The Investigation of the Antitumor Agent Toxicity and Capsaicin Effect on the Electron Transport Chain Enzymes, Catalase Activities and Lipid Peroxidation Levels in Lung, Heart and Brain Tissues of Rats


Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Animals
2.3. Cisplatin and Capsaicin Treatment
2.4. Cisplatin Levels
2.5. Isolation of Mitochondria
2.6. Succinate Dehydrogenase Activity Assay
2.7. Cytochrome c Oxidase Activity Assay
2.8. The Cytosolic Preparation for CAT Activity
2.9. Lipid Peroxidation
2.10. Nucleotide Levels
2.11. Protein Determination
2.12. Statistical Analysis
3. Results
3.1. Levels of Platin in Different Tissues of Male Sprague Dawley Adult Rats
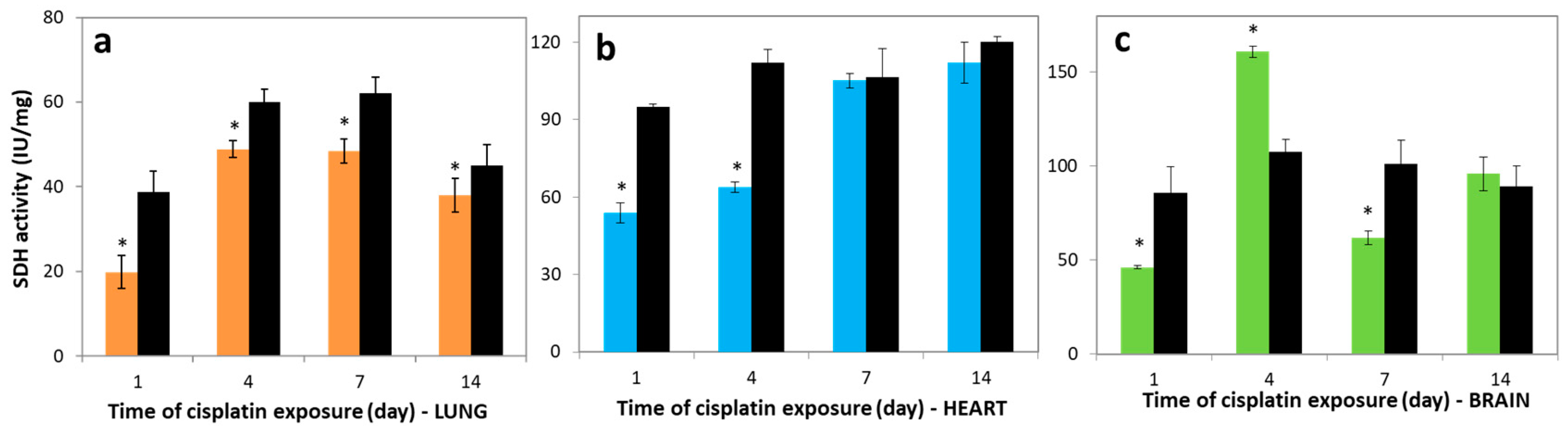
3.2. Variations in SDH Activities
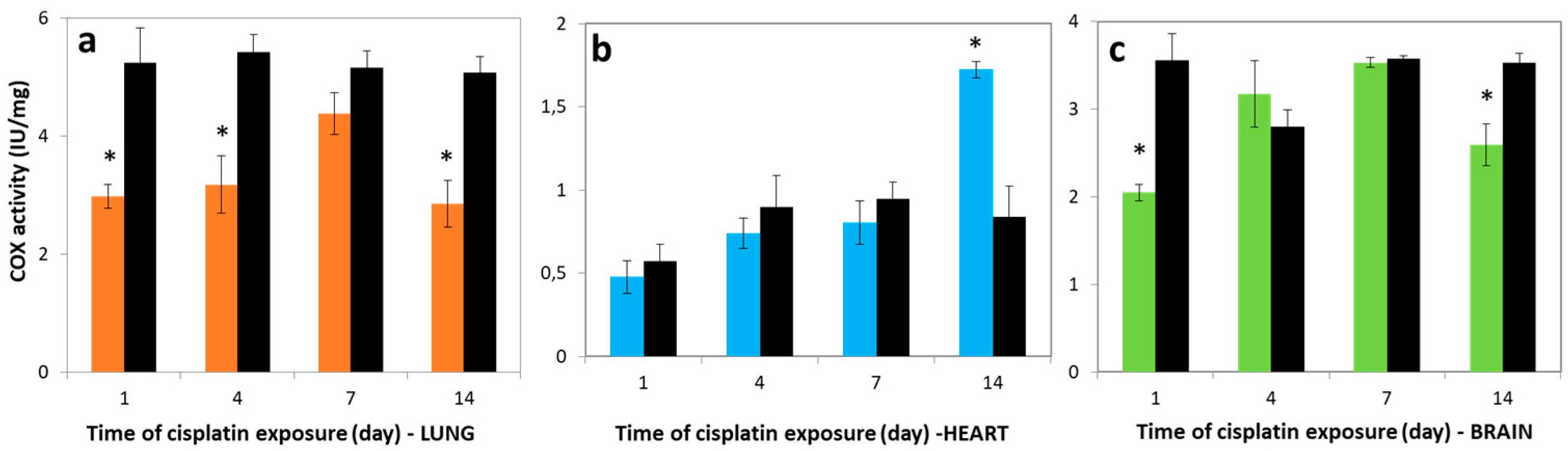
3.3. Variations in COX Activities
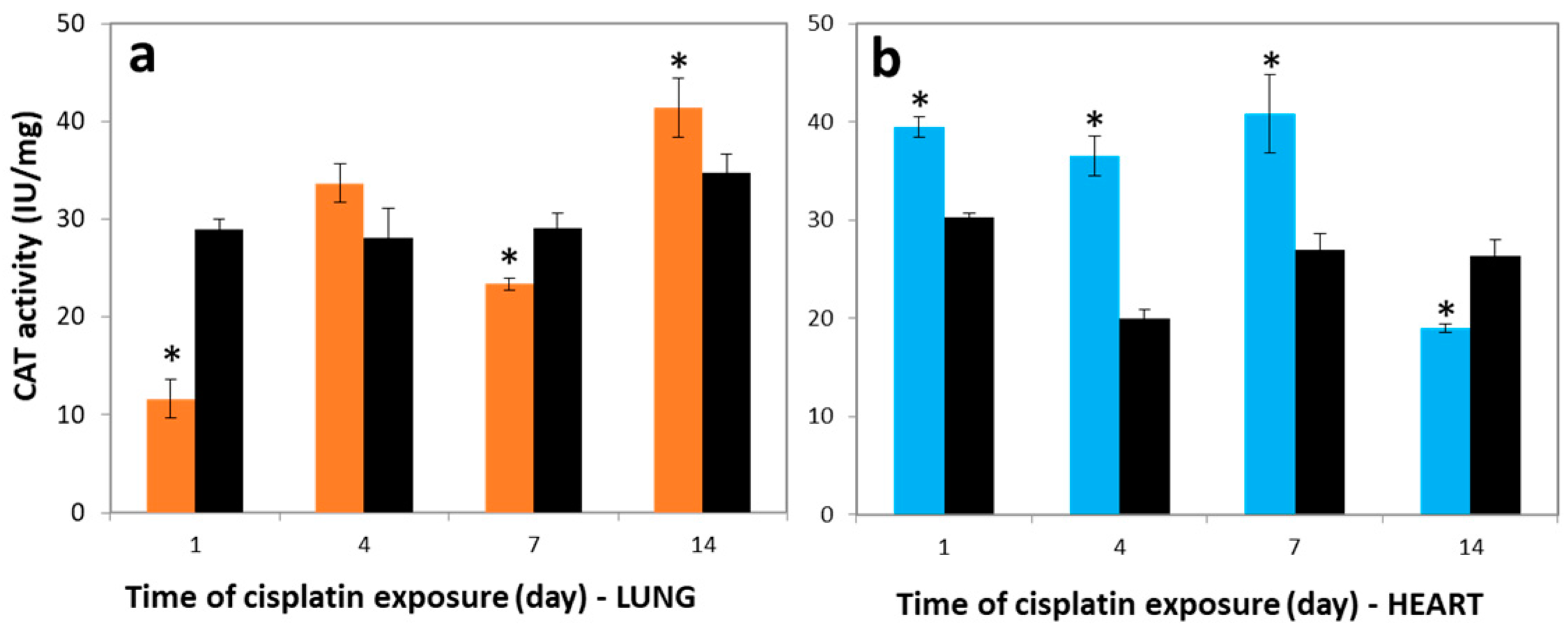
3.4. Variations in CAT Activities
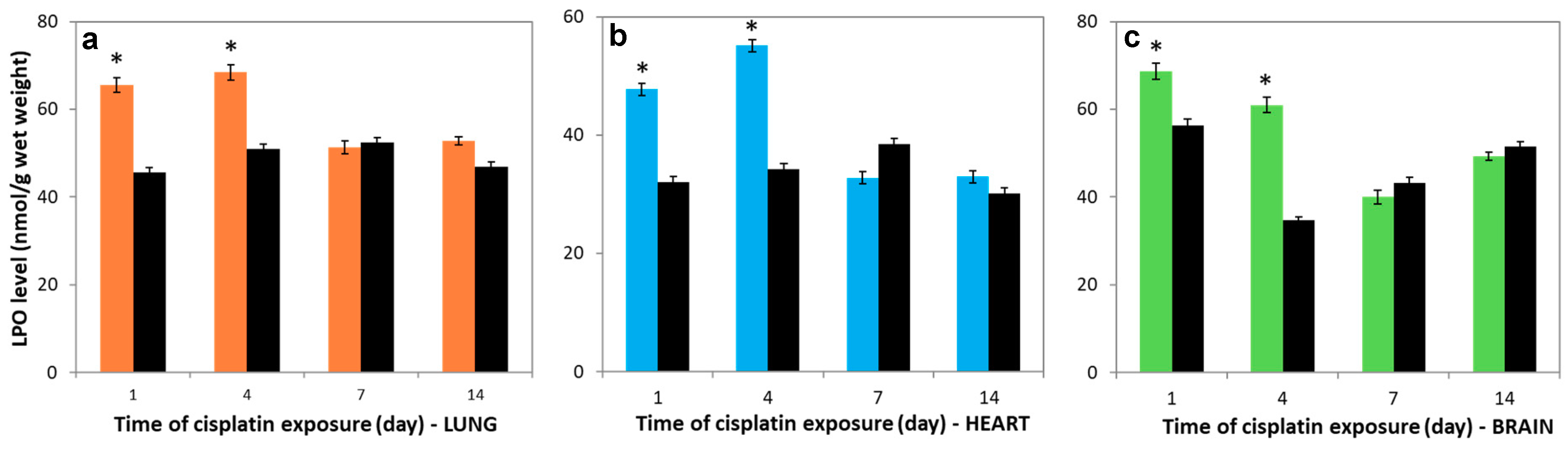
3.5. Variations in LPO levels
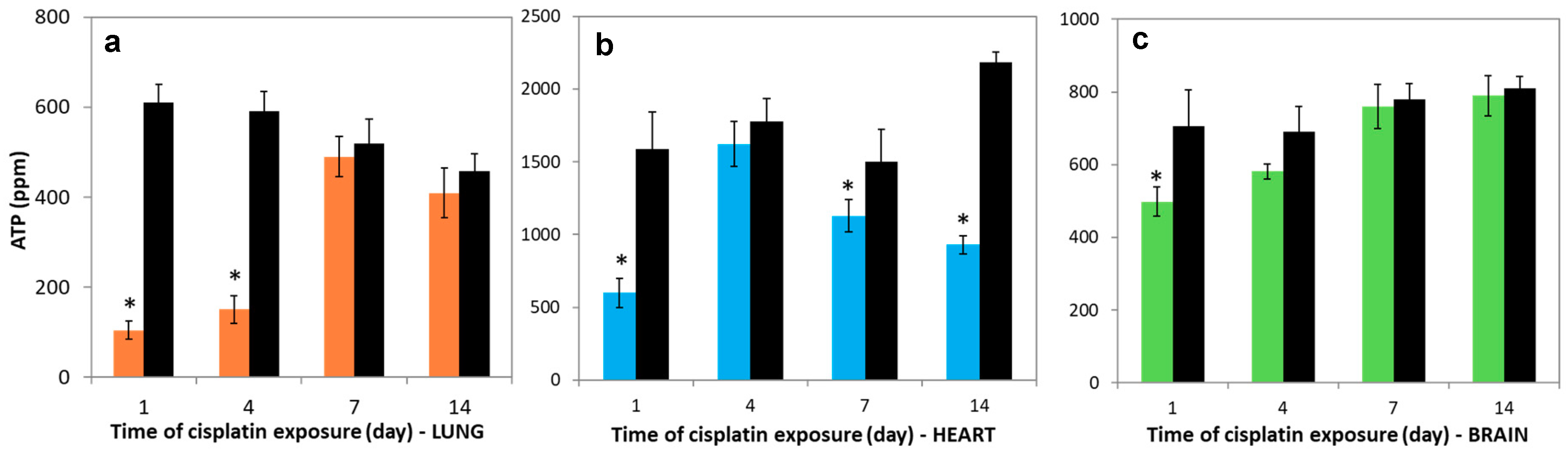
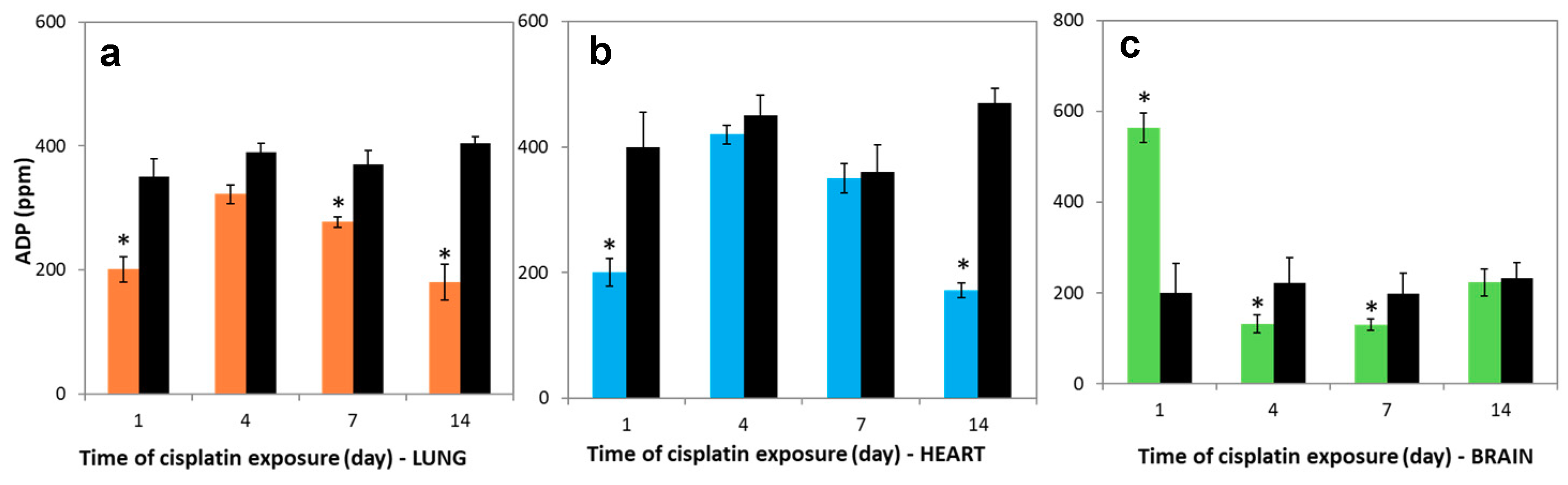
3.6. Variations in Nucleotides Levels
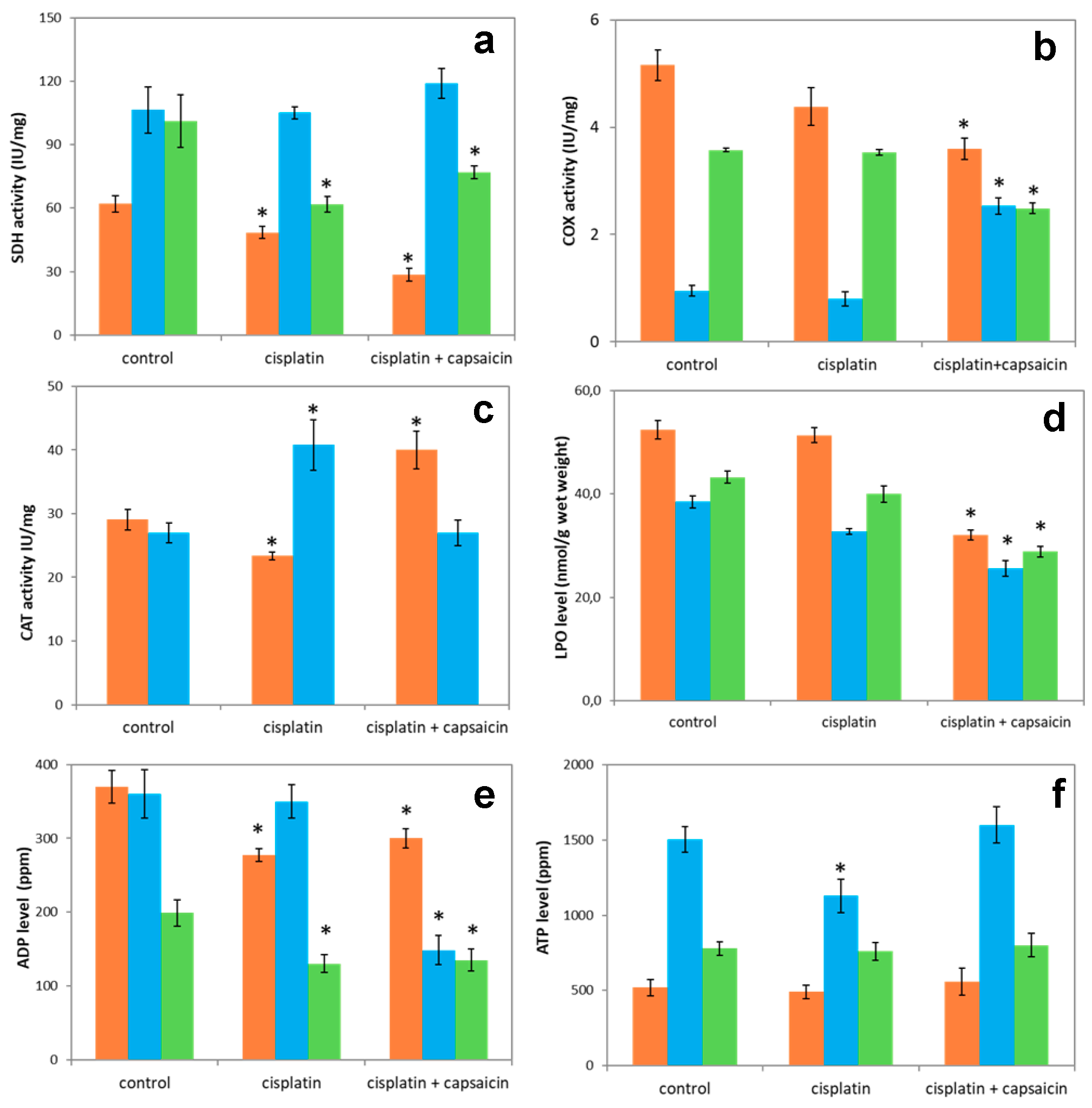
3.7. The Enzyme Activities of SDH, COX, CAT, and LPO and Nucleotide Levels in Lung, Heart, and Brain of Rats Treated with Cisplatin and Capsaicin
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Karp, G. Cell and Molecular Biology: Concepts and Experiments, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1999. [Google Scholar]
- Fruehauf, J.P.; Meyskens, F.L. Reactive oxygen species: A breath of life or death? Clin. Cancer Res. 2007, 13, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Reactive Species and Antioxidants. Redox Biology Is a Fundamental Theme of Aerobic Life. Plant Physiol. 