
Inhibitory Effect of Naphthoquinone-Tryptophan Hybrid towards Aggregation of PAP f39 Semen Amyloid

Abstract
:1. Introduction
2. Results and Discussion
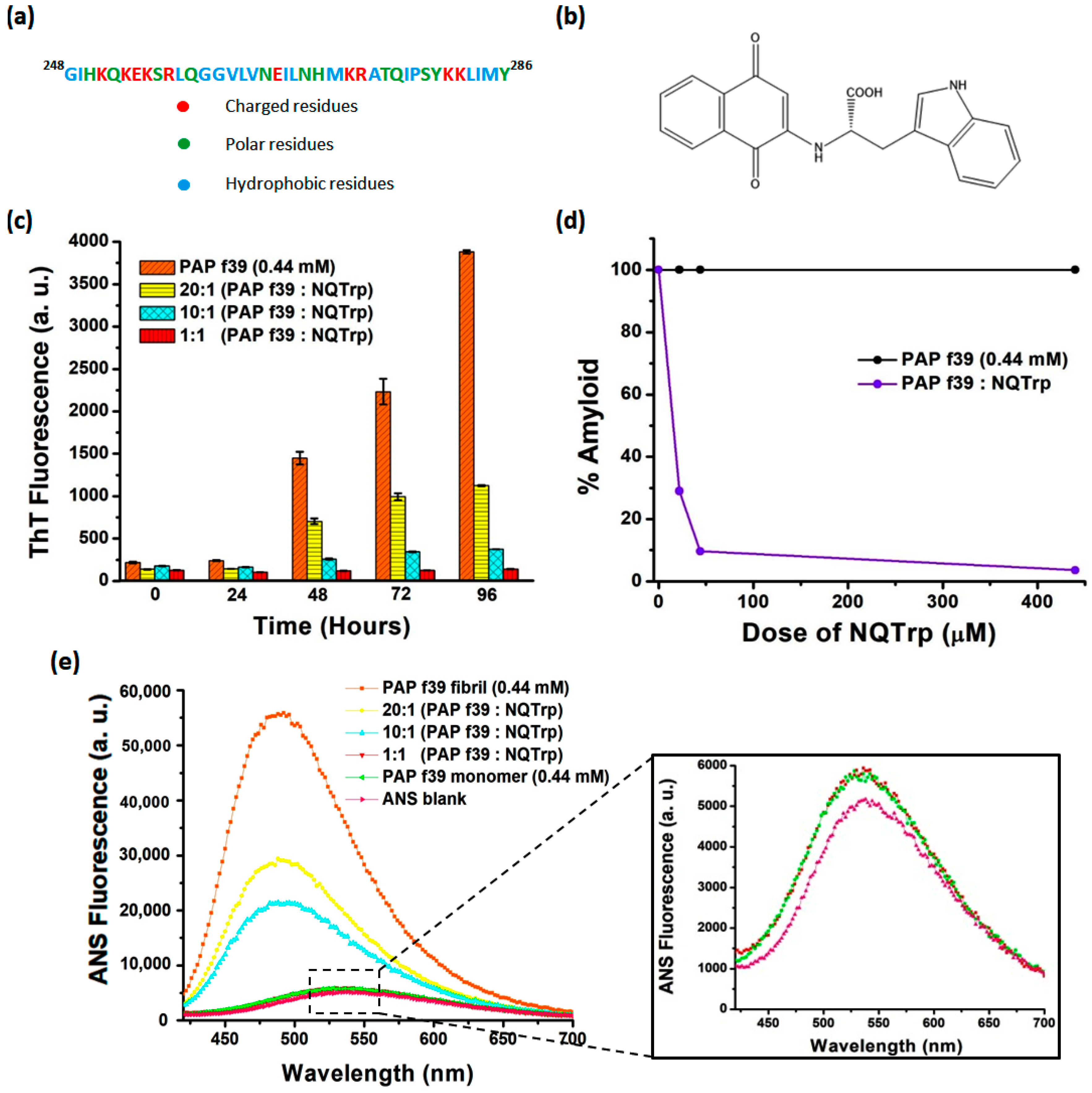
2.1. NQTrp Demonstrates Dose-Dependent Inhibition toward PAP f39 Amyloid Aggregation
2.2. NQTrp Retains the Native Conformation of the PAP f39 Monomers
2.3. Congo Red Birefringence Revealed That NQTrp Minimize PAP f39 Amyloid Deposits
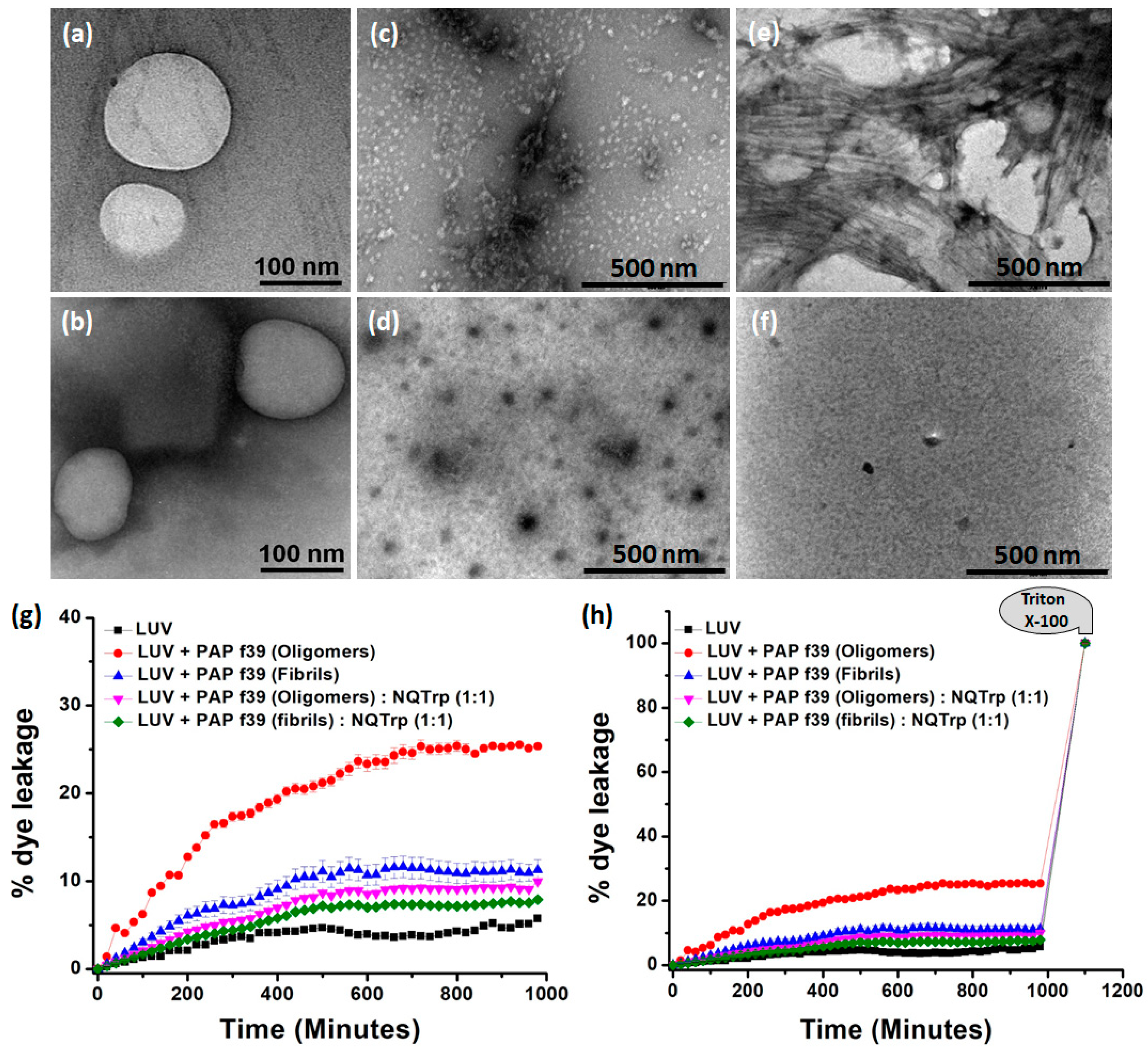
2.4. Morphology of the Inhibited PAP f39 Assemblies
2.5. Modulating PAP f39 Aggregation by NQTrp Renders Non-Toxic Intermediates
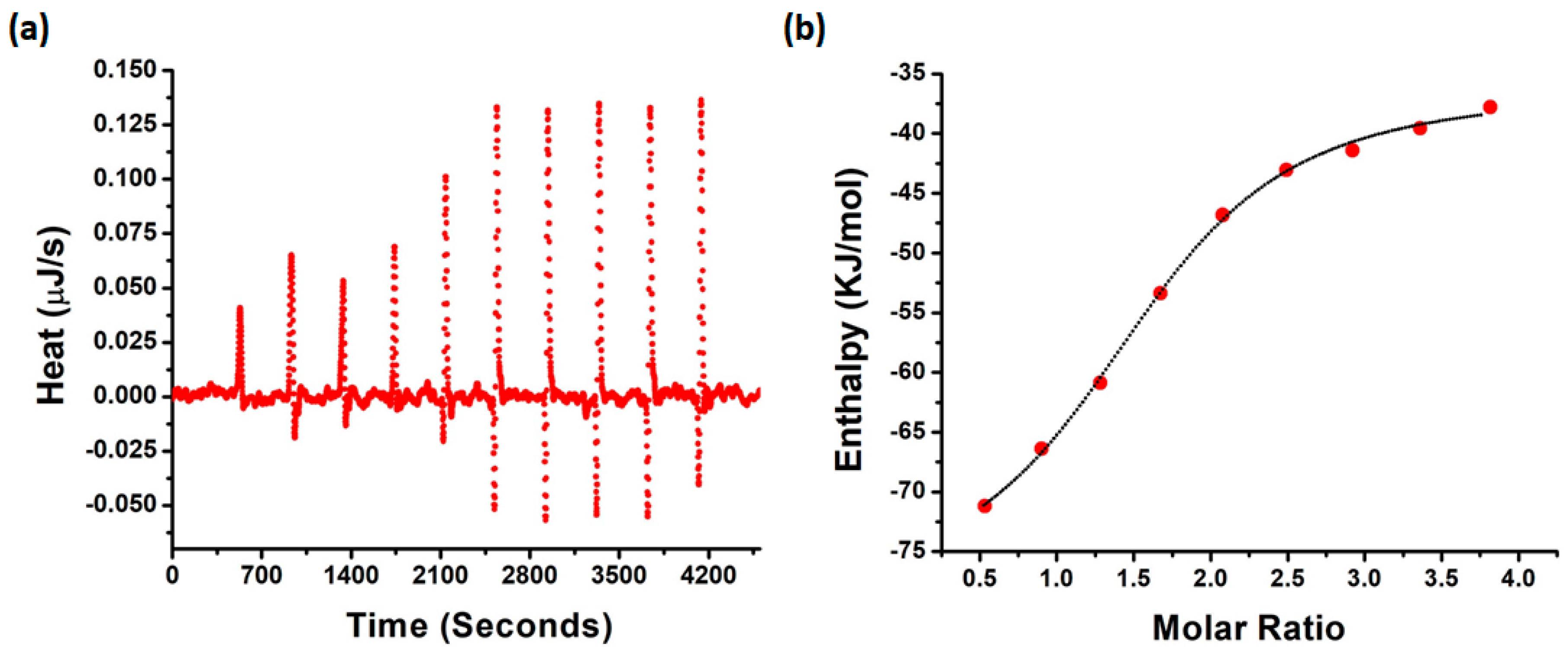
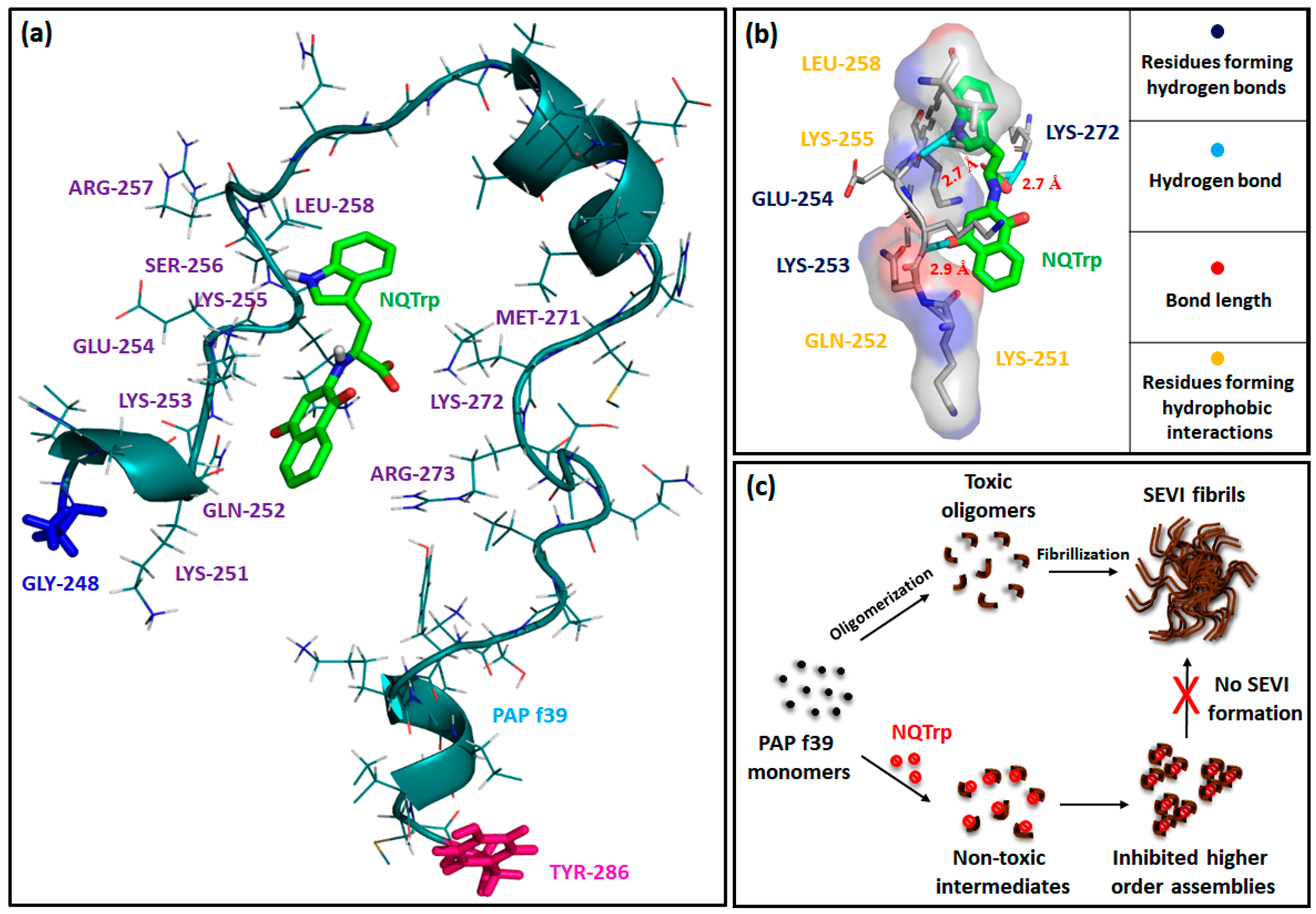
2.6. Interaction of NQTrp with PAP f39 Is Spontaneous and Involves Non-Covalent Contacts with Polar and Charged Amino Acid Residues
3. Materials and Methods
3.1. Materials
3.2. Stock Preparation
3.3. ThT Fluorescence-Based PAP f39 Aggregation and Inhibition Assay
3.4. 8-Anilinonaphthalene-1-Sulfonic Acid (ANS) Binding Assay
3.5. Congo Red Birefringence Assay
3.6. Transmission Electron Microscopy
3.7. Vesicle Dye Leakage Assay
3.8. Isothermal Titration Calorimetry
3.9. Molecular Docking
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gray, R.H.; Wawer, M.J.; Brookmeyer, R.; Sewankambo, N.K.; Serwadda, D.; Wabwire-Mangen, F.; Lutalo, T.; Li, X.; vanCott, T.; Quinn, T.C. Probability of HIV-1 transmission per coital act in monogamous, heterosexual, HIV-1-discordant couples in Rakai, Uganda. Lancet 2001, 357, 1149–1153. [Google Scholar] [CrossRef]
- Pilcher, C.D.; Tien, H.C.; Eron, J.J.J.; Vernazza, P.L.; Leu, S.-Y.; Stewart, P.W.; Goh, L.-E.; Cohen, M.S. Brief but efficient: Acute HIV infection and the sexual transmission of HIV. J. Infect. Dis. 2004, 189, 1785–1792. [Google Scholar] [CrossRef] [PubMed]
- Munch, J.; Rucker, E.; Standker, L.; Adermann, K.; Goffinet, C.; Schindler, M.; Wildum, S.; Chinnadurai, R.; Rajan, D.; Specht, A.; et al. Semen-derived amyloid fibrils drastically enhance HIV infection. Cell 2007, 131, 1059–1071. [Google Scholar] [CrossRef] [PubMed]
