Effects of Rhizome Extract of Dioscorea batatas and Its Active Compound, Allantoin, on the Regulation of Myoblast Differentiation and Mitochondrial Biogenesis in C2C12 Myotubes

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Effects of Yam Extract and Allantoin on Myoblast Differentiation into Myotubes
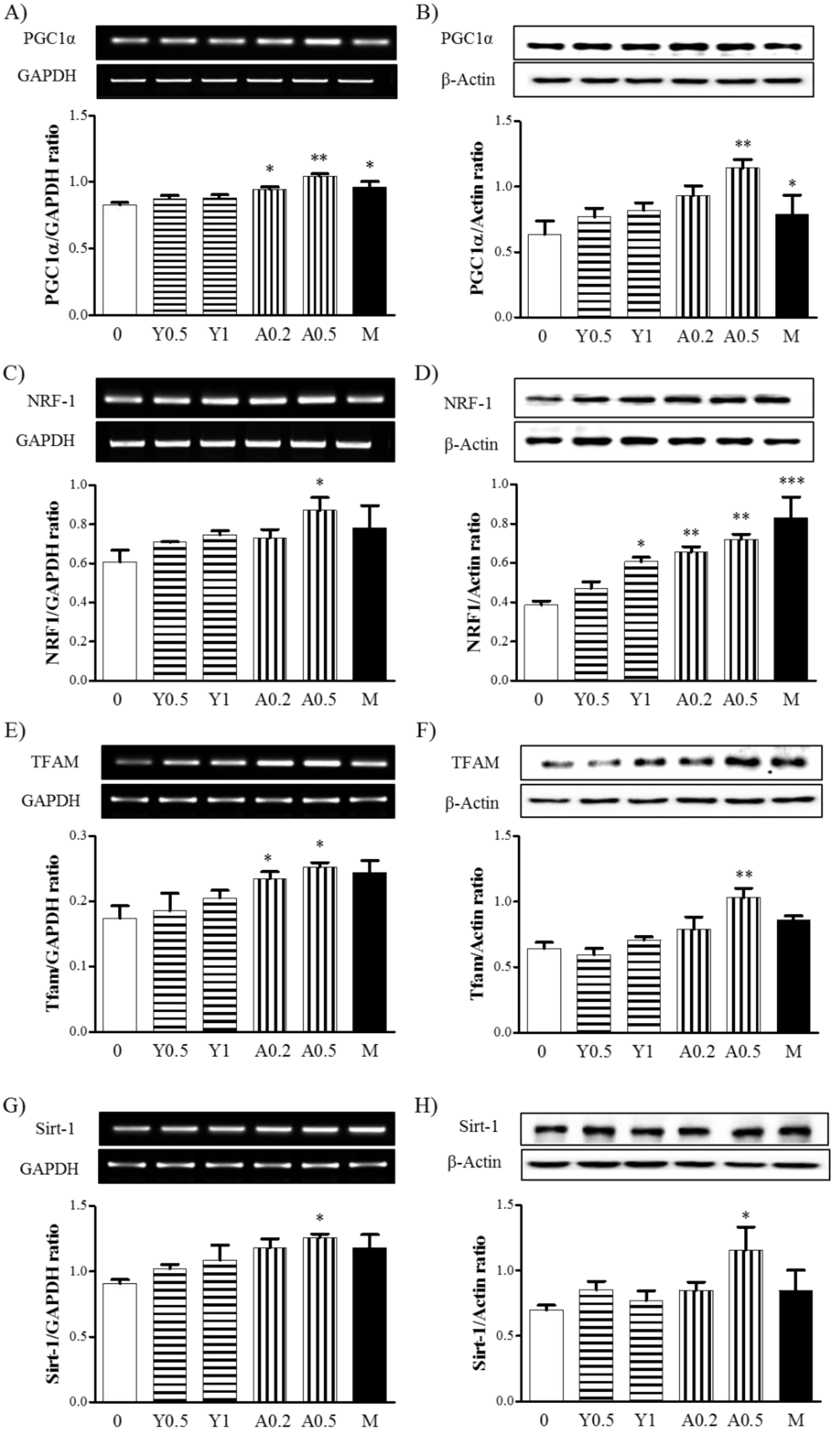
2.2. Effects of Yam Extract and Allantoin on the Expression of Mitochondria Biogenesis-Regulating Factors in Myotubes
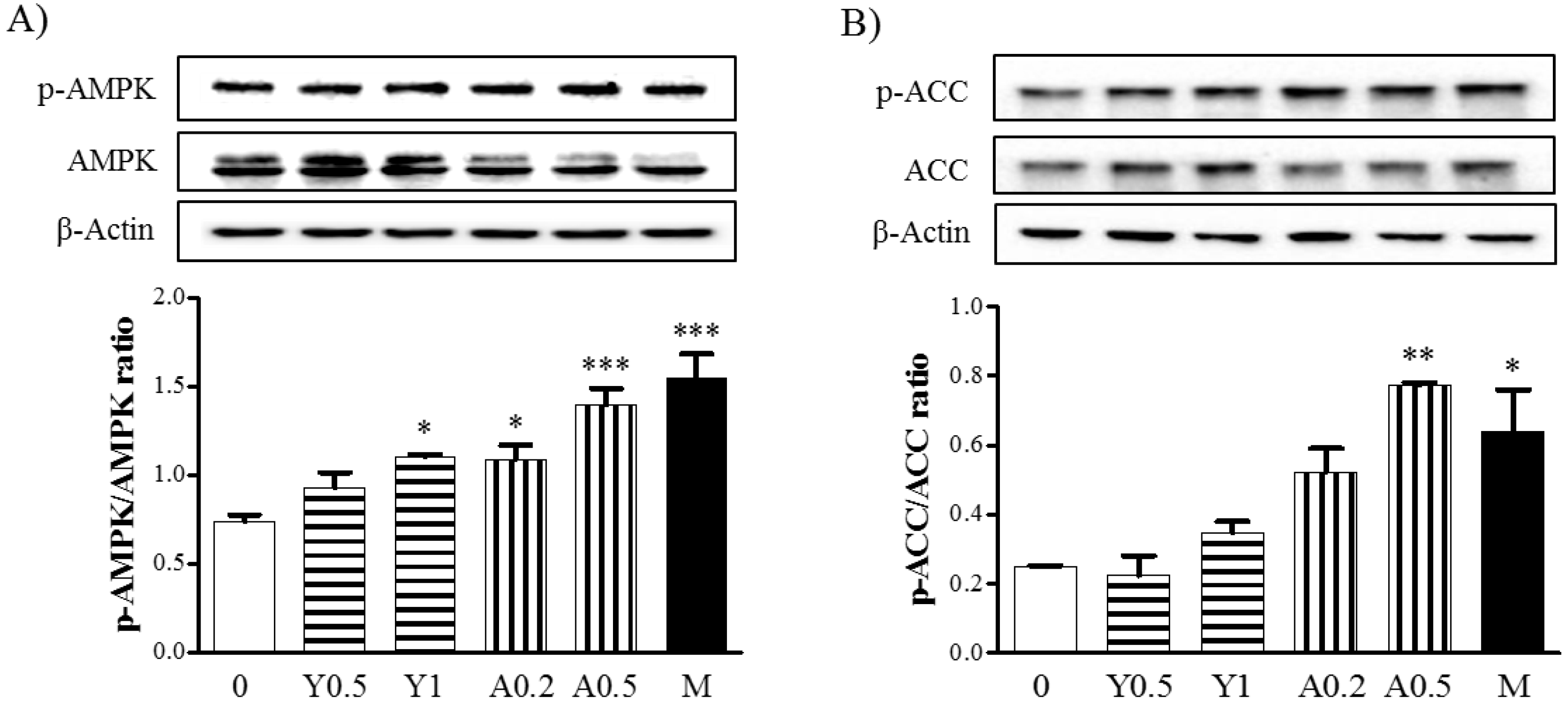
2.3. Effects of Yam Extract and Allantoin on the AMPK and ACC Pathways in Myotubes
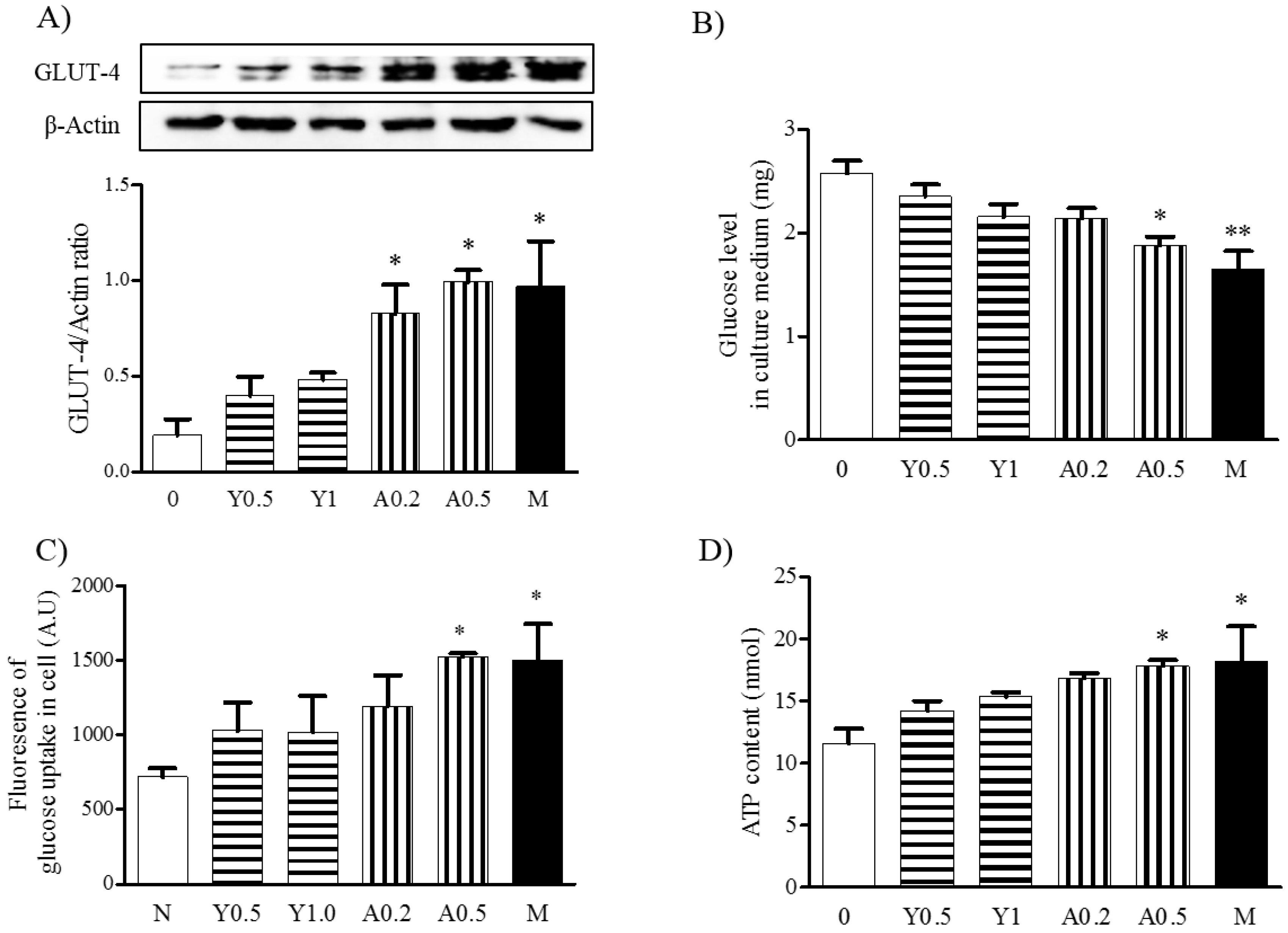
2.4. Effects of Yam Extract and Allantoin on Glucose Uptake in Myotubes
2.5. Effects of Yam Extract and Allantoin on ATP Production in Myotubes
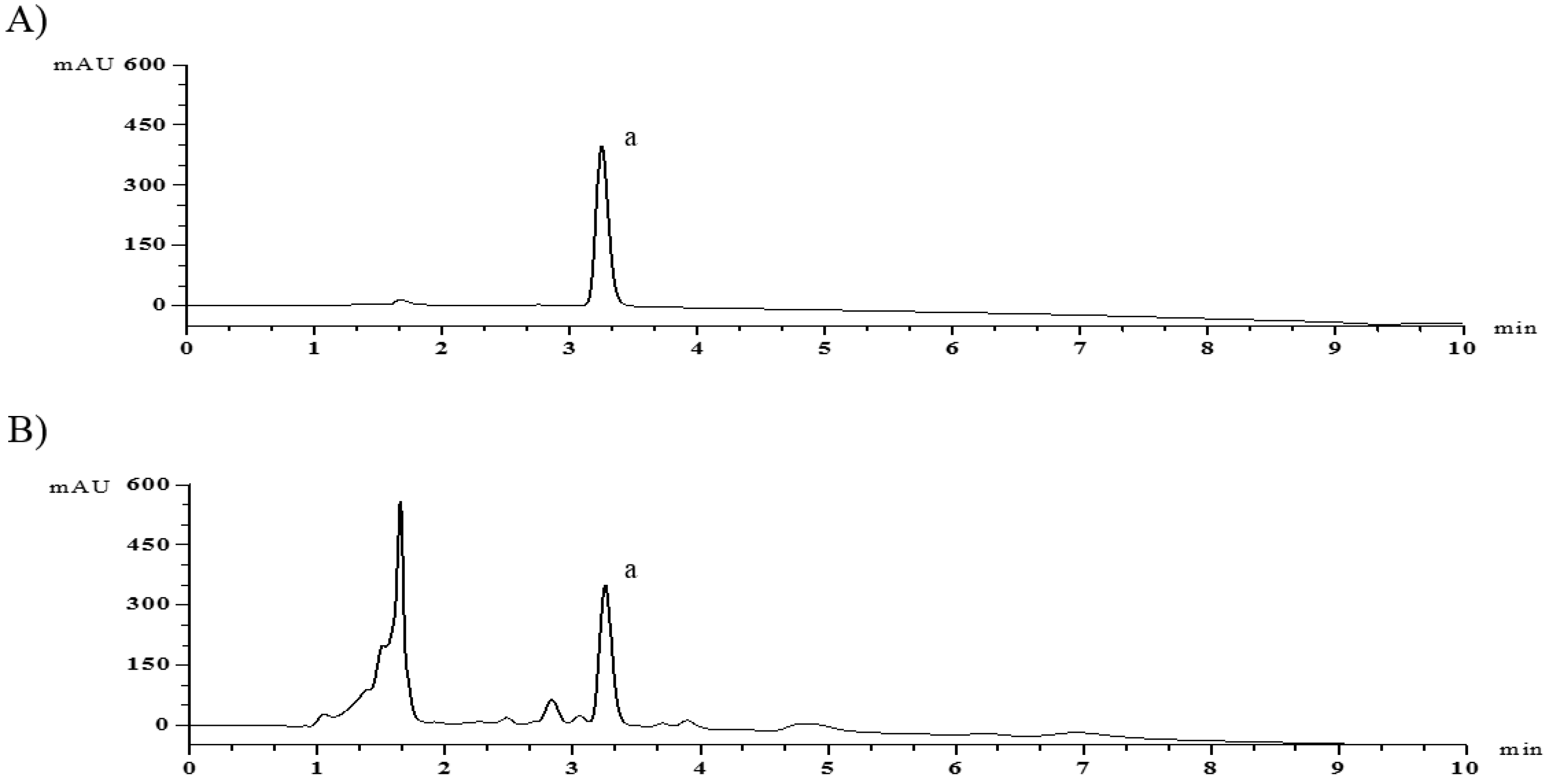
2.6. HPLC Analysis
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation of Yam Extract
4.3. Cell Culture and Drug Treatments
4.4. Western Blot
4.5. Reverse Transcriptase (RT)-Polymerase Chain Reaction (RT-PCR)
4.6. Immunocytochemistry
4.7. Glucose Assay
4.8. ATP Assay
4.9. HPLC Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ACC | Acetyl-CoA carboxylase |
| AMPK | AMP-activated protein kinase |
| BSA | Bovine serum albumin |
| DMEM | Dulbecco’s modified Eagle’s medium |
| FBS | Fetal bovine serum |
| HRP | Horseradish peroxidase |
| HS | Horse serum |
| MRFs | Myogenic regulatory factors |
| Myf5 | Myogenic factor 5 |
| MyoD | Myoblast determination protein |
| MyHC | Myosin heavy chain |
| NRF-1 | Nuclear respiratory factor-1 |