2016, 141, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Sato, E.F.; Nishikawa, M.; Park, A.M.; Kira, Y.; Imada, I.; Utsumi, K. Mitochondrial generation of reactive oxygen species and its role in aerobic life. Curr. Med. Chem. 2003, 10, 2495–2505. [Google Scholar] [CrossRef] [PubMed]
- Lippard, S.J. 40 Years of DNA Double Helix Structural and biological consequences of platinum anticancer drug binding to DNA. In Proceedings of the Robert A. Welch Foundation Conference on Chemical Research Houston, Houston, TX, USA, 25–26 October 1993; pp. 49–60. [Google Scholar]
- Singh, K.K. Mitochondria damage checkpoint, aging, and cancer. Ann. N. Y. Acad. Sci. 2006, 1067, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Turrens, J.F. Mitochondrial formation of reactive oxygen species. J. Physiol. 2003, 552, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Brady, H.R.; Kone, B.C.; Stromski, M.E.; Zeidel, M.L.; Giebisch, G.; Gullans, S.R. Mitochondrial injury: An early event in cisplatin toxicity to renal proximal tubules. Am. J. Physiol. 1990, 258, 1181–1187. [Google Scholar] [CrossRef] [PubMed]
- Baliga, R.; Ueda, N.; Walker, P.D.; Shah, S.V. Oxidant mechanisms in toxic acute renal failure. Am. J. Kidney Dis. 1997, 29, 465–477. [Google Scholar] [CrossRef]
- Lau, A.H. Apoptosis induced by cisplatin nephrotoxic injury. Kidney Int. 1999, 56, 1295–1298. [Google Scholar] [CrossRef] [PubMed]
- Appenroth, D.; Frob, S.; Kersten, L.; Splinter, F.K.; Winnefeld, K. Protective effects of vitamin E and C on cisplatin nephrotoxicity in developing rats. Arch. Toxicol. 1997, 71, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Roller, A.; Weller, M. Antioxidants specifically inhibit cisplatin cytotoxicity of human malignant glioma cells. Anticancer Res. 1998, 18, 4493–4497. [Google Scholar] [PubMed]