- Röcker, A.; Roan, N.R.; Yadav, J.K.; Fändrich, M.; Münch, J. Structure, function and antagonism of semen amyloids. Chem. Commun. 2018, 54, 7557–7569. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-A.; Yolamanova, M.; Zirafi, O.; Roan, N.R.; Staendker, L.; Forssmann, W.-G.; Burgener, A.; Dejucq-Rainsford, N.; Hahn, B.H.; Shaw, G.M.; et al. Semen-mediated enhancement of HIV infection is donor-dependent and correlates with the levels of SEVI. Retrovirology 2010, 7, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellano, L.M.; Shorter, J. The Surprising Role of Amyloid Fibrils in HIV Infection. Biology 2012, 1, 58–80. [Google Scholar] [CrossRef] [Green Version]
- Olsen, J.S.; DiMaio, J.T.M.; Doran, T.M.; Brown, C.; Nilsson, B.L.; Dewhurst, S. Seminal plasma accelerates semen-derived enhancer of viral infection (SEVI) fibril formation by the prostatic acid phosphatase (PAP248-286) peptide. J. Biol. Chem. 2012, 287, 11842–11849. [Google Scholar] [CrossRef]
- Roan, N.R.; Greene, W.C. A seminal finding for understanding HIV transmission. Cell 2007, 131, 1044–1046. [Google Scholar] [CrossRef]
- Capule, C.C.; Brown, C.; Olsen, J.S.; Dewhurst, S.; Yang, J. Oligovalent amyloid-binding agents reduce SEVI-mediated enhancement of HIV-1 infection. J. Am. Chem. Soc. 2012, 134, 905–908. [Google Scholar] [CrossRef]
- Hauber, I.; Hohenberg, H.; Holstermann, B.; Hunstein, W.; Hauber, J. The main green tea polyphenol epigallocatechin-3-gallate counteracts semen-mediated enhancement of HIV infection. Proc. Natl. Acad. Sci. USA 2009, 106, 9033–9038. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Dong, X.; Liu, Y.; Sun, Y. Brazilin Inhibits Prostatic Acidic Phosphatase Fibrillogenesis and Decreases its Cytotoxicity. Chem. Asian J. 2017, 12, 1062–1068. [Google Scholar] [CrossRef] [PubMed]
- LoRicco, J.G.; Xu, C.S.; Neidleman, J.; Bergkvist, M.; Greene, W.C.; Roan, N.R.; Makhatadze, G.I. Gallic Acid Is an Antagonist of Semen Amyloid Fibrils That Enhance HIV-1 Infection. J. Biol. Chem. 2016, 291, 14045–14055. [Google Scholar] [CrossRef] [PubMed]
