Mouse β-Defensin 3, A Defensin Inhibitor of Both Its Endogenous and Exogenous Potassium Channels


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Method and Materials
2.1. Construction of rmBD3 Peptide Expression Vector
2.2. Expression and Purification of rmBD3 Fusion Protein
2.3. The Sources of Potassium Channels
2.4. Cell Culture and Transfection
2.5. Electrophysiological Recordings and Data Analysis
3. Results
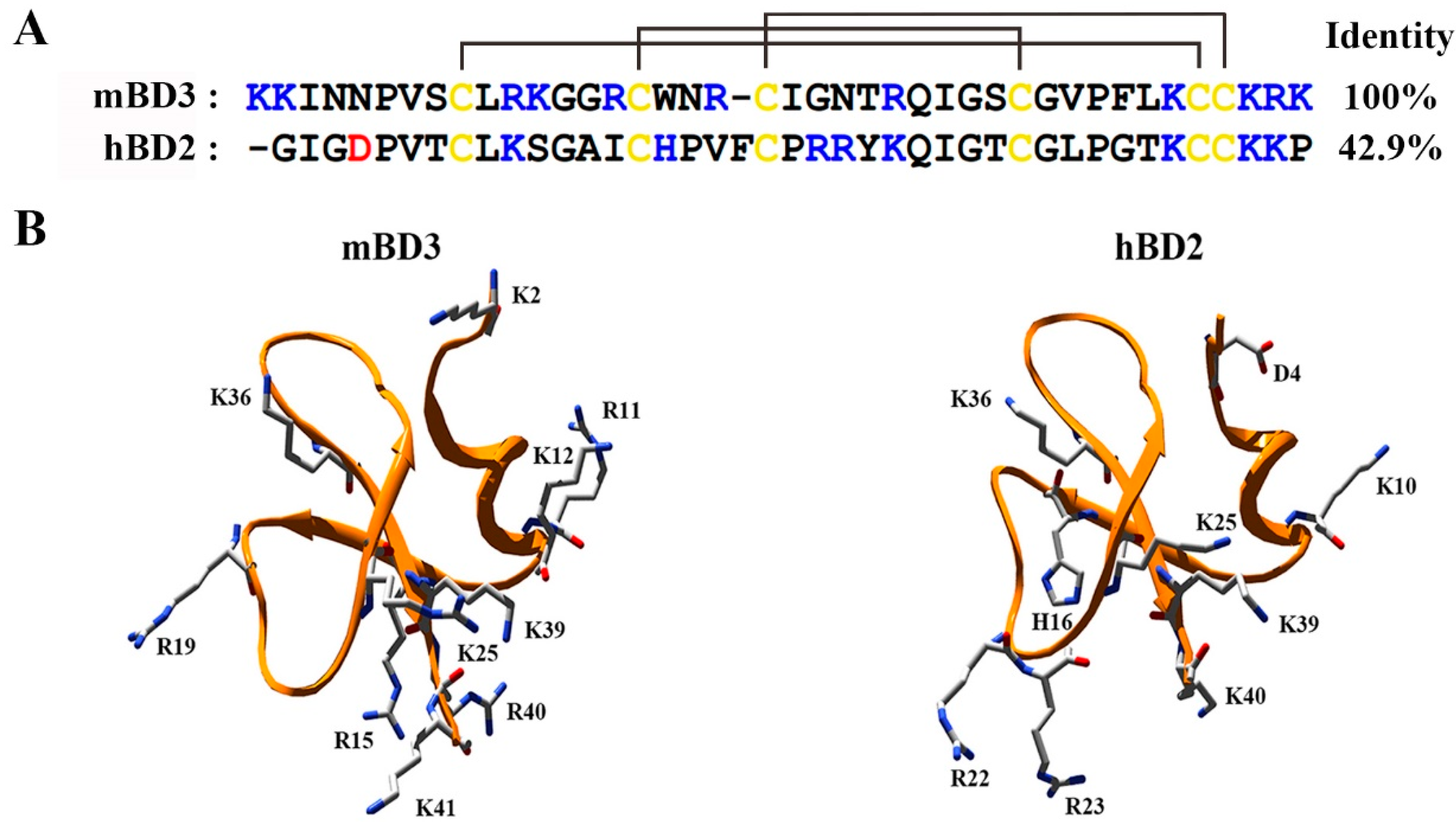
3.1. Structural Analysis of mBD3 as Potential Potassium Channel Inhibitor
3.2. Expression, Purification and Identification of Recombinant mBD3
3.3. Inhibition Effects of Potassium Channels by mBD3
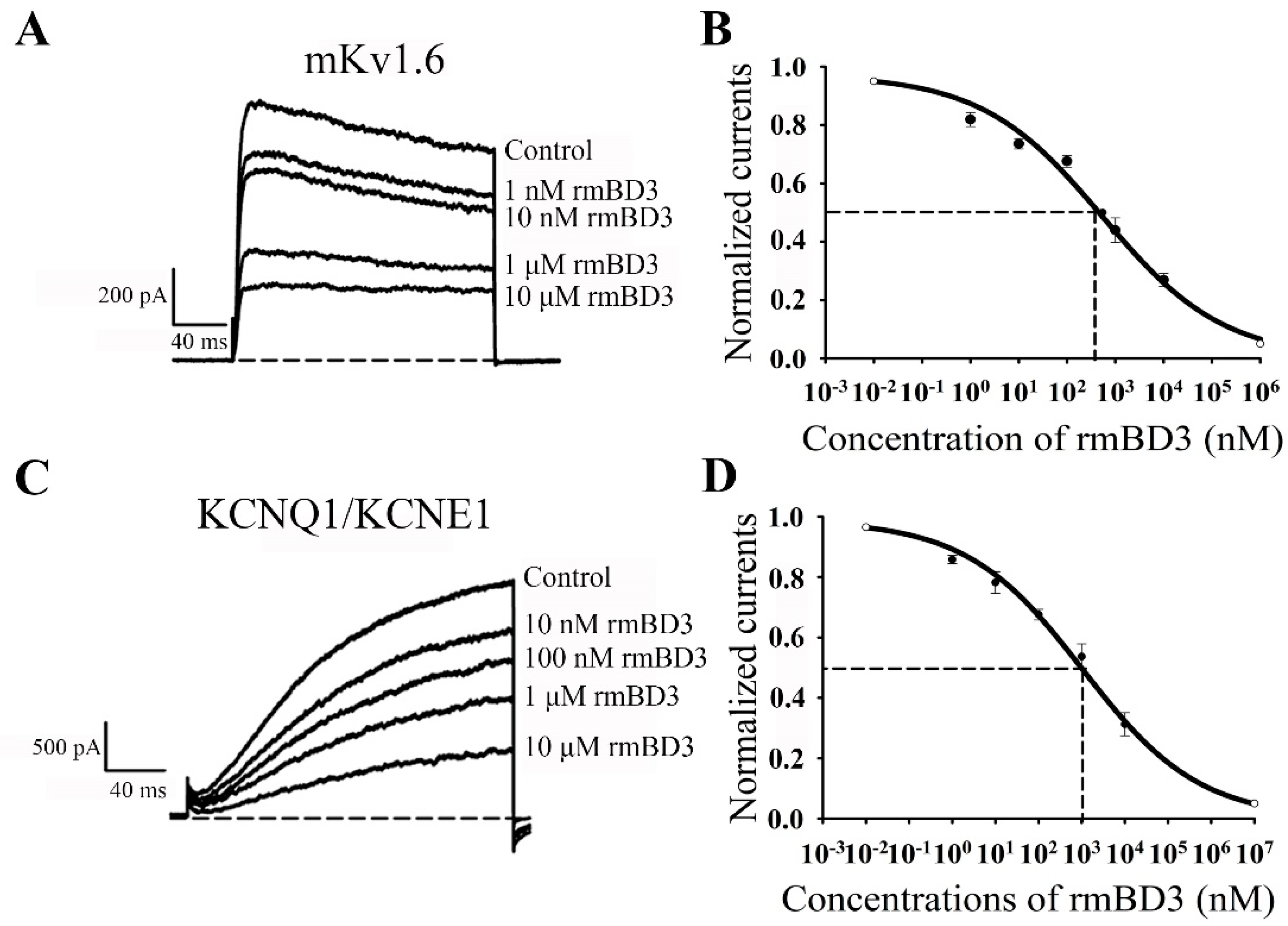
3.4. mBD3 Dose-Dependently Inhibits Mouse Kv1.6 and Human KCNQ1/KCNE1 Channels
3.5. The Pore Region of Mouse Kv1.6 Channel is Responsible for mBD3 Binding
4. Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Ng, T.B.; Cheung, R.C.; Wong, J.H.; Ye, X.J. Antimicrobial activity of defensins and defensin-like peptides with special emphasis on those from fungi and invertebrate animals. Curr. Protein Pept. Sci. 2013, 14, 515–531. [Google Scholar] [CrossRef] [PubMed]
- Yi, H.Y.; Chowdhury, M.; Huang, Y.D.; Yu, X.Q. Insect antimicrobial peptides and their applications. Appl. Microbiol. Biotechnol. 2014, 98, 5807–5822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tassanakajon, A.; Somboonwiwat, K.; Amparyup, P. Sequence diversity and evolution of antimicrobial peptides in invertebrates. Dev. Comp. Immunol. 2015, 48, 324–341. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Nagase, T.; Makita, R.; Fukuhara, S.; Tomita, T.; Tominaga, T.; Kurihara, H.; Ouchi, Y. Identification of multiple novel epididymis-specific beta-defensin isoforms in humans and mice. J. Immunol. 2002, 169, 2516–2523. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Yi, H.; Yin, S.J.; Chen, Z.Y.; Liu, H.; Cao, Z.J.; Wu, Y.; Li, W. Structural basis of a potent peptide inhibitor designed for Kv1.3 channel, a therapeutic target of autoimmune disease. J. Biol. Chem. 2008, 283, 19058–19065. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.J.; Jiang, L.; Yi, H.; Han, S.; Yang, D.W.; Liu, M.L.; Liu, H.; Cao, Z.J.; Wu, Y.L.; Li, W.X. Different residues in channel turret determining the selectivity of ADWX-1 inhibitor peptide between Kv1.1 and Kv1.3 channels. J. Proteome Res. 2008, 7, 4890–4897. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Hu, Y.; Hu, J.; Yang, W.; Sabatier, J.M.; De Waard, M.; Cao, Z.; Li, W.; Han, S.; Wu, Y. Unusual binding mode of scorpion toxin BmKTX onto potassium channels relies on its distribution of acidic residues. Biochem. Biophys. Res. Commun. 2014, 447, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Xiang, F.; Xie, Z.; Feng, J.; Yang, W.; Cao, Z.; Li, W.; Chen, Z.; Wu, Y. Plectasin, first animal toxin-like fungal defensin blocking potassium channels through recognizing channel pore region. Toxins (Basel) 2015, 7, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Vriens, K.; Peigneur, S.; De Coninck, B.; Tytgat, J.; Cammue, B.P.; Thevissen, K. The antifungal plant defensin AtPDF2.3 from Arabidopsis thaliana blocks potassium channels. Sci. Rep. 2016, 30, 32121. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Xie, Z.; Zhang, Q.; Li, Y.; Yang, F.; Chen, Z.; Li, W.; Cao, Z.; Wu, Y. Scorpion Potassium Channel-blocking Defensin Highlights a Functional Link with Neurotoxin. J. Biol. Chem. 2016, 291, 7097–7106. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Feng, J.; Yang, W.; Xiang, F.; Yang, F.; Zhao, Y.; Cao, Z.; Li, W.; Chen, Z.; Wu, Y. Human α-defensins are immune-related Kv1.3 channel inhibitors: New support for their roles in adaptive immunity. FASEB J. 2015, 29, 4324–4333. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Xie, Z.; Yang, W.; Zhao, Y.; Xiang, F.; Cao, Z.; Li, W.; Chen, Z.; Wu, Y. Human beta-defensin 1, a new animal toxin-like blocker of potassium channel. Toxicon 2016, 113, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.X.; Feng, J.; Xiang, F.; Xie, Z.L.; Zhang, G.Y.; Sabatier, J.M.; Cao, Z.J.; Li, W.X.; Chen, Z.Y.; Wu, Y.L. Endogenous animal toxin-like human beta-defensin 2 inhibits own K+ channels through interaction with channel extracellular pore region. Cell. Mol. Life Sci. 2015, 72, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Yang, W.X.; Xie, Z.L.; Xiang, F.; Cao, Z.J.; Li, W.X.; Hu, H.Z.; Chen, Z.Y.; Wu, Y.L. Kv channel S1-S2 linker working as a binding site of human β-defensin 2 for channel activation modulation. J. Biol. Chem. 2015, 290, 15487–15495. