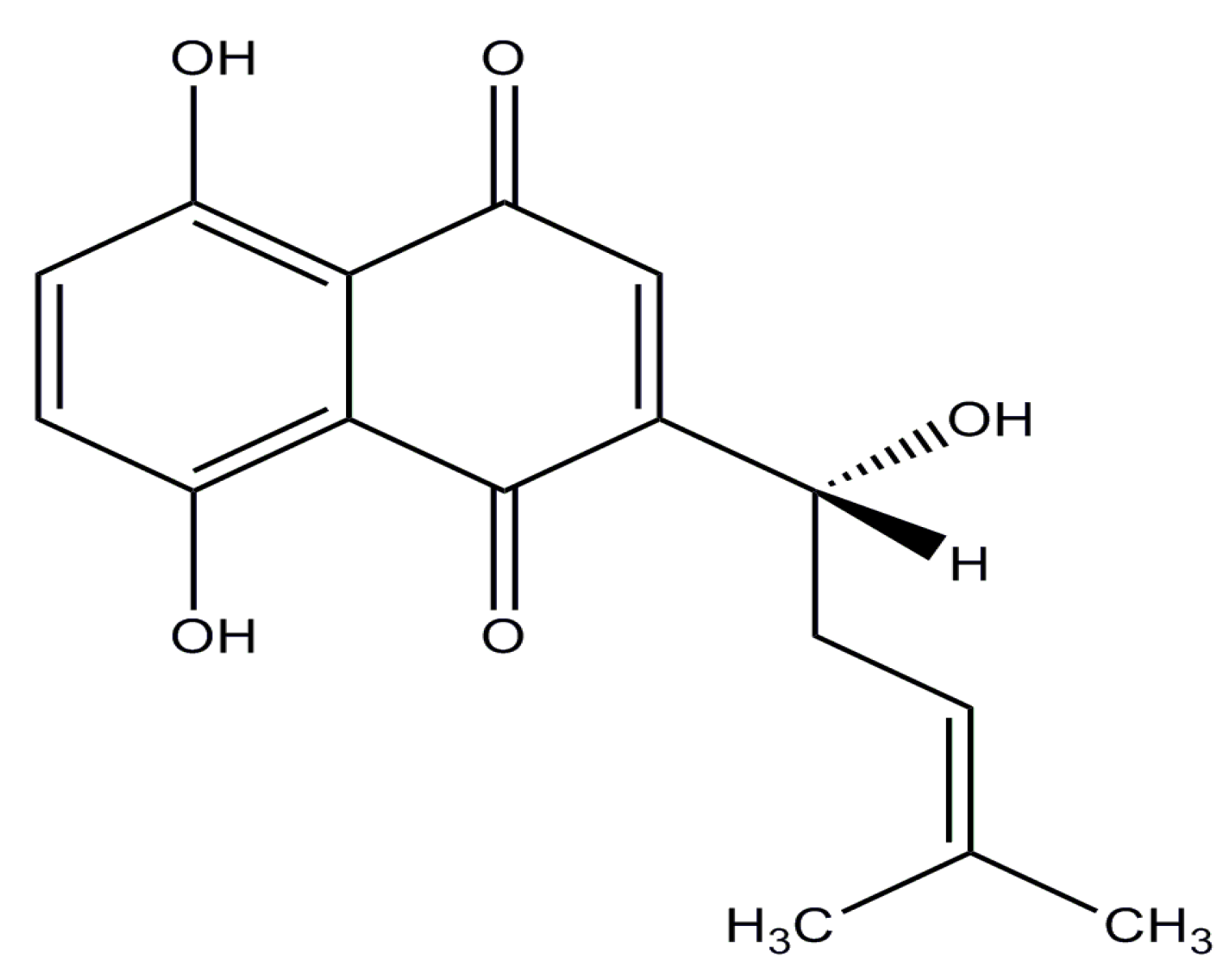
Anticancer Efficacy of Targeted Shikonin Liposomes Modified with RGD in Breast Cancer Cells

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
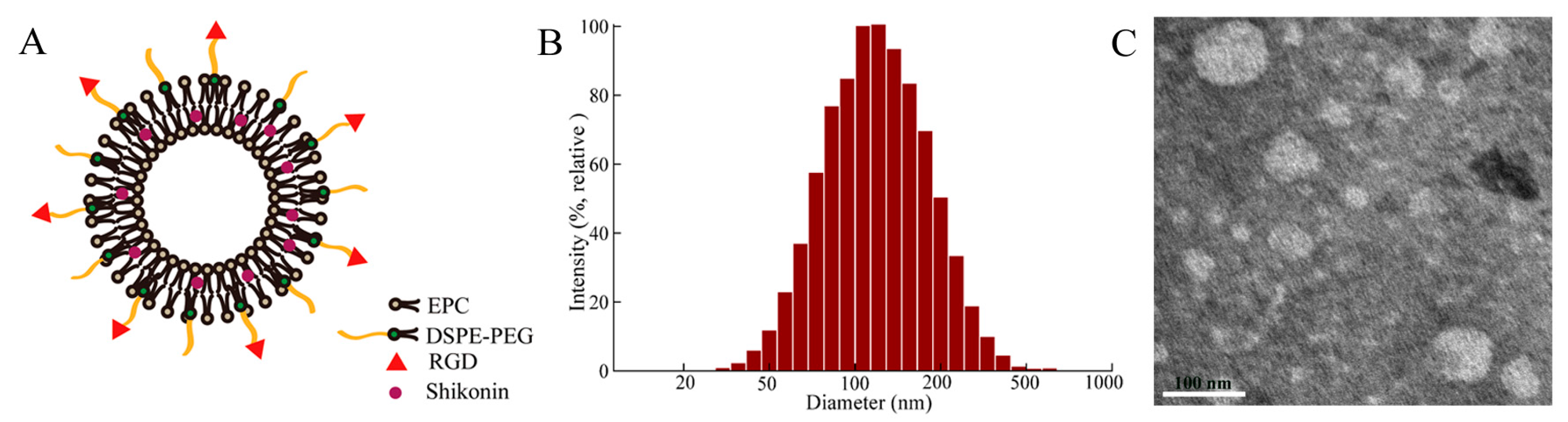
2.1. Preparation and Characterization of Liposomes
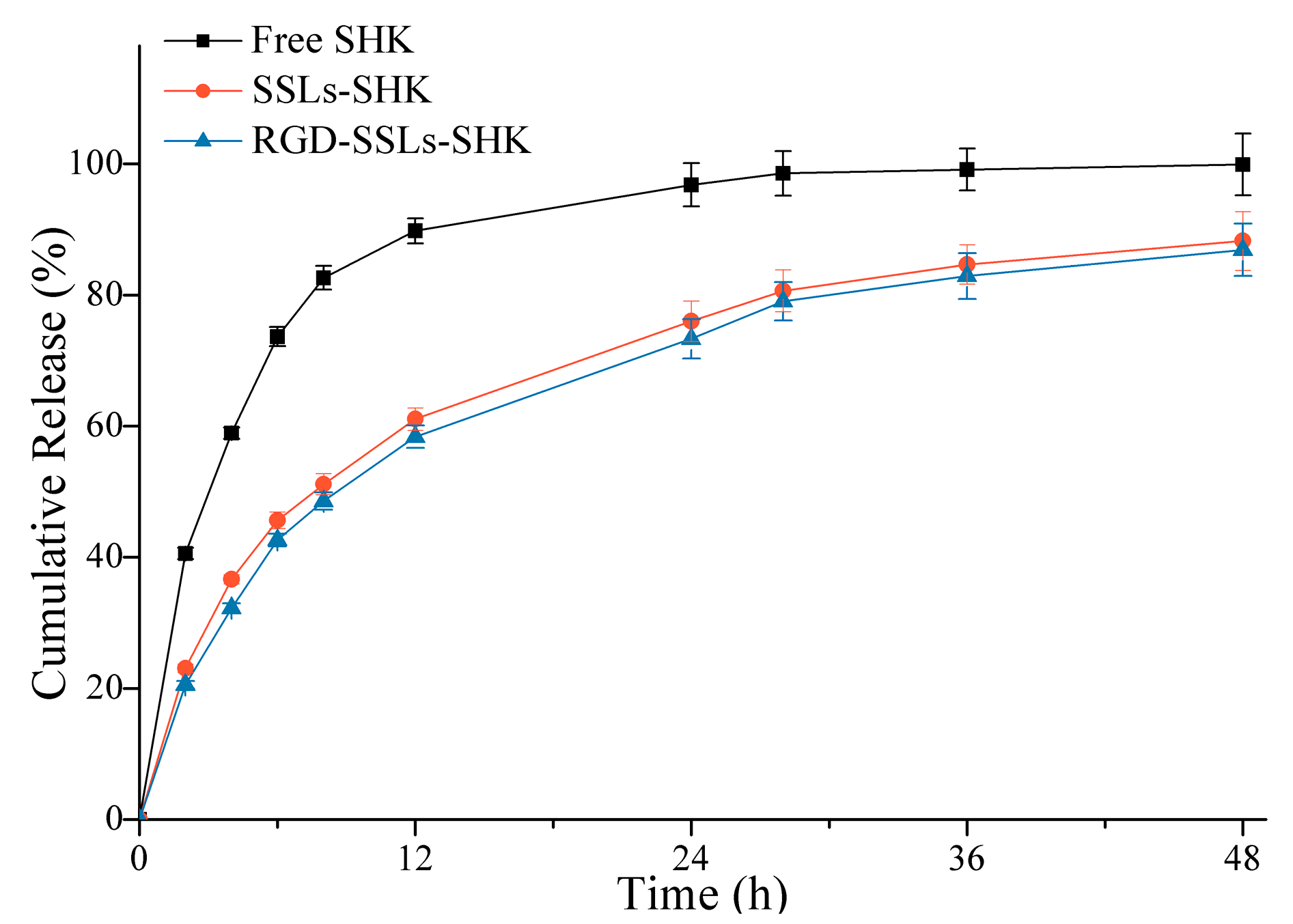
2.2. In Vitro Release Behavior of Liposomes
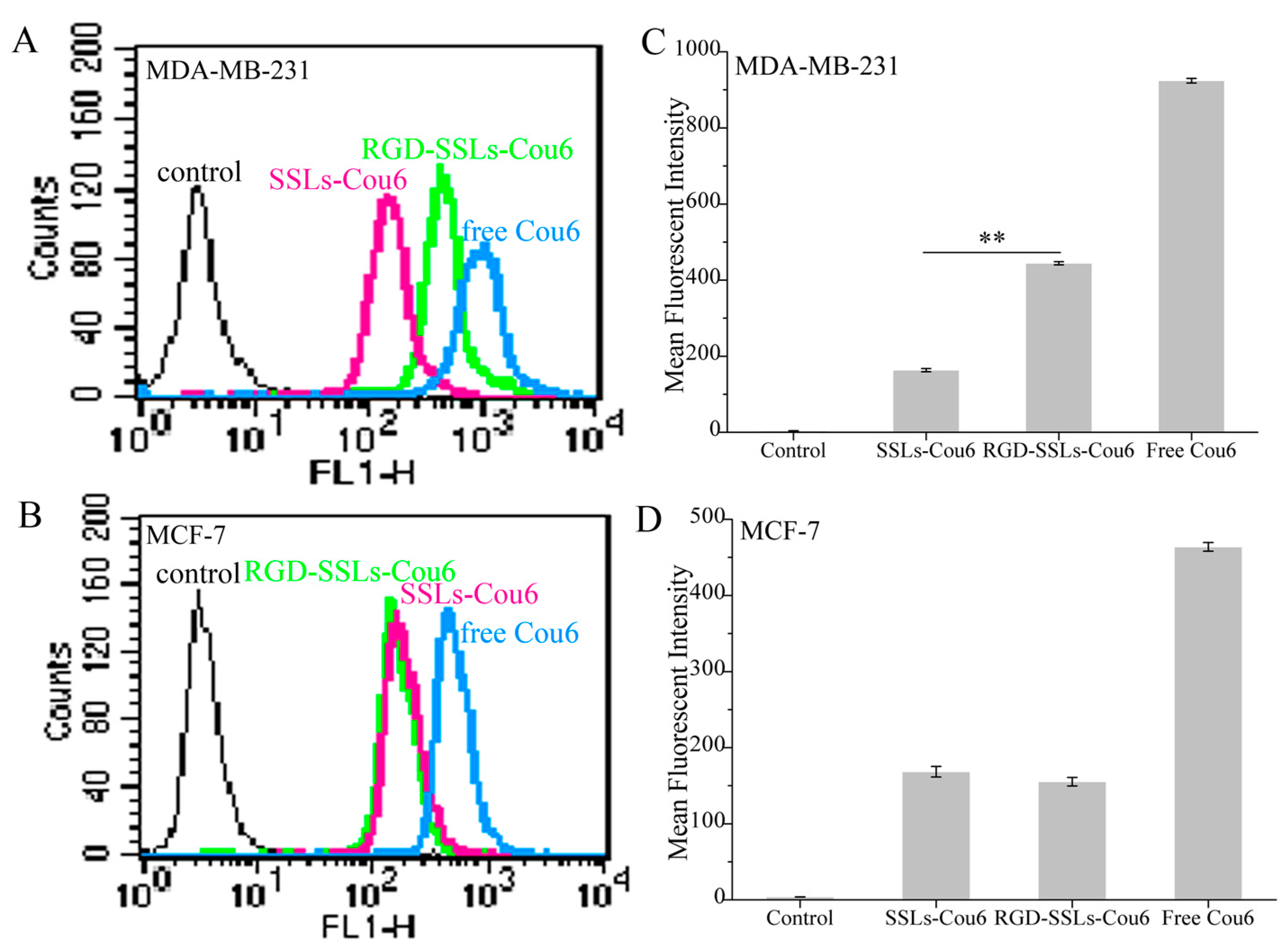
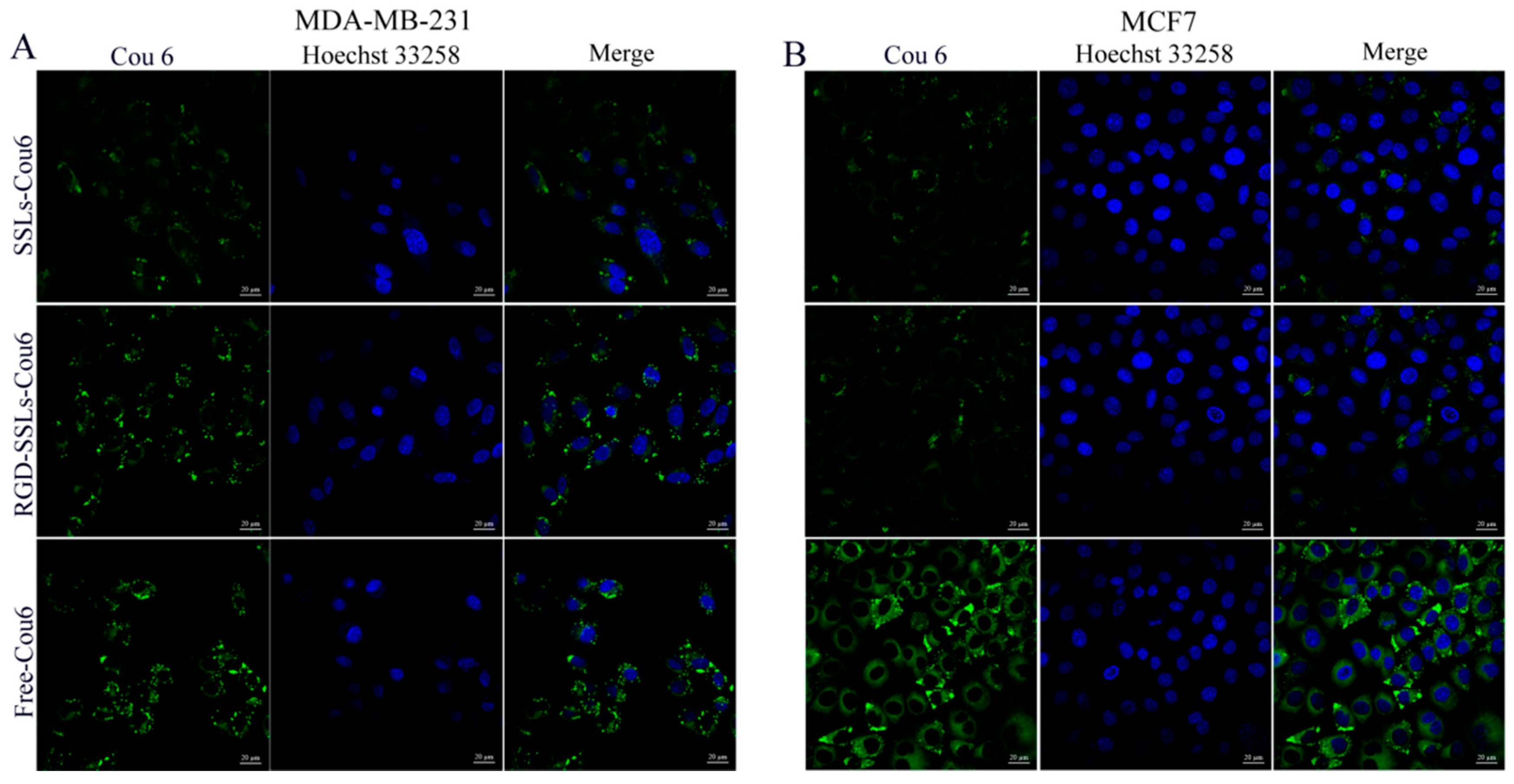
2.3. In Vitro Cellular Uptake
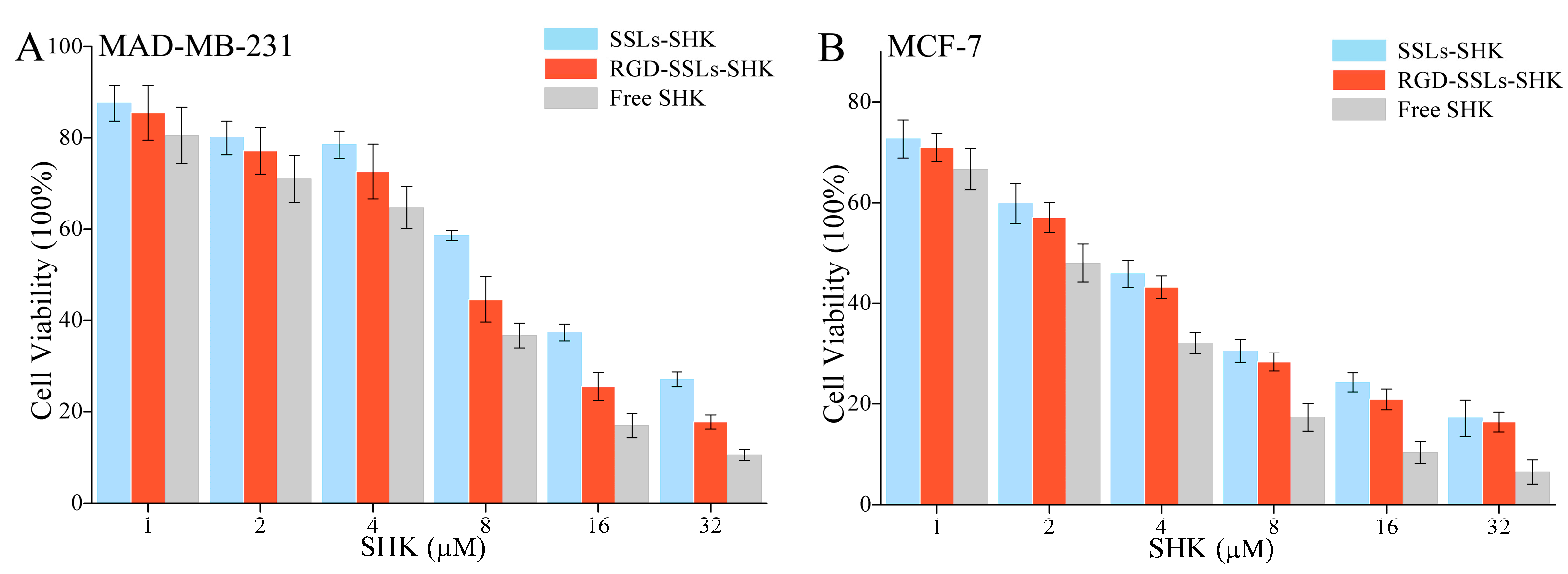
2.4. In Vitro Cytotoxicity Investigation
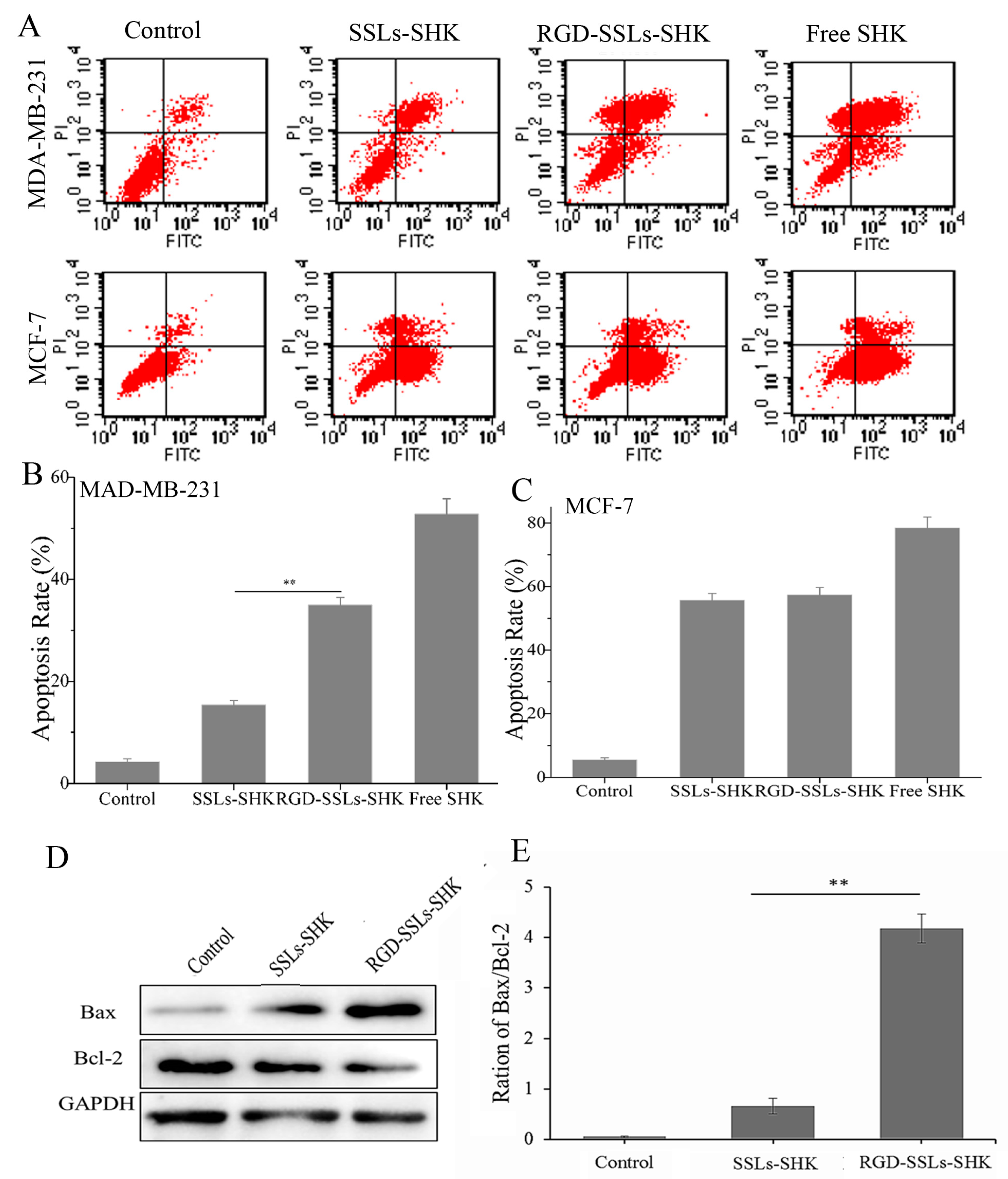
2.5. In Vitro Apoptosis-Promoting Effect of RGD-SSLs-SHK
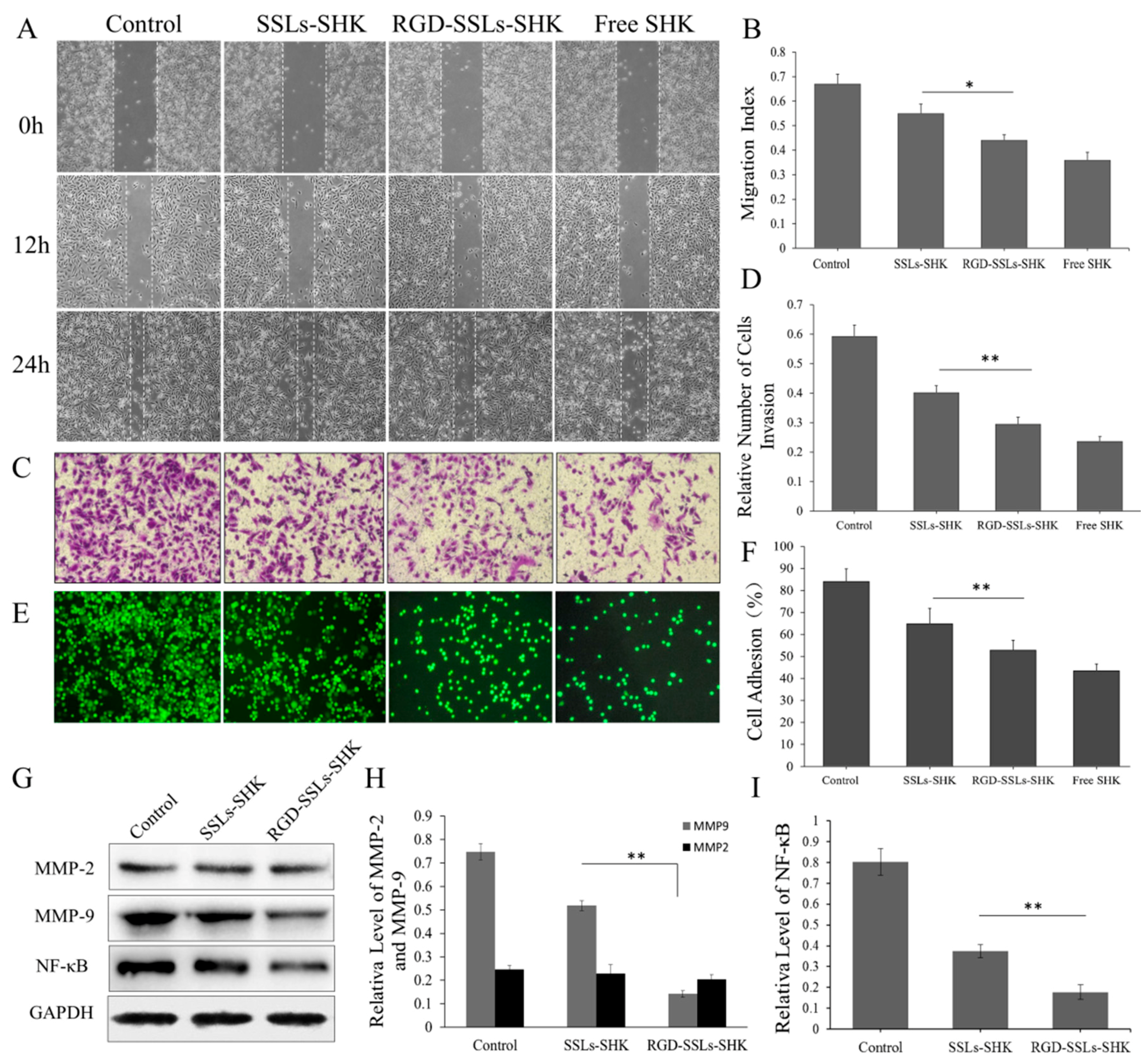
2.6. In Vitro Inhibition Effects of RGD-SSLs-SHK on Migration, Invasion, and Adhesion
3. Materials and Methods
3.1. Materials
3.2. Preparation of Liposomes
3.3. Characterization of Liposomes
3.4. Cell Culture
3.5. In Vitro Release of SHK from Liposomes
3.6. In Vitro Cellular Uptake
3.7. In Vitro Cytotoxicity of Liposomes Assay
3.8. In Vitro Apoptosis Measurement
3.9. Cell Migration Assay
3.10. Transwell Invasion Assay
3.11. Cell–Matrigel Adhesion Assay
3.12. In Vivo Western Blotting Analysis
3.13. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Deshpande, P.P.; Biswas, S.; Torchilin, V.P. Current trends in the use of liposomes for tumor targeting. Nanomedicine 2013, 8, 1509–1528. [Google Scholar] [CrossRef] [PubMed]
- Tu, C.F.; Wu, M.Y.; Lin, Y.C.; Kannagi, R.; Yang, R.B. FUT8 promotes breast cancer cell invasiveness by remodeling TGF-β receptor core fucosylation. Breast Cancer Res. 2017, 19, 111–126. [Google Scholar] [CrossRef] [PubMed]
