Identification, Characterization, and Expression Analysis of a FMRFamide-Like Peptide Gene in the Common Chinese Cuttlefish (Sepiella japonica)



Abstract
:1. Introduction
2. Results and Discussion
2.1. Whole-Length Sequence Analysis and Protein Structural Prediction
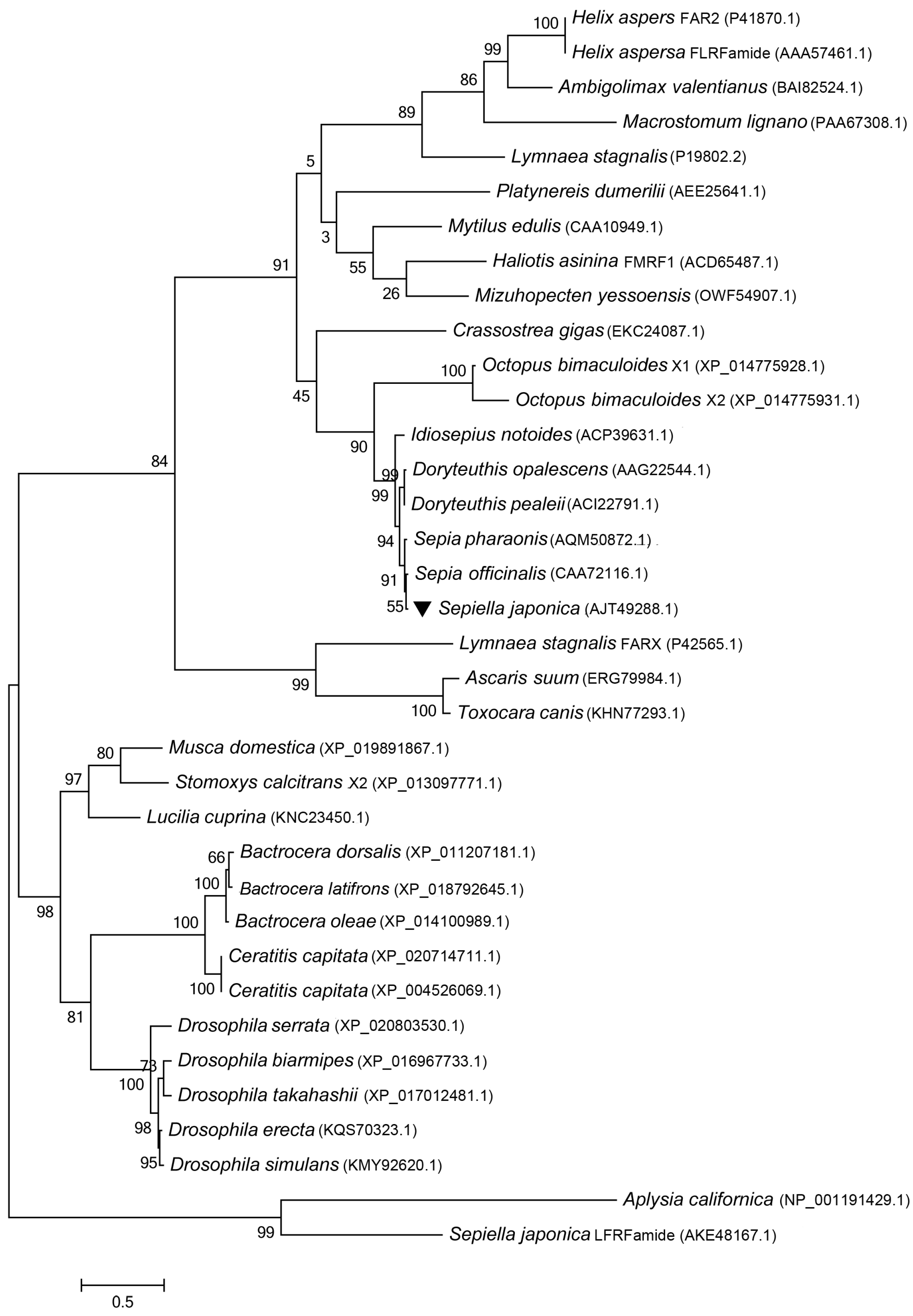
2.2. A Blast Search and Phylogenetic-Tree Construction
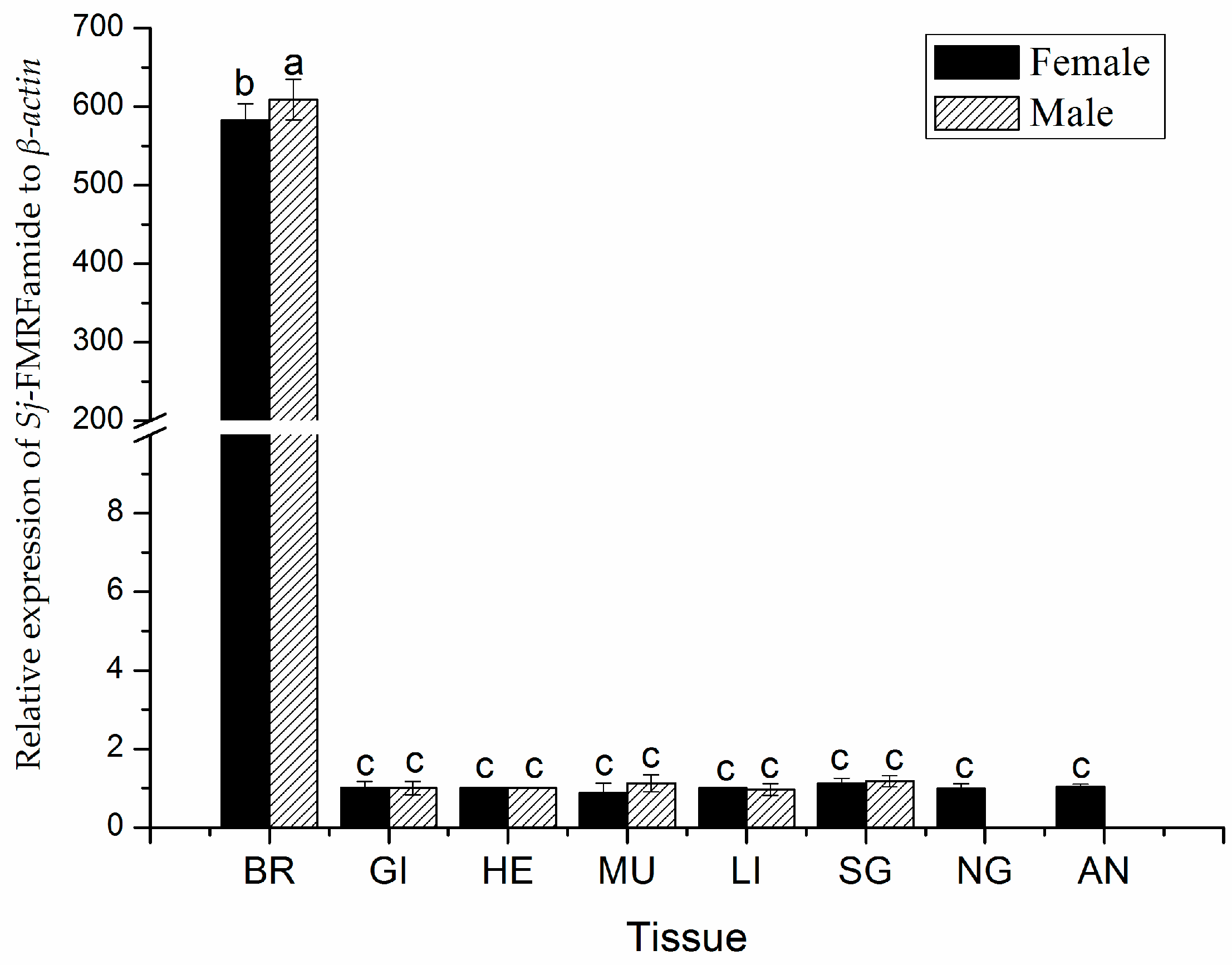
2.3. The Expression Pattern of SjFMRFamide mRNA in Different Tissues.
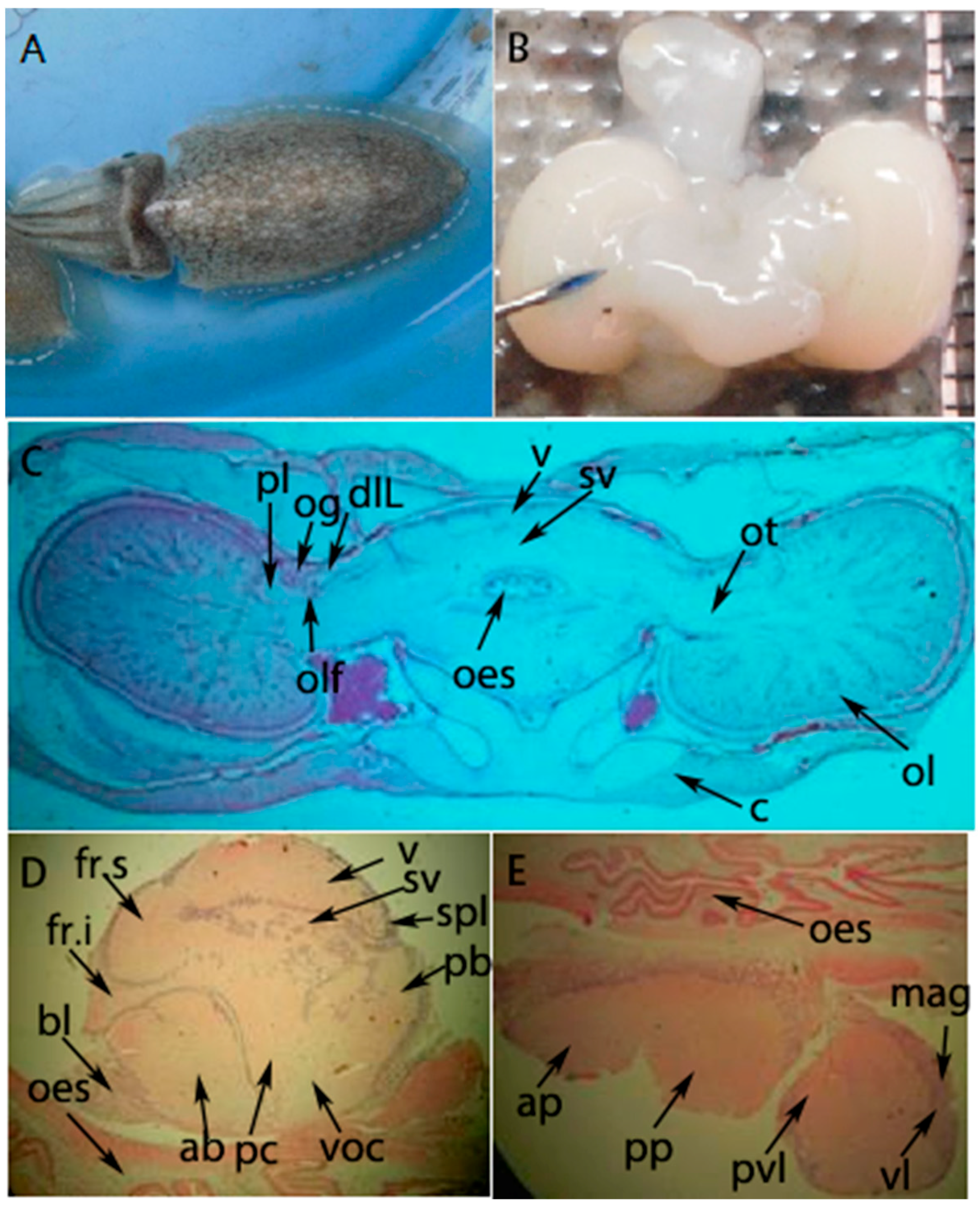
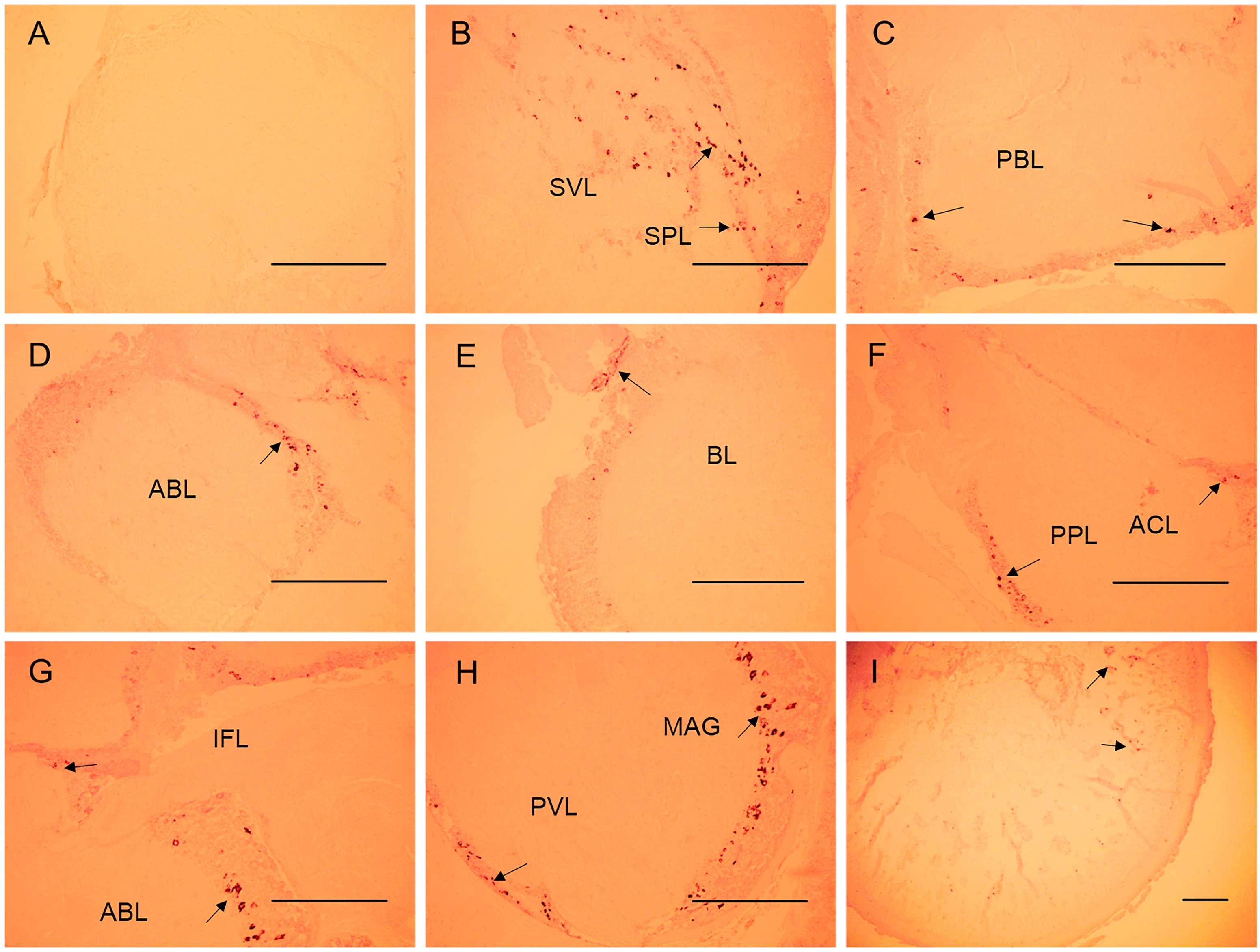
2.4. In Situ Hybridization of the SjFMRFamide mRNA for Expression Analysis
3. Experimental Section
3.1. Animals
3.2. RNA Extraction and cDNA Full-Length Amplification
3.3. Identification and Whole-Length Amplification of cDNA of SjFMRFamide
3.4. Bioinformatics Analysis of SjFMRFamide
3.5. qRT-PCR of SjFMRFamide in Different Tissue
3.6. The In Situ Hybridization Assay of SjFMRFamide
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Price, D.A.; Greenberg, M.J. Purification and characterization of a cardioexcitatory neuropeptide from the central ganglia of a bivalve mollusc. Prep. Biochem. 1977, 7, 261–281. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, E.; Carrigan, M.; Thomas, S.G.; Shaw, G.; Edison, A.S. A statistical view of FMRFamide neuropeptide diversity. Mol. Neurobiol. 2000, 21, 35–56. [Google Scholar] [PubMed]
