
Immunogenicity and Induction of Functional Antibodies in Rabbits Immunized with a Trivalent Typhoid-Invasive Nontyphoidal Salmonella Glycoconjugate Formulation

Abstract
:1. Introduction
2. Results
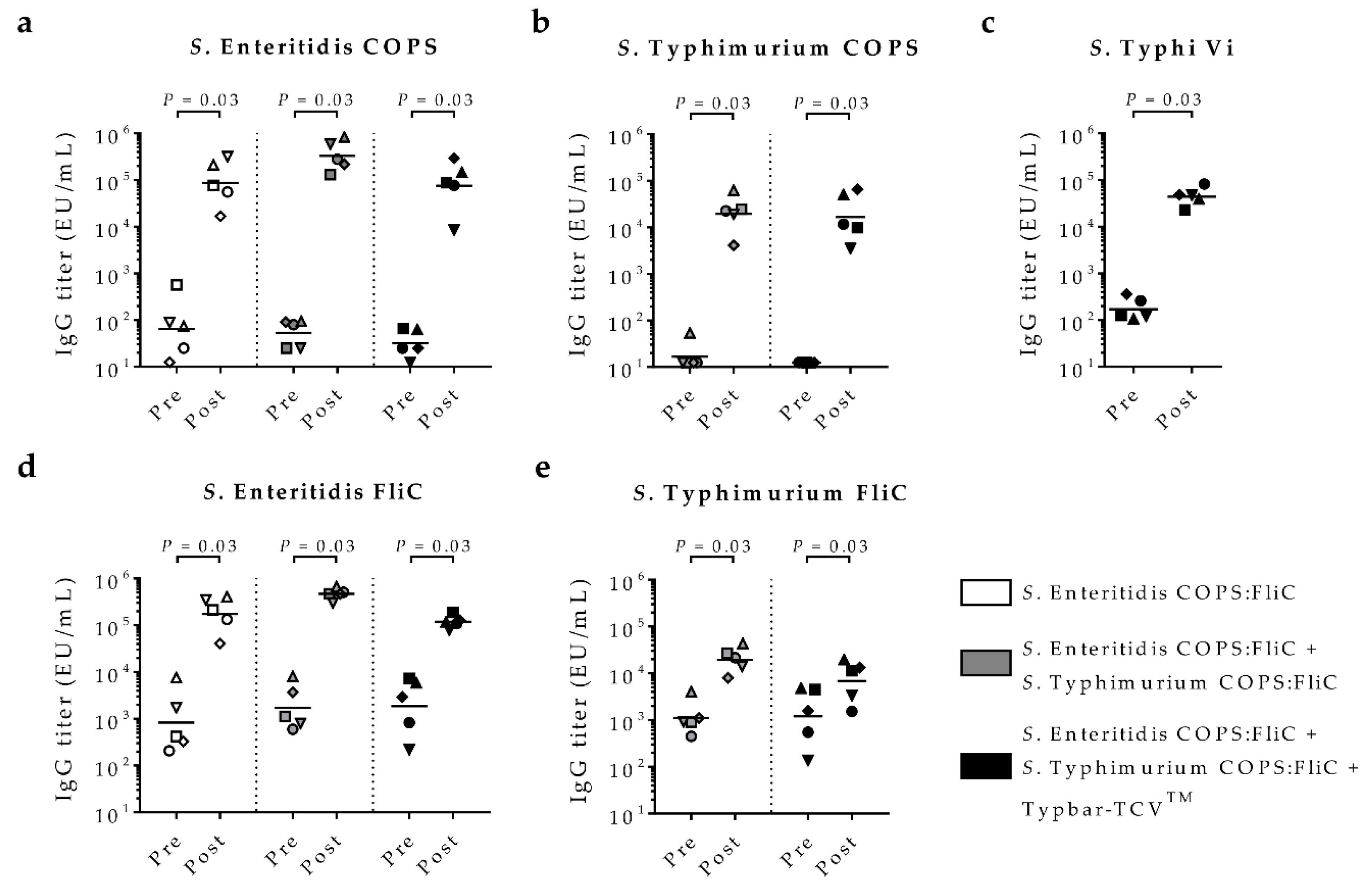
2.1. Immunogenicity of Monovalent and Multivalent Salmonella Glycoconjugate Vaccine Formulations in Rabbits
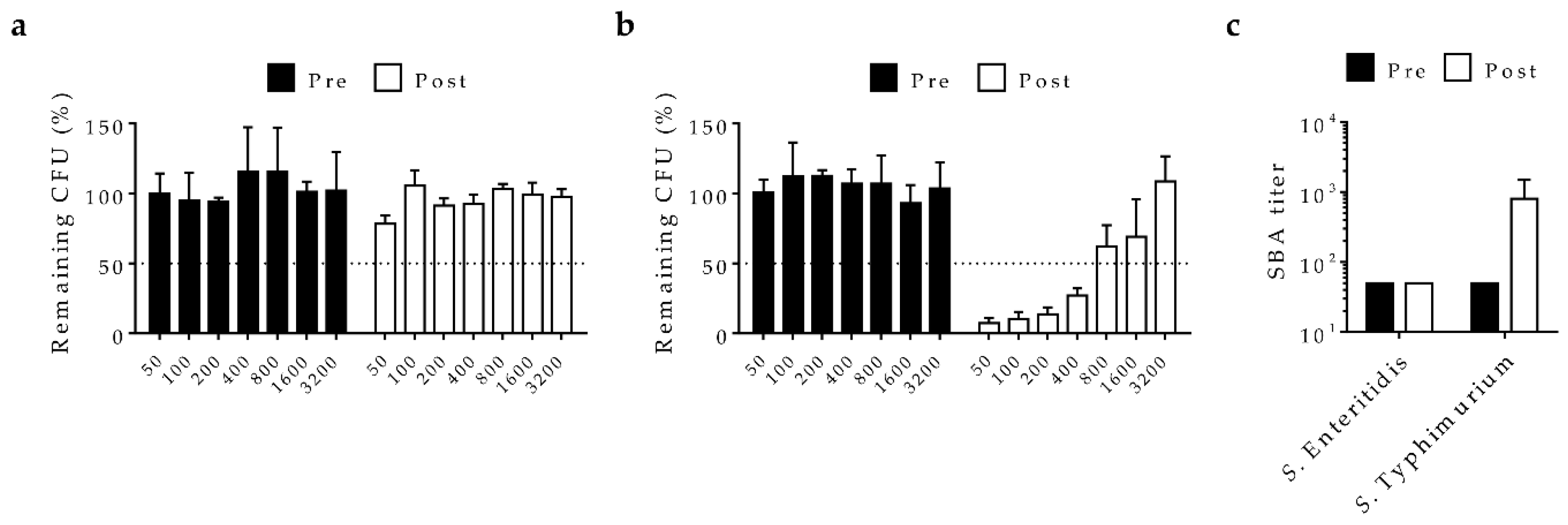
2.2. In-Vitro Functional Bactericidal Activity of Vaccine-Induced Antisera
2.3. Functional Activity of Passively Transferred Antibodies In-Vivo
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
4.2. Polysaccharides
4.3. Flagellin Proteins
4.4. Conjugate Vaccines
4.5. Ethics Statement
4.6. Animal Studies
4.6.1. Rabbit Immunization
4.6.2. Mouse Immunization
4.6.3. Passive Transfer Protection Studies
4.7. Enzyme Linked Immunosorbent Assay (ELISA)
4.8. Antibody Adsorptions
4.9. Western Blot Analysis of Lipopolysaccharide
4.10. Serum Bactericidal Activity (SBA) Assay
4.11. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lindberg, A.A.; Le Minor, L. Serology of Salmonella. In Methods in Microbiology; Bergan, T., Ed.; Academic Press: London, UK, 1984; Volume 15, pp. 1–141. [Google Scholar]
- Marks, F.; von Kalckreuth, V.; Aaby, P.; Adu-Sarkodie, Y.; El Tayeb, M.A.; Ali, M.; Aseffa, A.; Baker, S.; Biggs, H.M.; Bjerregaard-Andersen, M.; et al. Incidence of invasive Salmonella disease in sub-Saharan Africa: a multicentre population-based surveillance study. Lancet Glob. Health 2017, 5, e310–e323. [Google Scholar] [CrossRef]
- Feasey, N.A.; Dougan, G.; Kingsley, R.A.; Heyderman, R.S.; Gordon, M.A. Invasive non-typhoidal Salmonella disease: An emerging and neglected tropical disease in Africa. Lancet 2012, 379, 2489–2499. [Google Scholar] [CrossRef]