- Lieberthal, W.; Triaca, V.; Levine, J. Mechanisms of death induced by cisplatin in proximal tubular epithelial cells: Apoptosis vs. necrosis. Am. J. Physiol. 1996, 270, 700–708. [Google Scholar] [CrossRef] [PubMed]
- Tsutsumishita, Y.; Onda, T.; Okada, K.; Takeda, M.; Endou, H.; Futaki, S.; Niwa, M. Involvement of H2O2 production in cisplatin-induced nephrotoxicity. Biochem. Biophys. Res. Commun. 1998, 242, 310–312. [Google Scholar] [CrossRef] [PubMed]
- McGinness, J.E.; Proctor, P.H.; Demopoulos, H.B.; Hokanson, J.A.; Kirkpatrick, D.S. Amelioration of cis-platinum nephrotoxicity by orgotein (superoxide dismutase). Physiol. Chem. Phys. 1978, 10, 267–277. [Google Scholar] [PubMed]
- Tanaka-Kagawa, T.; Kitahara, J.; Seko, Y.; Toyoda, H.; Imura, N.; Naganuma, A. Reduced sensitivity of HeLa cells to cis-platinum by simultaneous overexpression of copper, zinc-superoxide dismutase and catalase. Biochem. Pharmacol. 1999, 57, 545–548. [Google Scholar] [CrossRef]
- Gómez-Sierra, T.; Eugenio-Pérez, D.; Sánchez-Chinchillas, A.; Pedraza-Chaverri, J. Role of food-derived antioxidants against cisplatin induced-nephrotoxicity. Food Chem. Toxicol. 2018, 120, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Mashhadi, M.A.; Arab, M.R.; Azizi, F.; Shahraki, M.R. Histological Study of Toxic Effects of Cisplatin Single Dose Injection on Rat Kidney. Gene Cell Tissue. 2014, 1, e21536. [Google Scholar] [CrossRef]
- Shimeda, Y.; Hirotani, Y.; Akimoto, Y.; Shindou, K.; Ijiri, Y.; Nishihori, T.; Tanaka, K. Protective effects of capsaicin against cisplatin-induced nephrotoxicity in rats. Biol. Pharm. Bull. 2005, 28, 1635–1638. [Google Scholar] [CrossRef] [PubMed]