| RT-PCR | Reverse transcriptase-polymerase chain reaction |
| PAGE | Polyacrylamide gel electrophoresis |
| PBS | Phosphate buffered saline |
| PGC1α | Peroxisome proliferator-activated receptor gamma coactivator |
| RIPA | Radioimmunoprecipitation assay |
| Sirt-1 | Sirtuin 1 |
| TBST | Tris-buffered saline containing 0.1% tween-20 |
| TFAM | Transcription factor A, mitochondrial |
| HPLC | High Performance Liquid Chromatography |
References
- Pyla, R.; Pichavaram, P.; Fairaq, A.; Park, M.A.; Kozak, M.; Kamath, V.; Patel, V.S.; Segar, L. Altered energy state reversibly controls smooth muscle contractile function in human saphenous vein during acute hypoxia-reoxygenation: Role of glycogen, AMP-activated protein kinase, and insulin-independent glucose uptake. Biochem. Pharmacol. 2015, 97, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Papa, E.V.; Dong, X.; Hassan, M. Skeletal Muscle Function Deficits in the Elderly: Current Perspectives on Resistance Training. J. Nat. Sci. 2017, 1, e272. [Google Scholar]
- Kalyani, P.R.; Corriere, M.; Ferrucci, L. Age-related and disease-related muscle loss: The effect of diabetes, obesity, and other diseases. Lancet Diabetes Endocrinol. 2014, 10, 819–829. [Google Scholar] [CrossRef]
- Jung, H.W.; Kang, A.N.; Kang, S.Y.; Park, Y.K.; Song, M.Y. The Root Extract of Puerarialobata and Its Main Compound, Puerarin, Prevent Obesity by Increasing the Energy Metabolism in Skeletal Muscle. Nutrients 2017, 9, 33. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.R.; Kim, H.C.; Kook, Y.B.; Park, S.J.; Park, Y.K.; Park, J.H.; Seo, B.I.; Seo, Y.B.; Song, H.J.; Shin, M.K.; et al. Herbology; Yonglims: Seoul, Korea, 2007; pp. 580–581. ISBN 978-89-85897-44-0. [Google Scholar]