- Uche, I.V.; MacLennan, C.A.; Saul, A. A systematic review of the incidence, risk factors and case fatality rates of invasive nontyphoidal Salmonella (iNTS) disease in Africa (1966 to 2014). PLoS Negl. Trop. Dis. 2017, 11, e0005118. [Google Scholar] [CrossRef] [PubMed]
- Reddy, E.A.; Shaw, A.V.; Crump, J.A. Community-acquired bloodstream infections in Africa: A systematic review and meta-analysis. Lancet Infect. Dis. 2010, 10, 417–432. [Google Scholar] [CrossRef]
- Feasey, N.A.; Hadfield, J.; Keddy, K.H.; Dallman, T.J.; Jacobs, J.; Deng, X.; Wigley, P.; Barquist, L.; Langridge, G.C.; Feltwell, T.; et al. Distinct Salmonella Enteritidis lineages associated with enterocolitis in high-income settings and invasive disease in low-income settings. Nat. Genet. 2016, 48, 1211–1217. [Google Scholar] [CrossRef] [PubMed]
- Kingsley, R.A.; Msefula, C.L.; Thomson, N.R.; Kariuki, S.; Holt, K.E.; Gordon, M.A.; Harris, D.; Clarke, L.; Whitehead, S.; Sangal, V.; et al. Epidemic multiple drug resistant Salmonella Typhimurium causing invasive disease in sub-Saharan Africa have a distinct genotype. Genome Res. 2009, 19, 2279–2287. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, G.; Panda, A.; Higginson, E.E.; Ateh, E.; Lipsky, M.M.; Sen, S.; Matson, C.A.; Permala-Booth, J.; DeTolla, L.J.; Tennant, S.M. Virulence of invasive Salmonella Typhimurium ST313 in animal models of infection. PLoS Negl. Trop. Dis. 2017, 11, e0005697. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, G.; Perkins, D.J.; Schmidlein, P.J.; Tulapurkar, M.E.; Tennant, S.M. Invasive Salmonella Typhimurium ST313 with naturally attenuated flagellin elicits reduced inflammation and replicates within macrophages. PLoS Negl. Trop. Dis. 2015, 9, e3394. [Google Scholar] [CrossRef] [PubMed]