- Hatefi, Y.; Galante, Y.M. Isolation of cytochrome b560 from complex II (succinate ubiquinone oxidoreductase) and its reconstitution with succinate dehydrogenase. J. Biol. Chem. 1980, 255, 5530–5537. [Google Scholar] [PubMed]
- Wharton, D.C.; Tzagoloff, A. Cytochrome oxidase from beef heart mitochondria. Meth. Enzymol. 1967, 10, 245–250. [Google Scholar]
- Aebi, H. Catalase. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Academic Press: New York, NY, USA, 1974; pp. 673–685. [Google Scholar]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. In Methods in Enzymology; Colowick, S.P., Kaplan, N.O., Eds.; Academic Press: New York, NY, USA, 1978; pp. 302–316. [Google Scholar]
- Cardoso, S.M.; Pereira, C.; Oliveira, R. Mitochondrial function is differentially affected upon oxidative stress. Free Radic. Biol. Med. 1999, 26, 3–13. [Google Scholar] [CrossRef]
- Masubuchi, Y.; Suda, C.; Horie, T. Involvement of mitochondrial permeability transition in acetaminophen-induced liver injury in mice. J. Hepatol. 2005, 42, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Ganzera, M.; Vrabl, P.; Wörle, E.; Burgstaller, W.; Stuppner, H. Determination of adenine and pyridine nucleotides in glucose-limited chemostat cultures of Penicillium simplicissimum by one-step ethanol extraction and ion-pairing liquid chromatography. Anal. Biochem. 2006, 359, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of proteins utilizing the principle of protein–dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Kartalou, M.; Essigmann, J.M. Mechanisms of resistance to cisplatin. Mutat. Res. 2001, 478, 23–43. [Google Scholar] [CrossRef]
- Golpich, M.; Amini, E.; Mohamed, Z.; Ali, R.A.; Ibrahim, N.M.; Ahmadiani, A. Mitochondrial Dysfunction and Biogenesis in Neurodegenerative diseases: Pathogenesis and Treatment. CNS Neurosci. Ther. 2017, 23, 5–22. [Google Scholar] [CrossRef] [PubMed]