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Graham, E.G.; Wailes, E.M.; Levi-Polyachenko, N.H. Multi-walled carbon nanotubes inhibit breast cancer cell migration. J. Biomed. Nanotechnol. 2015, 12, 308–319. [Google Scholar] [CrossRef]
- Wu, Y.; Yu, X.; Yi, X.; Wu, K.; Dwabe, S.; Atefi, M.; Elshimali, Y.; Kemp, K.T.; Bhat, K.; Haro, J.; et al. Aberrant phosphorylation of SMAD4 Thr 277-mediated USP9x-SMAD4 interaction by free fatty acids promotes breast cancer metastasis. Cancer Res. 2017, 77, 1383–1394. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.; Qian, J.; Yang, J. Shikonin induces apoptosis and inhibits migration of ovarian carcinoma cells by inhibiting the phosphorylation of Src and FAK. Oncol. Lett. 2015, 9, 629–633. [Google Scholar] [CrossRef] [PubMed]
- Balca-Silva, J.; Matias, D.; do Carmo, A.; Girão, H.; Moura-Neto, V.; Sarmento-Ribeiro, A.B.; Lopes, M.C. Tamoxifen in combination with temozolomide induce a synergistic inhibition of PKC-pan in GBM cell lines. Biochim. Biophys. Acta 2015, 1850, 722–732. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Hwang, K.E.; Park, D.S.; Oh, S.H.; Jun, H.Y.; Yoon, K.H.; Jeong, E.T.; Kim, H.R.; Kim, Y.S. Shikonin induced necroptosis is enhanced by the inhibition of autophagy in non small cell lung cancer cells. J. Transl. Med. 2017, 15, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Li, M.; Cui, S.; Wang, D.; Zhang, C.Y.; Zen, K.; Li, L. Shikonin Inhibits the proliferation of human breast cancer cells by reducing tumor-derived exosomes. Molecules 2016, 21, 777. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Brodie, A.M.; Davidson, N.E.; Kensler, T.W.; Zhou, Q. Inhibition of estrogen signaling activates the NRF2 pathway in breast cancer. Breast Cancer Res. Treat. 