- Pinelli, C.; D’Aniello, B.; Sordino, P.; Meyer, D.L.; Fiorentino, M.; Rastogi, R.K. Comparative immunocytochemical study of FMRFamide neuronal system in the brain of Danio rerio and Acipenser ruthenus during development. Brain Res. Dev. Brain Res. 2000, 119, 195–208. [Google Scholar] [CrossRef]
- Ons, S.; Sterkel, M.; Diambra, L.; Urlaub, H.; Rivera-Pomar, R. Neuropeptide precursor gene discovery in the Chagas disease vector Rhodnius prolixus. Insect Mol. Biol. 2011, 20, 29–44. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Vera, E.; Aguilar, M.B.; Heimer de la Cotera, E.P. FMRFamide and related peptides in the phylum mollusca. Peptides 2008, 29, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Henry, J.; Zatylny, C.E. Peptidergic control of egg-laying in the cephalopod Sepia officinalis: Involvement of FMRFamide and FMRFamide-related peptides. Peptides 1999, 20, 1061–1070. [Google Scholar] [CrossRef]
- Walker, R.J.; Papaioannou, S.; Holdendye, L. A review of FMRFamide- and RFamide-like peptides in metazoa. Invertebr. Neurosci. 2009, 9, 111–153. [Google Scholar] [CrossRef] [PubMed]
- Zatylnygaudin, C.; Favrel, P. Diversity of the RFamide Peptide Family in Mollusks. Front. Endocrinol. 2014, 5, 178. [Google Scholar]
- Cottrell, G.A.; Price, D.A.; Greenberg, M.J. FMRFamide-like activity in the ganglia and in a single identified neurone of Helix aspersa. Comp. Biochem. Physiol. C 1981, 70, 103–107. [Google Scholar] [CrossRef]