- Ling, Y.Q. Chinese Herbal Medicine; Shanghai Scientific and Technical Publishers: Shanghai, China, 2000; p. 214. [Google Scholar]
- Park, H.S.; Kim, M.J.; Moon, H.B. Occupational asthma caused by two herb materials, Dioscorea batatas and Pinellia ternata. Clin. Exp. Allergy 1994, 24, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, N.; Nagasawa, T.; Mabuchi, R.; Yasui, Y.; Wakabayashi, K.; Tanaka, T.; Ohshima, H. Chemoprevention of azoxymethane/dextran sodium sulfate-induced mouse colon carcinogenesis by freeze-dried yam sanyaku and its constituent diosgenin. Cancer Prev. Res. 2011, 4, 924–934. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, H.; Fan, Y.; Man, S.; Liu, Z.; Gao, W.; Wang, T. Antioxidant and Antitumor Activities of the Extracts from Chinese Yam (Dioscorea opposite Thunb) Flesh and Peel and the Effective Compounds. J. Food Sci. 2016, 81, 1553–1564. [Google Scholar] [CrossRef] [PubMed]
- Tsukayama, I.; Toda, K.; Takeda, Y.; Mega, T.; Tanaka, M.; Kawakami, Y.; Takahashi, Y.; Kimoto, M.; Yamamoto, K.; Miki, Y.; et al. Preventive effect of Dioscorea japonica on squamous cell carcinoma of mouse skin involving down-regulation of prostaglandin E2 synthetic pathway. J. Clin. Biochem. Nutr. 2018, 62, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Jwa, H.; Yanagawa, Y.; Park, T. Extract from Dioscorea batatas ameliorates insulin resistance