2010, 124, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Hong, D.; Jang, S.Y.; Jan, E.H.; Jung, B.; Cho, I.H.; Park, M.J.; Jeong, S.Y.; Kim, J.H. Shikonin as an inhibitor of the LPS-induced epithelial-to-mesenchymal transition in human breast cancer cells. Int. J. Mol. Med. 2015, 36, 1601–1606. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.Y.; Lee, J.K.; Jang, E.H.; Jeong, S.Y.; Kim, J.H. Shikonin blocks migration and invasion of human breast cancer cells through inhibition of matrix metalloproteinase-9 activation. Oncol. Rep. 2014, 31, 2827–2833. [Google Scholar] [CrossRef] [PubMed]
- Shahsavari, Z.; Karamitehrani, F.; Salami, S.; Ghasemzadeh, M. RIP1K and RIP3K provoked by shikonin induce cell cycle arrest in the triple negative breast cancer cell line, MDA-MB-468: Necroptosis as a desperate programmed suicide pathway. Tumor Biol. 2016, 37, 4479–4491. [Google Scholar] [CrossRef] [PubMed]
- Shahsavari, Z.; Karamitehrani, F.; Salami, S. Shikonin Induced necroptosis via reactive oxygen species in the T-47D breast cancer cell line. Asian Pac. J. Cancer Prev. 2015, 16, 7261–7266. [Google Scholar] [CrossRef] [PubMed]
- Thakur, R.; Trivedi, R.; Rastogi, N.; Singh, M.; Mishra, D.P. Inhibition of STAT3, FAK and Src mediated signaling reduces cancer stem cell load, tumorigenic potential and metastasis in breast cancer. Sci. Rep. 2015, 5, 10194. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Huang, N.; Chen, D.; Zhang, L.; Dong, X.; Sun, Y.; Zhu, X.; Zhang, F.; Gao, J.; Wang, Y.; et al. Successful in vivo hyperthermal therapy toward breast cancer by Chinese medicine shikonin-loaded thermosensitive micelle. Int. J. Nanomed. 2017, 12, 4019–4035. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Lin, Y.; Zhang, X.; Feng, C.; Lu, Y.; Gao, Y.; Dong, C. Cyclic RGD peptide-modified liposomal drug delivery system for targeted oral apatinib administration: enhanced cellular uptake and improved therapeutic effects. Int. J. Nanomed. 2017, 12, 1941–1958. [Google Scholar] [CrossRef] [PubMed]