- Stefano, G.B.; Martin, R. Enkephalin-like immunoreactivity in the pedal ganglion of Mytilus edulis (Bivalvia) and its proximity to dopamine-containing structures. Cell Tissue Res. 1983, 230, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Too, C.K.; Croll, R.P. Detection of FMRFamide-like immunoreactivities in the sea scallop Placopecten magellanicus by immunohistochemistry and Western blot analysis. Cell Tissue Res. 1995, 281, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Le, G.S.; Féral, C.; Van, M.J.; Marchand, C.R. Evidence for peptidergic innervation of the endocrine optic gland in Sepia by neurons showing FMRFamide-like immunoreactivity. Brain Res. 1988, 462, 83–88. [Google Scholar]
- Cosmo, A.D.; Cristo, C.D. Neuropeptidergic control of the optic gland of Octopus vulgaris: FMRF-amide and GnRH immunoreactivity. J. Comp. Neurol. 1998, 398, 1–12. [Google Scholar] [CrossRef]
- Loi, P.; Saunders, R.; Young, D.; Tublitz, N. Peptidergic regulation of chromatophore function in the European cuttlefish Sepia officinalis. J. Exp. Biol. 1996, 199, 1177–1187. [Google Scholar] [PubMed]
- Tublitz, N.J.; Gaston, M.R.; Loi, P.K. Neural regulation of a complex behavior: body patterning in cephalopod molluscs. Integr. Comp. Biol. 2006, 46, 880–889. [Google Scholar] [CrossRef] [PubMed]
- Di, C.C.; Delli, B.P.; Di, C.A. Role of FMRFamide in the reproduction of Octopus vulgaris: Molecular analysis and effect on visual input. Peptides 2003, 24, 1525–1532. [Google Scholar]
- Cao, Z.H.; Sun, L.L.; Chi, C.F.; Liu, H.H.; Zhou, L.Q.; Lv, Z.M.; Wu, C.W. Molecular cloning, expression analysis and cellular localization of an LFRFamide gene in the cuttlefish Sepiella japonica. Peptides 2015, 80, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Lin, X.; Wang, Z.; Yu, R.; Tian, C.; Li, Z. Studies on life cycle of cultured spineless cuttlefish Sepiella japonica. Trans. Oceanol. Limnol. 2010, 30, 24–28. [Google Scholar]