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Xu, L.; Liu, H.; He, Y.; Liang, S.; Li, W.; Wu, Y. Characterization of a novel BmαTX47 toxin modulating sodium channels: The crucial role of expression vectors in toxin pharmacological activity. Toxins (Basel) 2014, 6, 816–829. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Liu, S.; Zhang, Y.; Qin, C.; Xu, L.; Li, W.; Cao, Z.; Li, W.; Wu, Y. Expression of recombinant α-toxin BmKM9 from scorpion Buthus martensii Karsch and its functional characterization on sodium channels. Peptides 2017, 99, 30300–30305. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Zeng, D.Y.; Hu, Y.T.; He, Y.W.; Pan, N.; Ding, J.P.; Cao, Z.J.; Liu, M.L.; Li, W.X.; Yi, H.; et al. Structural and functional diversity of acidic scorpion potassium channel toxins. PLoS ONE 2012, 7, e35154. [Google Scholar] [CrossRef] [PubMed]
- Yi, H.; Qiu, S.; Cao, Z.; Wu, Y.; Li, W. Molecular basis of inhibitory peptide maurotoxin recognizing Kv1.2 channel explored by ZDOCK and molecular dynamic simulations. Proteins 2008, 70, 844–854. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Hu, Y.T.; Yang, W.S.; He, Y.W.; Feng, J.; Wang, B.; Zhao, R.M.; Ding, J.P.; Cao, Z.J.; Li, W.X.; et al. Hg1, novel peptide inhibitor specific for Kv1.3 channels from first scorpion Kunitz-type potassium channel toxin family. J. Biol. Chem. 2012, 287, 13813–13821. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Lee, A.; Campbell, E.; Mackinnon, R. Structure of a pore-blocking toxin in complex with a eukaryotic voltage-dependent K+ channel. eLife 2013, 21, e00594. [Google Scholar] [CrossRef] [PubMed]
- Brew, H.M.; Gittelman, J.X.; Silverstein, R.S.; Hanks, T.D.; Demas, V.P.; Robinson, L.C.; Robbins, C.A.; McKee-Johnson, J.; Chiu, S.Y.; Messing, A.; et al. Seizures and reduced life span in mice lacking the potassium channel subunit Kv1.2, but hypoexcitability and enlarged Kv1 currents in auditory neurons. J. Neurophysiol. 2007, 98, 1501–1525. [Google Scholar] [CrossRef] [PubMed]