- Veneti, E.; Tu, R.S.; Auguste, D.T. RGD-Targeted Liposome Binding and Uptake on Breast Cancer Cells Is Dependent on Elastin Linker Secondary Structure. Bioconjug. Chem. 2016, 27, 1813–1821. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Matsumura, Y. EPR effect based drug design and clinical outlook for enhanced cancer chemotherapy. Adv. Drug Deliv. Rev. 2011, 63, 129–130. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, G.; Gao, M.; Liu, X.; Ji, B.; Hua, R.; Zhou, Y.; Yang, Y. RGD-peptide conjugated inulin-ibuprofen nanoparticles for targeted delivery of Epirubicin. Colloids Surf. B Biointerfaces 2010, 148, 135–146. [Google Scholar] [CrossRef]
- Brooks, P.C.; Clark, R.A.; Cheresh, D.A. Requirement of vascular integrin alphav-beta3 for angiogenesis. Science 1994, 264, 569–571. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Zheng, J.H.; Nguyen, V.H.; Jiang, S.N.; Kim, D.Y.; Szardenings, M.; Min, J.H.; Hong, Y.J.; Choy, H.E.; Min, J.J. RGD Peptide cell-surface display enhances the targeting and therapeutic efficacy of attenuate salmonella-mediated cancer therapy. Theranostics 2016, 6, 1672–1682. [Google Scholar] [CrossRef] [PubMed]
- Fei, W.; Zhang, Y.; Han, S.; Tao, J.; Zheng, H.; Wei, Y.; Zhu, J.; Li, F.; Wang, X. RGD conjugated liposome-hollow silica hybrid nanovehicles for targeted and controlled delivery of arsenic trioxide against hepatic carcinoma. Int. J. Pharm. 2017, 519, 250–262. [Google Scholar] [CrossRef] [PubMed]