- Crump, J.A.; Heyderman, R.S. A Perspective on Invasive Salmonella Disease in Africa. Clin. Infect. Dis. 2015, 6 (Suppl. 4), S235–S240. [Google Scholar] [CrossRef]
- Klugman, K.P.; Koornhof, H.J.; Robbins, J.B.; Le Cam, N.N. Immunogenicity, efficacy and serological correlate of protection of Salmonella Typhi Vi capsular polysaccharide vaccine three years after immunization. Vaccine 1996, 14, 435–438. [Google Scholar] [CrossRef]
- Carlin, N.I.; Svenson, S.B.; Lindberg, A.A. Role of monoclonal O-antigen antibody epitope specificity and isotype in protection against experimental mouse typhoid. Microb. Pathogenes. 1987, 2, 171–183. [Google Scholar] [CrossRef]
- Simon, R.; Wang, J.Y.; Boyd, M.A.; Tulapurkar, M.E.; Ramachandran, G.; Tennant, S.M.; Pasetti, M.; Galen, J.E.; Levine, M.M. Sustained protection in mice immunized with fractional doses of Salmonella Enteritidis core and O polysaccharide-flagellin glycoconjugates. PLoS ONE 2013, 8, e64680. [Google Scholar] [CrossRef] [PubMed]
- Micoli, F.; Ravenscroft, N.; Cescutti, P.; Stefanetti, G.; Londero, S.; Rondini, S.; Maclennan, C.A. Structural analysis of O-polysaccharide chains extracted from different Salmonella Typhimurium strains. Carbohydr. Res. 2014, 385, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ada, G.; Isaacs, D. Carbohydrate-protein conjugate vaccines. Clin. Microbiol. Infect. 2003, 9, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Astronomo, R.D.; Burton, D.R. Carbohydrate vaccines: Developing sweet solutions to sticky situations? Nat. Rev. Drug Discov. 2010, 9, 308–324. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Typbar TCV® from Bharat Biotech, World’s First Typhoid Conjugate Vaccine Prequalified by WHO Geneva. Available online: http://www.who.int/medicines/news/2017/WHOprequalifies-breakthrough-typhoid-vaccine/en/ (accessed on 11 July 2018).