- Atli, G.; Alptekin, Ö.; Tükel, S.; Canli, M. Response of catalase activity to Ag+, Cd2+, Cr6+, Cu2+ and Zn2+ in five tissues of freshwater fish Oreochromis niloticus. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 143, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Chirra, H.D.; Sexton, T.; Biswal, D.; Hersh, L.B.; Hilt, J.Z. Catalase-coupled gold nanoparticles: Comparison between the carbodiimide and biotin-streptavidin methods. Acta Biomater. 2011, 7, 2865–2872. [Google Scholar] [CrossRef] [PubMed]
- Glorieux, C.; Calderon, P.B. Catalase, a remarkable enzyme: Targeting the oldest antioxidant enzyme to find a new cancer treatment approach. Biol. Chem. 2017, 26, 1095–1108. [Google Scholar] [CrossRef] [PubMed]
- Uysal, N.; Gönenç, S.; Sönmez, A.; Aksu, İ.; Topçu, A.; Kayatekin, B.M.; Açıkgöz, O. Adölesan sıçan beyninde antioksidan enzim aktiviteleri ve lipid peroksidasyon düzeyleri. Ege Tıp Dergisi. 2005, 44, 75–79. [Google Scholar]
- Cerri, S.; Piccolini, V.M.; Santin, G.; Bottone, M.G.; De Pascali, S.A.; Migoni, D.; Iadarola, P.; Fanizzi, F.P.; Bernocchi, G. The developmental neurotoxicity study of platinum compounds. Effects of cisplatin versus a novel Pt(II) complex on rat cerebellum. Neurotoxicol. Teratol. 2011, 33, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Kachadourian, R.; Leitner, H.M.; Day, B.J. Selected flavonoids potentiate the toxicity of cisplatin in human lung adenocarcinoma cells: A role for glutathione depletion. Int. J. Oncol. 2007, 31, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Praetz, M.; Laghmiche, M.; Pollakis, G.; Goormaghtigh, E.; Ruysschaert, J.M. In vivo and in vitro modifications of the mitochondrial membrane induced by 4′ Epi-adriamycin. Biochem. Pharmacol. 1986, 35, 2923–2928. [Google Scholar] [CrossRef]