- Fang, I.J.; Slowing, I.I.; Wu, K.C.; Lin, V.S.; Trewyn, B.G. Ligand conformation dictates membrane and endosomal trafficking of arginine-glycine-aspartate (RGD)-functionalized mesoporous silica nanoparticles. Chemistry 2012, 18, 7787–7792. [Google Scholar] [CrossRef] [PubMed]
- Shan, D.; Li, J.; Cai, P.; Prasad, P.; Liu, F.; Rauth, A.M.; Wu, X.Y. RGD-conjugated solid lipid nanoparticles inhibit adhesion and invasion of αvβ3 integrin-overexpressing breast cancer cells. Drug Deliv. Transl. Res. 2015, 5, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.B.; Huang, Y.; Lu, W.L.; Zhang, X.; Zhang, H.; Nagai, T.; Zhang, Q. Intracellular delivery of doxorubicin with RGD-modified sterically stabilized liposomes for an improved antitumor efficacy: In vitro and in vivo. J. Pharm. Sci. 2005, 94, 1782–1793. [Google Scholar] [CrossRef] [PubMed]
- Allen, T.M.; Cullis, P.R. Liposomal drug delivery systems: From concept to clinical applications. Adv. Drug Deliv. Rev. 2013, 65, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chen, Y.; Cheng, Y.; Gao, Y. Glycyrrhetinic acid liposomes containing mannose-diester lauric diacid-cholesterol conjugate synthesized by lipase-catalytic acylation for liver-specific delivery. Molecules 2017, 22, 1598. [Google Scholar] [CrossRef] [PubMed]
- Lai, H.; Zhao, Z.; Li, L.; Zheng, W.; Chen, T. Antiangiogenic ruthenium (ii) benzimidazole complexes, structure-based activation of distinct signaling pathways. Metallomics 2015, 7, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Düzgüneş, N.; Nir, S. Mechanisms and kinetics of liposome-cell interactions. Adv. Drug Deliv. Rev. 1999, 40, 3–18. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, Y.; Li, L.; Zhang, Y.; Lu, Y.; Jiang, X.; He, X.; Ma, H.; An, S.; Jiang, C. Cell Microenvironment-Controlled Antitumor Drug Releasing-Nanomicelles for GLUT1 Targeting Hepatocellular Carcinoma Therapy. ACS Appl. Mater Interfaces 2015, 7, 5444–5453. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.; Su, B.; Li, W.; Ding, Y.; Tang, L.; Zhou, W.; Song, Y.; Li, H.; Zhou, C. Enhanced antitumor effect of novel dual targeted paclitaxel liposomes. Nanotechnology 2010, 21, 415103. [Google Scholar] [CrossRef] [PubMed]
- Hematpoor, A.; Paydar, M.; Liew, S.Y.; Sivasothy, Y.; Mohebali, N.; Looi, C.Y.; Wong, W.F.; Azirun, M.S.; Awang, K. Phenylpropanoids isolated from Piper sarmentosum Roxb. induce apoptosis in breast cancer cells through reactive oxygen species and mitochondrial-dependent pathways. Chem. Biol. Interact. 2017, 22, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Del, P.G.; Venditti, A.; Principe, M.D.; Maurillo, L.; Buccisano, F.; Tamburini, A.; Cox, M.C.; Franchi, A.; Bruno, A.; Mazzone, C.; et al. Amount of spontaneous apoptosis detected by Bax/Bcl-2 ratio predicts outcome in acute myeloid leukemia (AML). Blood 2003, 101, 2125–2131. [Google Scholar]
- Ennen, M.; Klotz, R.; Touche, N.; Pinel, S.; Barbieux, C.; Besancenot, V.; Brunner, E.; Thiebaut, D.; Jung, A.C.; Ledrappier, S.; et al. DDB2: A novel regulator of NF-κB and breast tumor invasion. Cancer Res. 2013, 73, 5040–5052. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cui, L.; Feng, L.; Zhang, Z.; Song, J.; Liu, D.; Jia, X. Isoalantolactone inhibits the migration and invasion of human breast cancer MDA-MB-231 cells via suppression of the p38 MAPK/NF-κB signaling pathway. Oncol. Rep. 2016, 36, 1269–1276. [Google Scholar] [CrossRef] [PubMed]