- Baliban, S.M.; Yang, M.; Ramachandran, G.; Curtis, B.; Shridhar, S.; Laufer, R.S.; Wang, J.Y.; Van Druff, J.; Higginson, E.E.; Hegerle, N.; et al. Development of a glycoconjugate vaccine to prevent invasive Salmonella Typhimurium infections in sub-Saharan Africa. PLoS Negl. Trop. Dis. 2017, 11, e0005493. [Google Scholar] [CrossRef] [PubMed]
- Simon, R.; Tennant, S.M.; Wang, J.Y.; Schmidlein, P.J.; Lees, A.; Ernst, R.K.; Pasetti, M.F.; Galen, J.E.; Levine, M.M. Salmonella enterica serovar Enteritidis core O polysaccharide conjugated to H:g,m flagellin as a candidate vaccine for protection against invasive infection with S. Enteritidis. Infect. Immun. 2011, 79, 4240–4249. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, G.; Tennant, S.M.; Boyd, M.A.; Wang, J.Y.; Tulapurkar, M.E.; Pasetti, M.F.; Levine, M.M.; Simon, R. Functional activity of antibodies directed towards flagellin proteins of non-typhoidal Salmonella. PLoS ONE 2016, 11, e0151875. [Google Scholar] [CrossRef] [PubMed]
- Baliban, S.M.; Curtis, B.; Toema, D.; Tennant, S.M.; Levine, M.M.; Pasetti, M.F.; Simon, R. Immunogenicity and efficacy following sequential parenterally-administered doses of Salmonella Enteritidis COPS:FliC glycoconjugates in infant and adult mice. PLoS Negl. Trop. Dis. 2018, 12, e0006522. [Google Scholar] [CrossRef] [PubMed]
- Jorbeck, H.J.; Svenson, S.B.; Lindberg, A.A. Artificial Salmonella vaccines: Salmonella Typhimurium O-antigen-specific oligosaccharide-protein conjugates elicit opsonizing antibodies that enhance phagocytosis. Infect. Immun. 1981, 32, 497–502. [Google Scholar] [PubMed]
- Svenson, S.B.; Lindberg, A.A. Artificial Salmonella vaccines: Salmonella Typhimurium O-antigen-specific oligosaccharide-protein conjugates elicit protective antibodies in rabbits and mice. Infect. Immun. 1981, 32, 490–496. [Google Scholar] [PubMed]
- Tennant, S.M.; Wang, J.Y.; Galen, J.E.; Simon, R.; Pasetti, M.F.; Gat, O.; Levine, M.M. Engineering and preclinical evaluation of attenuated nontyphoidal Salmonella strains serving as live oral vaccines and as reagent strains. Infect. Immun. 2011, 79, 4175–4185. [Google Scholar] [CrossRef] [PubMed]
- Boyd, M.A.; Tennant, S.M.; Saague, V.A.; Simon, R.; Muhsen, K.; Ramachandran, G.; Cross, A.S.; Galen, J.E.; Pasetti, M.F.; Levine, M.M. Serum bactericidal assays to evaluate typhoidal and nontyphoidal Salmonella vaccines. Clin. Vaccine Immunol. 2014, 21, 712–721. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Noriega, F.R.; Galen, J.E.; Barry, E.; Levine, M.M. Constitutive expression of the Vi polysaccharide capsular antigen in attenuated Salmonella enterica serovar Typhi oral vaccine strain CVD 909. Infect. Immun. 2000, 68, 4647–4652. [Google Scholar] [CrossRef] [PubMed]
- Fuche, F.J.; Jones, J.A.; Ramachandran, G.; Higginson, E.E.; Simon, R.; Tennant, S.M. Deletions in guaBA and htrA but not clpX or rfaL constitute a live-attenuated vaccine strain of Salmonella Newport to protect against serogroup C2-C3 Salmonella in mice. Hum. Vaccin. Immunother. 2018, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Fiorino, F.; Rondini, S.; Micoli, F.; Lanzilao, L.; Alfini, R.; Mancini, F.; MacLennan, C.A.; Medaglini, D. Immunogenicity of a bivalent adjuvanted glycoconjugate vaccine against Salmonella Typhimurium and Salmonella Enteritidis. Front. Immunol. 2017, 8, 168. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.; Peng, H.; Rader, C. From rabbit antibody repertoires to rabbit monoclonal antibodies. Exp. Mol. Med. 2017, 49, e305. [Google Scholar] [CrossRef] [PubMed]