- Roan, N.R.; Sowinski, S.; Münch, J.; Kirchhoff, F.; Greene, W.C. Aminoquinoline Surfen Inhibits the Action of SEVI (Semen-derived Enhancer of Viral Infection). J. Biol. Chem. 2010, 285, 1861–1869. [Google Scholar] [CrossRef] [PubMed]
- Olsen, J.S.; Brown, C.; Capule, C.C.; Rubinshtein, M.; Doran, T.M.; Srivastava, R.K.; Feng, C.; Nilsson, B.L.; Yang, J.; Dewhurst, S. Amyloid-binding Small Molecules Efficiently Block SEVI (Semen-derived Enhancer of Virus Infection)- and Semen-mediated Enhancement of HIV-1 Infection. J. Biol. Chem. 2010, 285, 35488–35496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xun, T.; Li, W.; Chen, J.; Yu, F.; Xu, W.; Wang, Q.; Yu, R.; Li, X.; Zhou, X.; Lu, L.; et al. ADS-J1 Inhibits Semen-Derived Amyloid Fibril Formation and Blocks Fibril-Mediated Enhancement of HIV-1 Infection. Antimicrob. Agents Chemother. 2015, 59, 5123–5134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lump, E.; Castellano, L.M.; Meier, C.; Seeliger, J.; Erwin, N.; Sperlich, B.; Sturzel, C.M.; Usmani, S.; Hammond, R.M.; von Einem, J.; et al. A molecular tweezer antagonizes seminal amyloids and HIV infection. Elife 2015, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widera, M.; Klein, A.N.; Cinar, Y.; Funke, S.A.; Willbold, D.; Schaal, H. The D-amino acid peptide D3 reduces amyloid fibril boosted HIV-1 infectivity. AIDS Res. Ther. 2014, 11, 1. [Google Scholar] [CrossRef] [Green Version]
- Sievers, S.A.; Karanicolas, J.; Chang, H.W.; Zhao, A.; Jiang, L.; Zirafi, O.; Stevens, J.T.; Munch, J.; Baker, D.; Eisenberg, D. Structure-based design of non-natural amino-acid inhibitors of amyloid fibril formation. Nature 2011, 475, 96–100. [Google Scholar] [CrossRef] [Green Version]
- Sheftic, S.R.; Snell, J.M.; Jha, S.; Alexandrescu, A.T. Inhibition of semen-derived enhancer of virus infection (SEVI) fibrillogenesis by zinc and copper. Eur. Biophys. J. 2012, 41, 695–704. [Google Scholar] [CrossRef]