- Li, C.; Kim, K.; Nelson, L.S. FMRFamide-related neuropeptide gene family in Caenorhabditis elegans. Brain Res. 1999, 848, 26–34. [Google Scholar] [CrossRef]
- Fisher, T.; Lin, C.H.; Kaczmarek, L.K. The peptide FMRFa terminates a discharge in Aplysia bag cell neurons by modulating calcium, potassium, and chloride conductances. J. Neurophysiol. 1993, 69, 2164–2173. [Google Scholar] [CrossRef] [PubMed]
- Loi, P.K.; Tublitz, N. Molecular analysis of FMRFamide- and FMRFamide-related peptides (FaRPS) in the cuttlefish Sepia officinalis. J. Exp. Biol. 1997, 200, 1483–1489. [Google Scholar] [PubMed]
- Martin, R.; Voigt, K.H. The neurosecretory system of the octopus vena cava: A neurohemal organ. Experientia 1987, 43, 537–543. [Google Scholar] [CrossRef]
- Chin, G.J.; Payza, K.; Price, D.A.; Greenberg, M.J.; Doble, K.E. Characterization and Solubilization of the FMRFamide Receptor of Squid. Biol. Bull. 1994, 187, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Linacre, A.; Kellett, E.; Saunders, S.; Bright, K.; Benjamin, P.R.; Burke, J.F. Cardioactive neuropeptide Phe-Met-Arg-Phe-NH2 (FMRFamide) and novel related peptides are encoded in multiple copies by a single gene in the snail Lymnaea stagnalis. J. Neurosci. 1990, 10, 412–419. [Google Scholar]