- Gerschenson, M.; Paik, C.Y.; Gaukler, E.L.; Diwan, B.A.; Poirier, M.C. Cisplatin exposure induces mitochondrial toxicity in pregnant rats and their fetuses. Reprod. Toxicol. 2001, 15, 525–531. [Google Scholar] [CrossRef]
- Kong, L.K.; Sohal, R.S. Age-related changes in activities of mitochondrial electron transport complexes in various tissues of the mouse. Arch. Biochem. Biophys. 2000, 373, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Pratibha, R.; Sameer, R.; Rataboli, P.V.; Bhiwgade, D.A.; Dhume, C.Y. Enzymatic studies of cisplatin induced oxidative stress in hepatic tissue of rats. Eur. J. Pharmacol. 2006, 532, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Oral, H.B.; George, A.J.T.; Haskard, D.O. Prevention of hydrogen peroxide and cisplatin induced apoptosis by intracellular catalase overexpression. Turk. J. Biol. 2000, 24, 685–696. [Google Scholar]
- Jena, B.S.; Nayak, S.B.; Patnaik, B.K. Age-related changes in catalase activity and its inhibition by manganase (II) chloride in the brain of two species of poikilothermic vertebrates. Arch. Gerontol. Geriatr. 1998, 26, 119–129. [Google Scholar] [CrossRef]
- Srivastava, A.N.; Gupta, A.; Srivastava, S.; Natu, S.M.; Mittal, B.; Negi, M.P.S.; Prasad, R. Cisplatin combination chemotherapy induces oxidative stress in advance non small cell lung cancer patients. Asian Pac. J. Cancer Prev. 2010, 11, 465–471. [Google Scholar] [PubMed]
- Yasuyuki, S.; Takahiro, S.; Yoshio, T. Change of lipid peroxide levels in rat tissues after cisplatin administration. Toxicol. Lett. 1991, 57, 159–166. [Google Scholar] [CrossRef]