- Prasanphanich, N.S.; Song, X.; Heimburg-Molinaro, J.; Luyai, A.E.; Lasanajak, Y.; Cutler, C.E.; Smith, D.F.; Cummings, R.D. Intact reducing glycan promotes the specific immune response to lacto-N-neotetraose-BSA neoglycoconjugates. Bioconj. Chem. 2015, 26, 559–571. [Google Scholar] [CrossRef] [PubMed]
- De Velasco, E.A.; Verheul, A.F.; van Steijn, A.M.; Dekker, H.A.; Feldman, R.G.; Fernandez, I.M.; Kamerling, J.P.; Vliegenthart, J.F.; Verhoef, J.; Snippe, H. Epitope specificity of rabbit immunoglobulin G (IgG) elicited by pneumococcal type 23F synthetic oligosaccharide- and native polysaccharide-protein conjugate vaccines: Comparison with human anti-polysaccharide 23F IgG. Infect. Immun. 1994, 62, 799–808. [Google Scholar]
- Higginson, E.E.; Simon, R.; Tennant, S.M. Animal models for salmonellosis: Applications in vaccine research. Clin. Vaccine Immunol. 2016, 23, 746–756. [Google Scholar] [CrossRef] [PubMed]
- Simon, R.; Tennant, S.M.; Galen, J.E.; Levine, M.M. Mouse models to assess the efficacy of non-typhoidal Salmonella vaccines: Revisiting the role of host innate susceptibility and routes of challenge. Vaccine 2011, 29, 5094–5106. [Google Scholar] [CrossRef] [PubMed]
- Goh, Y.S.; Clare, S.; Micoli, F.; Saul, A.; Mastroeni, P.; MacLennan, C.A. Monoclonal antibodies of a diverse isotype induced by an O-antigen glycoconjugate vaccine mediate in vitro and in vivo killing of african invasive nontyphoidal Salmonella. Infect. Immun. 2015, 83, 3722–3731. [Google Scholar] [CrossRef] [PubMed]
- Lanzilao, L.; Stefanetti, G.; Saul, A.; MacLennan, C.A.; Micoli, F.; Rondini, S. Strain selection for generation of O-antigen-based glycoconjugate vaccines against invasive nontyphoidal Salmonella disease. PLoS ONE 2015, 10, e0139847. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.Y.; Ho, V.A.; Khiem, H.B.; Trach, D.D.; Bay, P.V.; Thanh, T.C.; Kossaczka, Z.; Bryla, D.A.; Shiloach, J.; Robbins, J.B.; et al. The efficacy of a Salmonella Typhi Vi conjugate vaccine in two-to-five-year-old children. New Engl. J. Med. 2001, 344, 1263–1269. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Gibani, M.M.; Moore, M.; Juel, H.B.; Jones, E.; Meiring, J.; Harris, V.; Gardner, J.; Nebykova, A.; Kerridge, S.A.; et al. Efficacy and immunogenicity of a Vi-tetanus toxoid conjugate vaccine in the prevention of typhoid fever using a controlled human infection model of Salmonella Typhi: A randomised controlled, phase 2b trial. Lancet 2017, 390, 2472–2480. [Google Scholar] [CrossRef]
- Mohan, V.K.; Varanasi, V.; Singh, A.; Pasetti, M.F.; Levine, M.M.; Venkatesan, R.; Ella, K.M. Safety and immunogenicity of a Vi polysaccharide-tetanus toxoid conjugate vaccine (Typbar-TCV) in healthy infants, children, and adults in typhoid endemic areas: A multicenter, 2-cohort, open-label, double-blind, randomized controlled phase 3 study. Clin. Infect. Dis. 2015, 61, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Nyirenda, T.S.; Gilchrist, J.J.; Feasey, N.A.; Glennie, S.J.; Bar-Zeev, N.; Gordon, M.A.; MacLennan, C.A.; Mandala, W.L.; Heyderman, R.S. Sequential acquisition of T cells and antibodies to nontyphoidal Salmonella in Malawian children. J. Infect. Dis. 2014, 210, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Sow, S.O.; Tapia, M.D.; Diallo, S.; Keita, M.M.; Sylla, M.; Onwuchekwa, U.; Pasetti, M.F.; Kotloff, K.L.; Levine, M.M. Haemophilus influenzae Type B conjugate vaccine introduction in Mali: Impact on disease burden and serologic correlate of protection. Am. J. Trop. Med. Hyg. 2009, 80, 1033–1038. [Google Scholar] [PubMed]
- Bornstein, K.; Hungerford, L.; Hartley, D.; Sorkin, J.D.; Tapia, M.D.; Sow, S.O.; Onwuchekwa, U.; Simon, R.; Tennant, S.M.; Levine, M.M. Modeling the potential for vaccination to diminish the burden of invasive non-typhoidal Salmonella disease in young children in Mali, West Africa. PLoS Negl. Trop. Dis. 2017, 11, e0005283. [Google Scholar] [CrossRef] [PubMed]