- Sarnico, I.; Lanzillotta, A.; Boroni, F.; Benarese, M.; Alghisi, M.; Schwaninger, M.; Inta, I.; Battistin, L.; Spano, P.; Pizzi, M. NF-kappaB p50/RelA and c-Rel-containing dimers: Opposite regulators of neuron vulnerability to ischaemia. J. Neurochem. 2009, 8, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Cai, F.; Liu, X.; Guo, L. Sesamin inhibits lipopolysaccharide-induced proliferation and invasion through the p38-MAPK and NF-κB signaling pathways in prostate cancer cells. Oncol. Rep. 2015, 33, 3117–3123. [Google Scholar] [CrossRef] [PubMed]
- Gubernator, J. Active methods of drug loading into liposomes: recent strategies for stable drug entrapment and increased in vivo activity. Expert Opin. Drug Deliv. 2011, 8, 565–580. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Li, L.; He, X.; Yi, Q.; He, B.; Cao, J.; Pan, W.; Gu, Z. Overcoming drug resistant lung cancer by paclitaxel loaded dual-functional liposomes with mitochondria targeting and pH-response. Biomaterials 2015, 52, 126–139. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Chen, T.; You, Y.; Hu, H.; Zheng, W.; Kwong, W.L.; Zou, T.; Che, C.M. A cancer-targeted nanosystem for delivery of gold (III) complexes: Enhanced selectivity and apoptosis-inducing efficacy of a gold(III) porphyrin complex. Angew. Chem. Int. Ed. Engl. 2014, 53, 12532–12536. [Google Scholar] [PubMed]
- Liang, L.; Gao, C.; Li, Y.; Sun, M.; Xu, J.; Li, H.; Jia, L.; Zhao, Y. MiR-125a-3p/FUT5-FUT6 axis mediates colorectal cancer cell proliferation, migration, invasion and pathological angiogenesis via PI3K-Akt pathway. Cell Death Dis. 2017, 8, e2968. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Bower, K.A.; Wang, S.; Frank, J.A.; Chen, G.; Ding, M.; Wang, S.; Shi, S.; Ke, Z.; Luo, J. Cyanidin-3-glucoside inhibits ethanol induced invasion of breast cancer cells overexpressing ErbB2. Mol. Cancer 2010, 9, 285. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Liposomes | Size (nm) | Zeta Potential (mV) | PDI | EE (%) |
