocha, S.M.; Delgadillo, I. Quantification of polymeric mannose in wine extracts by FT-IR spectroscopy and OSC-PLS1 regression. Carbohydr. Polym. 2005, 61, 434–440. [Google Scholar] [CrossRef]
- Coimbra, M.A.; Gonçalves, F.; Barros, A.S.; Delgadillo, I. Fourier transform infrared spectroscopy and chemometric analysis of white wine polysaccharide extracts. J. Agric. Food Chem. 2002, 50, 3405–3411. [Google Scholar] [CrossRef] [PubMed]
- González-Royo, E.; Esteruelas, M.; Kontoudakis, N.; Fort, F.; Canals, J.M.; Zamora, F. The effect of supplementation with three commercial inactive dry yeasts on the colour, phenolic compounds, polysaccharides and astringency of a model wine solution and red wine. J. Sci. Food Agric. 2017, 97, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Quirós, M.; Gonzalez, R.; Morales, P. A simple method for total quantification of mannoprotein content in real wine samples. Food Chem. 2012, 134, 1205–1210. [Google Scholar] [CrossRef] [PubMed]
- Quiros, M.; Morales, P.; Perez-Traves, L.; Barcenilla, J.M.; Gonzalez, R. A new methodology to determine cell wall mannoprotein content and release in wine yeasts. Food Chem. 2011, 125, 760–766. [Google Scholar] [CrossRef]
- Vidal, S.; Williams, P.; Neill, M.A.O.; Pellerin, P. Polysaccharides from grape berry cell walls. Part I: Tissue distribution and structural characterization of the pectic polysaccharides. Carbohydr. Res. 2001, 45, 315–323. [Google Scholar] [CrossRef]
- Blakeney, A.B.; Harris, P.J.; Henry, R.J.; Stone, B.A. A simple and rapid preparation of alditol acetates for monosaccharide analysis. Carbohydr. Res. 1983, 113, 291–299. [Google Scholar] [CrossRef]
- Vidal, S.; Doco, T.; Moutounet, M.; Pellerin, P. Soluble polysaccharide content at initial time of experimental must preparation. Am. J. Enol. Vitic. 2000, 51, 115–121. [Google Scholar]
- Coelho, E.; Reis, A.; Domingues, M.R.M.; Rocha, S.M.; Coimbra, M.A. Synergistic effect of high and low molecular weight molecules in the foamability and foam stability of sparkling wines. J. Agric. Food Chem. 2011, 59, 3168–3179. [Google Scholar] [CrossRef] [PubMed]
- Ducasse, M.-A.; Canal-Llauberes, R.-M.; de Lumley, M.; Williams, P.; Souquet, J.-M.; Fulcrand, H.; Doco, T.; Cheynier, V. Effect of macerating enzyme treatment on the polyphenol and polysaccharide composition of red wines. Food Chem. 2010, 118, 369–376. [Google Scholar] [CrossRef]
- Brandão, E.; Silva, M.S.; García-Estévez, I.; Williams, P.; Mateus, N.; Doco, T.; de Freitas, V.; Soares, S. The role of wine polysaccharides on salivary protein-tannin interaction: A molecular approach. Carbohydr. Polym. 2017, 177, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.M.; Sugii, S.; Herp, A. A Guide for Carbohydrate Specificities of Lectins; Springer: Boston, MA, USA, 1988; pp. 819–847. [Google Scholar]
- Duk, M.; Lisowska, E.; Wu, J.H.; Wu, A.M. The Biotin/Avidin-Mediated Microtiter Plate Lectin Assay with the Use of Chemically Modified Glycoprotein Ligand. Anal. Biochem. 1994, 221, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.M.; Lisowska, E.; Duk, M.; Yang, Z. Lectins as tools in glycoconjugate research. Glycoconj. J. 2009, 26, 899–913. [Google Scholar] [CrossRef] [PubMed]
- Shannon, L.M.; Kay, E.; Lew, J.Y. Peroxidase isozymes from horseradish roots. I. Isolation and physical properties. J. Biol. Chem. 1966, 241, 2166–2172. [Google Scholar] [PubMed]
- Sasaki, D.; Mitchell, R.A. How to obtain reproducible quantitative ELISA results. Oxf. Biomed. Res. 2001, 1–11. [Google Scholar]
- Gascón, S.; Neumann, N.P.; Lampen, J.O. Comparative study of the properties of the purified internal and external invertases from yeast. J. Biol. Chem. 1968, 243, 1573–1577. [Google Scholar] [PubMed]
- Dambrouck, T.; Marchal, R.; Cilindre, C.; Parmentier, M.; Jeandet, P. Determination of the grape invertase content (using PTA-ELISA) following various fining treatments versus changes in the total protein content of wine. Relationships with wine foamability. J. Agric. Food Chem. 2005, 53, 8782–8789. [Google Scholar] [CrossRef] [PubMed]
- Dambrouck, T.; Marchal, R.; Marchal-Delahaut, L.; Parmentier, M.; Maujean, A.; Jeandet, P. Immunodetection of proteins from grapes and yeast in a white wine. J. Agric. Food Chem. 2003, 51, 2727–2732. [Google Scholar] [CrossRef] [PubMed]