- Eipper, B.A.; Mains, R.E. Peptide α-amidation. Annu. Rev. Physiol. 1988, 50, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Kellett, E.; Saunders, S.E.; Li, K.W.; Staddon, J.W.; Benjamin, P.R.; Burke, J.F. Genomic organization of the FMRFamide gene in Lymnaea: multiple exons encoding novel neuropeptides. J. Neurosci. 1994, 14, 6564–6570. [Google Scholar] [PubMed]
- Lutz, E.M.; Macdonald, M.; Hettle, S.; Price, D.A.; Cottrell, G.A.; Sommerville, J. Structure of cDNA clones and genomic DNA FMRFamide-related peptides (FaRPs) in Helix. Mol. Cell. Neurosci. 1992, 3, 373–382. [Google Scholar] [CrossRef]
- Cristo, C.D.; Lisa, E.D.; Cosmo, A.D. GnRH in the brain and ovary of Sepia officinalis. Peptides 2009, 30, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Onitsuka, C.; Yamaguchi, A.; Kanamaru, H.; Oikawa, S.; Takeda, T.; Matsuyama, M. Molecular Cloning and Expression Analysis of a GnRH-Like Dodecapeptide in the Swordtip Squid, Loligo edulis. Zool. Sci. 2009, 26, 203–208. [Google Scholar] [CrossRef] [PubMed]
- George, D.; Mallery, P. SPSS for Windows Step by Step: A Simple Guide and Reference, 4th ed.; 11.0 Update; Allyn & Bacon: Boston, MA, USA, 2003. [Google Scholar]
- Iwakoshi, U.E.; Ukena, K.K.K.; Kanda, A.; Tsutsui, K.; Minakata, H. Expression and distribution of octopus gonadotropin-releasing hormone in the central nervous system and peripheral organs of the octopus (Octopus vulgaris) by in situ hybridization and immunohistochemistry. J. Comp. Neurol. 2004, 477, 310–323. [Google Scholar] [CrossRef] [PubMed]