- Arnold, F.; Schnell, J.; Zirafi, O.; Sturzel, C.; Meier, C.; Weil, T.; Standker, L.; Forssmann, W.-G.; Roan, N.R.; Greene, W.C.; et al. Naturally occurring fragments from two distinct regions of the prostatic acid phosphatase form amyloidogenic enhancers of HIV infection. J. Virol. 2012, 86, 1244–1249. [Google Scholar] [CrossRef]
- Roan, N.R.; Muller, J.A.; Liu, H.; Chu, S.; Arnold, F.; Sturzel, C.M.; Walther, P.; Dong, M.; Witkowska, H.E.; Kirchhoff, F.; et al. Peptides released by physiological cleavage of semen coagulum proteins form amyloids that enhance HIV infection. Cell Host Microbe 2011, 10, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Sheik, D.A.; Brooks, L.; Frantzen, K.; Dewhurst, S.; Yang, J. Inhibition of the enhancement of infection of human immunodeficiency virus by semen-derived enhancer of virus infection using amyloid-targeting polymeric nanoparticles. ACS Nano 2015, 9, 1829–1836. [Google Scholar] [CrossRef] [PubMed]
- Sheik, D.A.; Chamberlain, J.M.; Brooks, L.; Clark, M.; Kim, Y.H.; Leriche, G.; Kubiak, C.P.; Dewhurst, S.; Yang, J. Hydrophobic Nanoparticles Reduce the beta-Sheet Content of SEVI Amyloid Fibrils and Inhibit SEVI-Enhanced HIV Infectivity. Langmuir 2017, 33, 2596–2602. [Google Scholar] [CrossRef] [PubMed]
- Haj, E.; Losev, Y.; Guru KrishnaKumar, V.; Pichinuk, E.; Engel, H.; Raveh, A.; Gazit, E.; Segal, D. Integrating in vitro and in silico approaches to evaluate the “dual functionality” of palmatine chloride in inhibiting and disassembling Tau-derived VQIVYK peptide fibrils. Biochim. Biophys. Acta 2018, 1862, 1565–1575. [Google Scholar] [CrossRef] [PubMed]
- Habchi, J.; Chia, S.; Limbocker, R.; Mannini, B.; Ahn, M.; Perni, M.; Hansson, O.; Arosio, P.; Kumita, J.R.; Challa, P.K.; et al. Systematic development of small molecules to inhibit specific microscopic steps of Aβ42 aggregation in Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2016, 114, E200–E208. [Google Scholar] [CrossRef] [PubMed]
- Kurnik, M.; Sahin, C.; Andersen, C.B.; Lorenzen, N.; Giehm, L.; Mohammad-Beigi, H.; Jessen, C.M.; Pedersen, J.S.; Christiansen, G.; Petersen, S.V.; et al. Potent alpha-Synuclein Aggregation Inhibitors, Identified by High-Throughput Screening, Mainly Target the Monomeric State. Cell Chem. Biol. 2018. [Google Scholar] [CrossRef]