- Prasad, S.B.; Rosangkima, G.; Kharbangar, A. Structural and biochemical changes in mitochondria after cisplatin treatment of Dalton’s lymphoma-bearing mice. Mitochondrion 2010, 10, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Cersosimo, R.J. Hepatotoxicity associated with cisplatin chemotherapy. Ann. Pharmacother. 1993, 27, 438–441. [Google Scholar] [CrossRef] [PubMed]
- Daley-Yates, P.T.; McBrien, D.C.H. The inhibition of renal ATPase by cisplatin and some bio-transformation products. Chem. Biol. Interact. 1982, 40, 325–334. [Google Scholar] [CrossRef]
- Arabaci, B.; Gülçin, I.; Alwasel, S. Capsaicin: A potent inhibitor of carbonic anhydrase isoenzymes. Molecules 2014, 19, 10103–10114. [Google Scholar] [CrossRef] [PubMed]
- Galano, A.; Martínez, A. Capsaicin, a tasty free radical scavenger: Mechanism of action and kinetics. J. Phys. Chem. B 2012, 116, 1200–1208. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.K.; Vij, A.S.; Sharma, M. Mechanisms and clinical uses of capsaicin. Eur. J. Pharmacol. 2013, 720, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Surh, Y.J.; Lee, S.S. Capsaicin, a double-edged sword: Toxicity, metabolism, and chemopreventive potential. Life Sci. 1995, 56, 1845–1855. [Google Scholar] [CrossRef]
- Kogure, K.; Goto, S.; Nishimura, M.; Yasumoto, M.; Abe, K.; Ohiwa, C.; Sassa, H.; Kusumi, T.; Terada, H. Mechanism of potent antiperoxidative effect of capsaicin. Biochim. Biophys. Acta 2002, 1573, 84–92. [Google Scholar] [CrossRef]
- Lee, C.Y.; Kim, M.; Yoon, S.W.; Lee, C.H. Short-term control of capsaicin on blood and oxidative stress of rats in vivo. Phytother. Res. 2003, 17, 454–458. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.H.; Kim, H.J.; Oh, G.S.; Shen, A.; Lee, S.; Choe, S.K.; Park, R.; So, H.S. Capsaicin ameliorates cisplatin-induced renal injury through induction of heme oxygenase-1. Mol. Cells 2014, 37, 234–240. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lung | Heart | Brain | |