in mice fed a high-fat diet. J. Med. Food 2012, 15, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Go, H.K.; Rahman, M.M.; Kim, G.B.; Na, C.S.; Song, C.H.; Kim, J.S.; Kim, S.J.; Kang, H.S. Antidiabetic Effects of Yam (Dioscorea batatas) and Its Active Constituent, Allantoin, in a Rat Model of Streptozotocin-Induced Diabetes. Nutrients 2015, 7, 8532–8544. [Google Scholar] [CrossRef] [PubMed]
- Yeh, Y.H.; Hsieh, Y.L.; Lee, Y.T. Effects of yam peel extract against carbon tetrachloride-induced hepatotoxicity in rats. J. Agric. Food Chem. 2013, 61, 7387–7396. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.S.; Deng, J.S.; Chang, H.Y.; Chen, Y.C.; Lee, M.M.; Hou, W.C.; Lee, C.Y.; Huang, S.S.; Huang, G.J. Antioxidant and anti-inflammatory properties of Taiwanese yam (Dioscorea japonica Thunb. var. pseudojaponica (Hayata) Yamam.) and its reference compounds. Food Chem. 2013, 141, 1087–1096. [Google Scholar] [PubMed]
- Cronin, H.; Draelos, Z.D. Top 10 botanical ingredients in 2010 anti-aging creams. J. Cosmet. Dermatol. 2010, 9, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.R.; Lee, J.S.; Lee, C.H.; Kim, J.Y.; Kim, S.D.; Nam, D.H. Effect of ethanol extract of dried Chinese yam (Dioscorea batatas) flour containing dioscin on gastrointestinal function in rat model. Arch. Pharm. Res. 2006, 29, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Aumsuwan, P.; Khan, S.I.; Khan, I.A.; Ali, Z.; Avula, B.; Walker, L.A.; Shariat-Madar, Z.; Helferich, W.G.; Katzenellenbogen, B.S.; Dasmahapatra, A.K. The anticancer potential of steroidal saponin, dioscin, isolated from wild yam (Dioscorea villosa) root extract in invasive human breast cancer cell line MDA-MB-231 in vitro. Arch. Biochem. Biophys. 