- Hestrin, S. The reaction of acetylcholine and other carboxylic acid derivatives with hydroxylamine, and its analytical application. J. Biol. Chem. 1949, 180, 249–261. [Google Scholar] [PubMed]
- Simon, R.; Curtis, B.; Deumic, V.; Nicki, J.; Tennant, S.M.; Pasetti, M.F.; Lees, A.; Wills, P.W.; Chacon, M.; Levine, M.M. A scalable method for biochemical purification of Salmonella flagellin. Protein Expr. Purif. 2014, 102, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Watson, D.C.; Robbins, J.B.; Szu, S.C. Protection of mice against Salmonella Typhimurium with an O-specific polysaccharide-protein conjugate vaccine. Infect. Immun. 1992, 60, 4679–4686. [Google Scholar] [PubMed]
Sample Availability: Samples of the NTS conjugate vaccine compounds are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Characteristics (Use) | Reference |
|---|---|---|
| S. Enteritidis R11 | Wild-type Malian blood isolate (challenge strain, epitope specificity analyses) | [24] |
| S. Enteritidis S15 | Wild-type Malian blood isolate (functional antibody analyses) | [25] |
| S. Typhimurium D65 | Wild-type Malian blood isolate (challenge strain, functional antibody analyses) | [24] |
| S. Typhimurium I77 | Wild-type Malian blood isolate (epitope specificity analyses) | [24] |
| S. Enteritidis CVD 1943 | R11 ΔguaBA ΔclpP ΔfliD (reagent strain for S. Enteritidis COPS and FliC production) | [24] |
| S. Typhimurium CVD 1925 | S. Typhimurium I77 ΔguaBA ΔclpP ΔfliD ΔfljB (reagent strain for S. Typhimurium FliC production) | [24] |
| S. Typhimurium CVD 1925wzzB | S. Typhimurium I77 ΔguaBA ΔclpP ΔfliD ΔfljB pSEC10-wzzB (reagent strain for S. Typhimurium COPS production) | [18] |
| S. Typhi CVD 909 | S. Typhi Ty2 ΔaroCD ΔhtrA PtacviaB (reagent strain for Vi production) | [26] |
| S. Newport Chile 361 ΔrfaL | Genetically mutated S. Newport isolate lacking OPS (epitope specificity analyses) | [27] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baliban, S.M.; Allen, J.C.; Curtis, B.; Amin, M.N.; Lees, A.; Rao, R.N.; Naidu, G.; Venkatesan, R.; Rao, D.Y.; Mohan, V.K.; et al. Immunogenicity and Induction of Functional Antibodies in Rabbits Immunized with a Trivalent Typhoid-Invasive Nontyphoidal Salmonella Glycoconjugate Formulation. Molecules 2018, 23, 1749. https://doi.org/10.3390/molecules23071749
Baliban SM, Allen JC, Curtis B, Amin MN, Lees A, Rao RN, Naidu G, Venkatesan R, Rao DY, Mohan VK, et al. Immunogenicity and Induction of Functional Antibodies in Rabbits Immunized with a Trivalent Typhoid-Invasive Nontyphoidal Salmonella Glycoconjugate Formulation. Molecules. 2018; 23(7):1749. https://doi.org/10.3390/molecules23071749
Chicago/Turabian StyleBaliban, Scott M., Jessica C. Allen, Brittany Curtis, Mohammed N. Amin, Andrew Lees, R. Nageswara Rao, Gangadhara Naidu, Ramasamy Venkatesan, D. Yogeswara Rao, Vadrevu Krishna Mohan, and et al. 2018. "Immunogenicity and Induction of Functional Antibodies in Rabbits Immunized with a Trivalent Typhoid-Invasive Nontyphoidal Salmonella Glycoconjugate Formulation" Molecules 23, no. 7: 1749. https://doi.org/10.3390/molecules23071749
APA StyleBaliban, S. M., Allen, J. C., Curtis, B., Amin, M. N., Lees, A., Rao, R. N., Naidu, G., Venkatesan, R., Rao, D. Y., Mohan, V. K., Ella, K. M., Levine, M. M., & Simon, R. (2018). Immunogenicity and Induction of Functional Antibodies in Rabbits Immunized with a Trivalent Typhoid-Invasive Nontyphoidal Salmonella Glycoconjugate Formulation. Molecules, 23(7), 1749. https://doi.org/10.3390/molecules23071749