|---|---|---|---|
| 1st Day (ppm platin) | 0.24 | 0.15 | 0.24 |
| 4th Day (ppm platin) | 0.23 | 0.039 | Non Detectable |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kursunluoglu, G.; Taskiran, D.; Ayar Kayali, H. The Investigation of the Antitumor Agent Toxicity and Capsaicin Effect on the Electron Transport Chain Enzymes, Catalase Activities and Lipid Peroxidation Levels in Lung, Heart and Brain Tissues of Rats. Molecules 2018, 23, 3267. https://doi.org/10.3390/molecules23123267
Kursunluoglu G, Taskiran D, Ayar Kayali H. The Investigation of the Antitumor Agent Toxicity and Capsaicin Effect on the Electron Transport Chain Enzymes, Catalase Activities and Lipid Peroxidation Levels in Lung, Heart and Brain Tissues of Rats. Molecules. 2018; 23(12):3267. https://doi.org/10.3390/molecules23123267
Chicago/Turabian StyleKursunluoglu, Gizem, Dilek Taskiran, and Hulya Ayar Kayali. 2018. "The Investigation of the Antitumor Agent Toxicity and Capsaicin Effect on the Electron Transport Chain Enzymes, Catalase Activities and Lipid Peroxidation Levels in Lung, Heart and Brain Tissues of Rats" Molecules 23, no. 12: 3267. https://doi.org/10.3390/molecules23123267
APA StyleKursunluoglu, G., Taskiran, D., & Ayar Kayali, H. (2018). The Investigation of the Antitumor Agent Toxicity and Capsaicin Effect on the Electron Transport Chain Enzymes, Catalase Activities and Lipid Peroxidation Levels in Lung, Heart and Brain Tissues of Rats. Molecules, 23(12), 3267. https://doi.org/10.3390/molecules23123267