- Scherzer-Attali, R.; Pellarin, R.; Convertino, M.; Frydman-Marom, A.; Egoz-Matia, N.; Peled, S.; Levy-Sakin, M.; Shalev, D.E.; Caflisch, A.; Gazit, E.; et al. Complete phenotypic recovery of an Alzheimer’s disease model by a quinone-tryptophan hybrid aggregation inhibitor. PLoS ONE 2010, 5, e11101. [Google Scholar] [CrossRef]
- Frenkel-Pinter, M.; Tal, S.; Scherzer-Attali, R.; Abu-Hussien, M.; Alyagor, I.; Eisenbaum, T.; Gazit, E.; Segal, D. Naphthoquinone-Tryptophan Hybrid Inhibits Aggregation of the Tau-Derived Peptide PHF6 and Reduces Neurotoxicity. J. Alzheimer’s Dis. 2016, 51, 165–178. [Google Scholar] [CrossRef]
- KrishnaKumar, V.G.; Paul, A.; Gazit, E.; Segal, D. Mechanistic insights into remodeled Tau-derived PHF6 peptide fibrils by Naphthoquinone-Tryptophan hybrids. Sci. Rep. 2018, 8, 71. [Google Scholar] [CrossRef] [Green Version]
- Scherzer-Attali, R.; Shaltiel-Karyo, R.; Adalist, Y.H.; Segal, D.; Gazit, E. Generic inhibition of amyloidogenic proteins by two naphthoquinone-tryptophan hybrid molecules. Proteins 2012, 80, 1962–1973. [Google Scholar] [CrossRef]
- Groenning, M. Binding mode of Thioflavin T and other molecular probes in the context of amyloid fibrils-current status. J. Chem. Biol. 2009, 3, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Chatani, E.; Yamamoto, N. Recent progress on understanding the mechanisms of amyloid nucleation. Biophys. Rev. 2017, 10, 527–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- French, K.C.; Makhatadze, G.I. Core sequence of PAPf39 amyloid fibrils and mechanism of pH-dependent fibril formation: The role of monomer conformation. Biochemistry 2012, 51, 10127–10136. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, K.R.; French, K.C.; Tzul, F.O.; Makhatadze, G.I.; Lapidus, L.J. Intramolecular diffusion controls aggregation of the PAPf39 peptide. Biophys. Chem. 2016, 216, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Lu, L.; Li, L.; Liu, J.; Oksov, Y.; Lu, H.; Jiang, S.; Liu, S. Polyanionic candidate microbicides accelerate the formation of semen-derived amyloid fibrils to enhance HIV-1 infection. PLoS ONE 2013, 8, e59777. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Lin, T.Y.; Chang, D.; Guo, Z. Thioflavin T as an amyloid dye: Fibril quantification, optimal concentration and effect on aggregation. R. Soc. Open Sci. 2017, 4, 160696. [Google Scholar] [CrossRef] [PubMed]
- Weber, G.; Laurence, D.J. Fluorescent indicators of adsorption in aqueous solution and on the solid phase. Biochem. J. 1954, 56. [Google Scholar]
- Hawe, A.; Sutter, M.; Jiskoot, W. Extrinsic Fluorescent Dyes as Tools for Protein Characterization. Pharm. Res. 2008, 25, 1487–1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goto, Y.; Yagi, H.; Yamaguchi, K.; Chatani, E.; Ban, T. Structure, formation and propagation of amyloid fibrils. Curr. Pharm. Des. 2008, 14, 3205–3218. [Google Scholar] [CrossRef]
- Gazit, E. A possible role for pi-stacking in the self-assembly of amyloid fibrils. FASEB J. 2002, 16, 77–83. [Google Scholar] [CrossRef]
- KrishnaKumar, V.G.; Baweja, L.; Ralhan, K.; Gupta, S. Carbamylation promotes amyloidogenesis and induces structural changes in Tau-core hexapeptide fibrils. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 2590–2604. [Google Scholar] [CrossRef] [PubMed]
- Westermark, G.T.; Johnson, K.H.; Westermark, P. Staining methods for identification of amyloid in tissue. Methods Enzymol. 1999, 309, 3–25. [Google Scholar] [PubMed]
- Ward, S.M.; Himmelstein, D.S.; Lancia, J.K.; Binder, L.I. Tau oligomers and tau toxicity in neurodegenerative disease. Biochem. Soc. Trans. 2012, 40, 667–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sengupta, U.; Nilson, A.N.; Kayed, R. The Role of Amyloid-β Oligomers in Toxicity, Propagation, and Immunotherapy. EBioMedicine 2016, 6, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Flach, K.; Hilbrich, I.; Schiffmann, A.; Gartner, U.; Kruger, M.; Leonhardt, M.; Waschipky, H.; Wick, L.; Arendt, T.; Holzer, M. Tau oligomers impair artificial membrane integrity and cellular viability. J. Biol. Chem. 2012, 287, 43223–43233. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.; Nadimpally, K.C.; Mondal, T.; Thalluri, K.; Mandal, B. Inhibition of Alzheimer’s amyloid-beta peptide aggregation and its disruption by a conformationally restricted alpha/beta hybrid peptide. Chem. Commun. 2015, 51, 2245–2248. [Google Scholar] [CrossRef] [PubMed]
- McLaurin, J.; Chakrabartty, A. Membrane disruption by Alzheimer beta-amyloid peptides mediated through specific binding to either phospholipids or gangliosides. Implications for neurotoxicity. J. Biol. Chem. 1996, 271, 26482–26489. [Google Scholar] [CrossRef] [PubMed]
- Ross, P.D.; Subramanian, S. Thermodynamics of protein association reactions: Forces contributing to stability. Biochemistry 1981, 20, 3096–3102. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-H.; Liu, F.-F.; Dong, X.-Y.; Sun, Y. Thermodynamic analysis of the molecular interactions between amyloid beta-peptide 42 and (−)-epigallocatechin-3-gallate. J. Phys. Chem. B 2010, 114, 11576–11583. [Google Scholar] [CrossRef] [PubMed]
- Popovych, N.; Brender, J.R.; Soong, R.; Vivekanandan, S.; Hartman, K.; Basrur, V.; Macdonald, P.M.; Ramamoorthy, A. Site specific interaction of the polyphenol EGCG with the SEVI amyloid precursor peptide PAP(248-286). J. Phys. Chem. B 2012, 116, 3650–3658. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.; Kalita, S.; Kalita, S.; Sukumar, P.; Mandal, B. Disaggregation of Amylin Aggregate by Novel Conformationally Restricted Aminobenzoic Acid containing α/β and α/γ Hybrid Peptidomimetics. Sci. Rep. 2017, 7, 40095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, T.L.; Day, I.J.; Serpell, L.C. The effect of Alzheimer’s Abeta aggregation state on the permeation of biomimetic lipid vesicles. Langmuir 2010, 26, 17260–17268. [Google Scholar] [CrossRef] [PubMed]