- Albarwani, S.; Nemetz, L.T.; Madden, J.A.; Tobin, A.A.; England, S.K.; Pratt, P.F.; Rusch, N.J. Voltage-gated K+ channels in rat small cerebral arteries: Molecular identity of the functional channels. J. Physiol. 2003, 15, 751–763. [Google Scholar] [CrossRef] [PubMed]
- Bals, R.; Wang, X.; Meegalla, R.L.; Wattler, S.; Weiner, D.; Nehls, M.C.; Wilson, J.M. Mouse β-defensin 3 is an inducible antimicrobial peptide expressed in the epithelia of multiple organs. Infect. Immun. 1999, 67, 3542–3547. [Google Scholar] [PubMed]
- Harmer, S.C.; Mohal, J.S.; Royal, A.A.; McKenna, W.J.; Lambiase, P.D.; Tinker, A. Cellular mechanisms underlying the increased disease severity seen for patients with long QT syndrome caused by compound mutations in KCNQ1. Biochem. J. 2014, 15, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Chen, J.; Wang, B.; Yang, W.; Zhang, C.; Hu, J.; Xie, Z.; Cao, Z.; Li, W.; Wu, Y.; et al. Engineering a peptide inhibitor towards the KCNQ1/KCNE1 potassium channel (IKs). Peptides 2015, 71, 77–83. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Zhao, Y.; Liu, H.; Yu, W.; Yang, F.; Li, W.; Cao, Z.; Wu, Y. Mouse β-Defensin 3, A Defensin Inhibitor of Both Its Endogenous and Exogenous Potassium Channels. Molecules 2018, 23, 1489. https://doi.org/10.3390/molecules23061489
Zhang Y, Zhao Y, Liu H, Yu W, Yang F, Li W, Cao Z, Wu Y. Mouse β-Defensin 3, A Defensin Inhibitor of Both Its Endogenous and Exogenous Potassium Channels. Molecules. 2018; 23(6):1489. https://doi.org/10.3390/molecules23061489
Chicago/Turabian StyleZhang, Yaoyun, Yonghui Zhao, Hongyue Liu, Weiwei Yu, Fan Yang, Wenhua Li, Zhijian Cao, and Yingliang Wu. 2018. "Mouse β-Defensin 3, A Defensin Inhibitor of Both Its Endogenous and Exogenous Potassium Channels" Molecules 23, no. 6: 1489. https://doi.org/10.3390/molecules23061489
APA StyleZhang, Y., Zhao, Y., Liu, H., Yu, W., Yang, F., Li, W., Cao, Z., & Wu, Y. (2018). Mouse β-Defensin 3, A Defensin Inhibitor of Both Its Endogenous and Exogenous Potassium Channels. Molecules, 23(6), 1489. https://doi.org/10.3390/molecules23061489