- Xin-Xiu, Y.U.; Chang-Wen, W.U.; Chi, C.F. Brain microstructure and optic gland ultrastructure in Sepiella maindroni. Oceanol. Limnol. Sin. 2011, 42, 300–304. [Google Scholar]
- Aroua, S.; Andouche, A.; Martin, M.; Baratte, S.; Bonnaud, L. FaRP cell distribution in the developing CNS suggests the involvement of FaRPs in all parts of the chromatophore control pathway in Sepia officinalis (Cephalopoda). Zoology 2011, 114, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Yamamoto, T.; Nakagawa, M.; Uemura, H. Neuropeptide Y-immunoreactive neuronal system and colocalization with FMRFamide in the optic lobe and peduncle complex of the octopus (Octopus vulgaris). Cell Tissue Res. 2000, 865, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Wollesen, T.; Loesel, R.; Wanninger, A. FMRFamide-like immunoreactivity in the central nervous system of the cephalopod mollusc, Idiosepius notoides. Acta Biol. Hung. 2008, 59 (Suppl. 2), 111–116. [Google Scholar] [CrossRef] [PubMed]
- Dyrlov, B.J.; Nielsen, H.H.G.; Brunak, S. Improved Prediction of Signal Peptides: SignalP 3.0. J. Mol. Biol. 2004, 340, 783–795. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4, a molecular evolutionary genetic analysis MEGA software version 4.0. Mol. Biol. Evol. 2007, 24, 596–1599. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′–3′) | Position |
|---|---|---|
| FMRF-F | CAGCGTGWMCCTTYKTATCTT | 172–192 |
| FMRF-R | CAAAWAAGTCAATCWCGCAGG | 1621–1641 |
| 5′-FMRF-outer | GATGACTTAGGTCCGACTAC | 252–271 |
| 5′-FMRF-inner | GATGGAGCCGCACACACAGAC | 193–213 |
| 3′-FMRF-outer | GCAAAGGGTGTCTGGAAGGT | 1307–1326 |