- Nanga, R.P.R.; Brender, J.R.; Vivekanandan, S.; Popovych, N.; Ramamoorthy, A. NMR structure in a membrane environment reveals putative amyloidogenic regions of the SEVI precursor peptide PAP(248-286). J. Am. Chem. Soc. 2009, 131, 17972–17979. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated Docking with Selective Receptor Flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple ligand-protein interaction diagrams for drug discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Sample of the compound reported is available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value | Standard Deviation |
|---|---|---|
| Kd (μM) | 5.94 | 1.26 |
| n | 1.502 | 0.060 |
| ΔH (kJ/mol) | −42.51 | 2.57 |
| ΔS (J/mol·K) | −37.02 | NA |
| Hydrogen Bonds | Hydrophobic Contacts | ||
|---|---|---|---|
| Interacting Atom of Amino Acid Residues | Interacting Atoms of NQTrp | Residues | Region of NQTrp |
| N1 of Lys253 | O4 | Lys251 | Naphthoquinone ring |
| O1 of Glu254 | N2 | Gln252 | Naphthoquinone ring |
| N2 of Lys272 | O2 | Lys255 | Indole ring |
| Leu258 | Indole ring | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Viswanathan, G.K.; Mohapatra, S.; Paul, A.; Arad, E.; Jelinek, R.; Gazit, E.; Segal, D. Inhibitory Effect of Naphthoquinone-Tryptophan Hybrid towards Aggregation of PAP f39 Semen Amyloid. Molecules 2018, 23, 3279. https://doi.org/10.3390/molecules23123279
Viswanathan GK, Mohapatra S, Paul A, Arad E, Jelinek R, Gazit E, Segal D. Inhibitory Effect of Naphthoquinone-Tryptophan Hybrid towards Aggregation of PAP f39 Semen Amyloid. Molecules. 2018; 23(12):3279. https://doi.org/10.3390/molecules23123279
Chicago/Turabian StyleViswanathan, Guru KrishnaKumar, Satabdee Mohapatra, Ashim Paul, Elad Arad, Raz Jelinek, Ehud Gazit, and Daniel Segal. 2018. "Inhibitory Effect of Naphthoquinone-Tryptophan Hybrid towards Aggregation of PAP f39 Semen Amyloid" Molecules 23, no. 12: 3279. https://doi.org/10.3390/molecules23123279
APA StyleViswanathan, G. K., Mohapatra, S., Paul, A., Arad, E., Jelinek, R., Gazit, E., & Segal, D. (2018). Inhibitory Effect of Naphthoquinone-Tryptophan Hybrid towards Aggregation of PAP f39 Semen Amyloid. Molecules, 23(12), 3279. https://doi.org/10.3390/molecules23123279