- Hovasse, A.; Alayi, T.D.; Van Dorsselaer, A.; Marchal, R.; Jégou, S.; Schaeffer-Reiss, C. In-depth glycoproteomic characterisation of grape berry vacuolar invertase using a combination of mass spectrometry-based approaches. Food Chem. 2016, 200, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Moine-Ledoux, V.; Dubourdieu, D. An invertase fragment responsible for improving the protein stability of dry white wines. J. Sci. Food Agric. 1999, 79, 537–543. [Google Scholar] [CrossRef]
- Cilindre, C.; Jégou, S.; Hovasse, A.; Schaeffer, C.; Castro, A.J.; Clément, C.; Van Dorsselaer, A.; Jeandet, P.; Marchal, R. Van Proteomic Approach to Identify Champagne Wine Proteins as Modified by Botrytis cinerea Infection. J. Proteome Res. 2008, 7, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Núñez, Y.P.; Carrascosa, A.V.; Gonzalez, R.; Polo, M.C.; Martinez-Rodriguez, A. Isolation and characterization of a thermally extracted yeast cell wall fraction potentially useful for improving the foaming properties of sparkling wines. J. Agric. Food Chem. 2006, 54, 7898–7903. [Google Scholar] [CrossRef] [PubMed]
- Winzler, R.J. Glycoproteins. Plasma Proteins 1960, 1, 309–347. [Google Scholar] [CrossRef]
- Mitchell, P.A.; Miller, T.A.; Schmidt, K.L. Effects of alcohol on lectin binding affinity in rat gastric mucosa. Dig. Dis. Sci. 1990, 35, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Juega, M.; Gonzalez-Ramos, D.; Bartolome, B.; Carrascosa, A.V.; Martínez-Rodríguez, A.J. Chemical evaluation of white wines elaborated with a recombinant Saccharomyces cerevisiae strain overproducing mannoproteins. Food Chem. 2014, 147, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.P.; Divol, B. Tracking the careers of grape and wine polymers using biotechnology and systems biology. In Biotechnology in Functional Foods and Nutraceuticals; Bagchi, D., Lau, F.C., Ghosh, D.K., Eds.; CRC Press: Boca Raton, FL, USA, 2010; p. 18. ISBN 9781420087123. [Google Scholar]
- Waterhouse, A.L.; Sacks, G.L.; Jeffery, D.W. Understanding Wine Chemistry, 1st ed.; John Wiley & Sons, Ltd.: Chichester, UK, 2016. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spiked Yeast Invertase (mg/L) | Measured Yeast Invertase (mg/L) (Mean ± SD b) | Average B/B0 c | Average Recovery (%) | CV d (%) |
|---|---|---|---|---|
| 12.5 | 10.56 ± 0.85 | 36.8 | 84.5 | 8.1 |
| 6.25 | 6.84 ± 0.72 | 49.7 | 109.4 | 10.5 |
| 3.125 | 3.23 ± 0.34 | 71.2 | 103.3 | 10.6 |
| 1.5625 | 2.07 ± 0.36 | 73.4 | 132.8 | 17.2 |
| Yeast Strain for 2nd Fermentation | Mannoprotein Content (mg/L) (Mean ± SD) |
|---|---|
| IOC 18-2007 | 123.8 ± 7.6 d |
| AWRI1616 | 303.2 ± 2.4 a |
| AWRI1502 | 199.1 ± 4.2 b |
| AWRI1571 | 167.8 ± 14.9 c |
| F(3,8) | 231.3 |
| p value | <0.0001 |
| Wine Sample | Method of Production | Variety | Wine Style | MPs Content (mg/L) (Mean ± SD) |
|---|---|---|---|---|
| Manzoni bianco | White vinification with limited batonnage | Manzoni bianco | Dry white wine | 67.6 ± 4.8 |
| Sparkling Moscato | Asti method | Moscato bianco | Sweet sparkling wine | 88.8 ± 4.8 |
| Recioto passito | Withered grapes White vinification | Garganega | Sweet wine (passito style) | 18.1 ± 6.0 |
| Prosecco 1 | Charmat | Glera | Sparkling extra dry | 56.6 ± 1.8 |
| Prosecco 2 | Charmat | Glera | Sparkling extra dry | 49.0 ± 6.9 |
| Prosecco 3 | Charmat | Glera | Sparkling extra brut | 52.9 ± 4.1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marangon, M.; Vegro, M.; Vincenzi, S.; Lomolino, G.; De Iseppi, A.; Curioni, A. A Novel Method for the Quantification of White Wine Mannoproteins by a Competitive Indirect Enzyme-Linked Lectin Sorbent Assay (CI-ELLSA). Molecules 2018, 23, 3070. https://doi.org/10.3390/molecules23123070
Marangon M, Vegro M, Vincenzi S, Lomolino G, De Iseppi A, Curioni A. A Novel Method for the Quantification of White Wine Mannoproteins by a Competitive Indirect Enzyme-Linked Lectin Sorbent Assay (CI-ELLSA). Molecules. 2018; 23(12):3070. https://doi.org/10.3390/molecules23123070
Chicago/Turabian StyleMarangon, Matteo, Mara Vegro, Simone Vincenzi, Giovanna Lomolino, Alberto De Iseppi, and Andrea Curioni. 2018. "A Novel Method for the Quantification of White Wine Mannoproteins by a Competitive Indirect Enzyme-Linked Lectin Sorbent Assay (CI-ELLSA)" Molecules 23, no. 12: 3070. https://doi.org/10.3390/molecules23123070
APA StyleMarangon, M., Vegro, M., Vincenzi, S., Lomolino, G., De Iseppi, A., & Curioni, A. (2018). A Novel Method for the Quantification of White Wine Mannoproteins by a Competitive Indirect Enzyme-Linked Lectin Sorbent Assay (CI-ELLSA). Molecules, 23(12), 3070. https://doi.org/10.3390/molecules23123070