| 3′-FMRF-inner | GACACACACGCAAACATACAC | 1469–1489 |
| RT-actin-F | TGAGAGGGAGATTGTGCGTG | 815–834 |
| RT-actin-R | GAACATAGATTCTGGAGCACGG | 968–989 |
| RT-FMRF-F | CGTCATCGCCATCTACTGTC | 366–385 |
| RT-FMRF-R | CGCTTGCTTCTCAGTCCATC | 514–533 |
| FMRF-probeF | CCCAAGCGTGATGCGTTGTTGGAGT | 330–348 |
| FMRF-probeR | CCGGAAGCGCTTGCTTCTCAGTCCATC | 514–533 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Cao, Z.; Li, H.; Liu, H.; Lü, Z.; Chi, C. Identification, Characterization, and Expression Analysis of a FMRFamide-Like Peptide Gene in the Common Chinese Cuttlefish (Sepiella japonica). Molecules 2018, 23, 742. https://doi.org/10.3390/molecules23040742
Li Y, Cao Z, Li H, Liu H, Lü Z, Chi C. Identification, Characterization, and Expression Analysis of a FMRFamide-Like Peptide Gene in the Common Chinese Cuttlefish (Sepiella japonica). Molecules. 2018; 23(4):742. https://doi.org/10.3390/molecules23040742
Chicago/Turabian StyleLi, Ying, Zihao Cao, Haifeng Li, Huihui Liu, Zhenming Lü, and Changfeng Chi. 2018. "Identification, Characterization, and Expression Analysis of a FMRFamide-Like Peptide Gene in the Common Chinese Cuttlefish (Sepiella japonica)" Molecules 23, no. 4: 742. https://doi.org/10.3390/molecules23040742
APA StyleLi, Y., Cao, Z., Li, H., Liu, H., Lü, Z., & Chi, C. (2018). Identification, Characterization, and Expression Analysis of a FMRFamide-Like Peptide Gene in the Common Chinese Cuttlefish (Sepiella japonica). Molecules, 23(4), 742. https://doi.org/10.3390/molecules23040742