|---|---|---|---|---|
| SSLs-SHK | 121.36 ± 2.89 | −16.32 ± 1.45 | 0.208 ± 0.021 | 92.51 ± 2.55 |
| SSLs-Cou6 | 119.96 ± 3.41 | −16.01 ± 1.68 | 0.213 ± 0.023 | 92.71 ± 2.91 |
| RGD-SSLs-SHK | 117.53 ± 3.05 | −15.37 ± 0.91 | 0.180 ± 0.015 | 94.89 ± 1.83 |
| RGD-SSLs-Cou6 | 120.67 ± 2.63 | −15.86 ± 0.93 | 0.201 ± 0.014 | 93.84 ± 1.41 |
| Liposomes | IC50 (µM) in MDA-MB-231 Cells | IC50 (µM) in MCF-7 Cells |
|---|---|---|
| SSLs-SHK | 10.92 ± 1.03 ∆ | 3.34 ± 0.18 ∆ |
| RGD-SSLs-SHK | 7.16 ± 0.62 ∆ ** | 2.96 ± 0.12 ∆ |
| free SHK | 4.92 ± 0.29 | 1.90 ± 0.11 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, X.; Li, J.; Cai, D.; Yue, L.; Wang, Q.; Zhou, L.; Fan, L.; Sun, J.; Wu, Y. Anticancer Efficacy of Targeted Shikonin Liposomes Modified with RGD in Breast Cancer Cells. Molecules 2018, 23, 268. https://doi.org/10.3390/molecules23020268
Wen X, Li J, Cai D, Yue L, Wang Q, Zhou L, Fan L, Sun J, Wu Y. Anticancer Efficacy of Targeted Shikonin Liposomes Modified with RGD in Breast Cancer Cells. Molecules. 2018; 23(2):268. https://doi.org/10.3390/molecules23020268
Chicago/Turabian StyleWen, Xianchun, Jiping Li, Defu Cai, Liling Yue, Qi Wang, Li Zhou, Li Fan, Jianwen Sun, and Yonghui Wu. 2018. "Anticancer Efficacy of Targeted Shikonin Liposomes Modified with RGD in Breast Cancer Cells" Molecules 23, no. 2: 268. https://doi.org/10.3390/molecules23020268
APA StyleWen, X., Li, J., Cai, D., Yue, L., Wang, Q., Zhou, L., Fan, L., Sun, J., & Wu, Y. (2018). Anticancer Efficacy of Targeted Shikonin Liposomes Modified with RGD in Breast Cancer Cells. Molecules, 23(2), 268. https://doi.org/10.3390/molecules23020268