2016, 591, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Oyama, M.; Tokiwano, T.; Kawaii, S.; Yoshida, Y.; Mizuno, K.; Oh, K.; Yoshizawa, Y. Protodioscin, Isolated from the Rhizome of Dioscorea tokoro Collected in Northern Japan is the Major Antiproliferative Compound to HL-60 Leukemic Cells. Curr. Bioact. Compd. 2017, 13, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Pan, P.; Chad, S.; Jian, H.Y.; Hui, Z.; He, R.; Kiyoko, O.; Lin, S.W. Berries and other natural products in the pancreatic cancer chemoprevention in human clinical trials. J. Berry Res. 2017, 7, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.Y.; Lee, N.H.; Jung, D.; Lee, J.A.; Seo, C.S.; Lee, H.; Kim, J.H.; Shin, H.K. Protective effects of allantoin against ovalbumin (OVA)-induced lung inflammation in a murine model of asthma. Int. Immunopharmacol. 2010, 10, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Niu, C.S.; Chen, W.; Wu, H.T.; Cheng, K.C.; Wen, Y.J.; Lin, K.C.; Cheng, J.T. Decrease of plasma glucose by allantoin, an active principle of yam (Dioscorea spp.) in streptozotocin-induced diabetic rats. J. Agric. Food Chem. 2010, 58, 12031–12035. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.F.; Tsai, J.T.; Chem, L.J.; Wu, T.P.; Yang, J.J.; Yin, L.T.; Yang, Y.L.; Chiang, T.A.; Lu, H.L.; Wu, M.C. Antihypertensive action of allantoin in animals. Biomed. Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Ahn, Y.J.; Park, S.J.; Woo, H.; Lee, H.E.; Kwon, G.; Gao, Q.; Jang, D.S.; Ryu, J.H. Effects of allantoin on cognitive function and hippocampal neurogenesis. Food Chem. Toxicol. 2014, 64, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Song, M.Y.; Kang, S.Y.; Oh, T.W.; Kumar, R.V.; Jung, H.W.; Park, Y.K. The Roots of Atractylodes macrocephala Koidzumi Enhanced Glucose and Lipid Metabolism in C2C12 Myotubes via Mitochondrial Regulation. Evid. Based Complement. Altern. Med. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Beaudart, C.; Rizzoli, R.; Bruyère, O.; Reginster, J.Y.; Bive, E. Sarcopenia: Burden and challenges for public health. Arch. Public Health 2014, 72. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.C.; Ferng, L.H.A.; Huang, P.Y. Quantitative analysis of allantoin and allantonic acid in yam tuber, mucilage, skin and bulbil of the Dioscorea species. Food Chem. 2006, 94, 541–549. [Google Scholar] [CrossRef]
- Argilés, J.M.; Campos, N.; Lopez-Pedrosa, J.M.; Rueda, R.; Rodriguez-Mañas, L. Skeletal Muscle Regulates Metabolism via Interorgan Crosstalk: Roles in Health and Disease. J. Am. Med. Dir. Assoc. 2016, 17, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Sayer, A.A.; Robinson, S.M.; Patel, H.P.; Shavlakadze, T.; Cooper, C.; Grounds, M.D. New horizons in the pathogenesis, diagnosis and management of sarcopenia. Age Ageing 2013, 42, 145–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.Y.; Kwon, Q. Culinary plant and their potential impact on metabolic overload. Ann. N. Y. Acad. Sci. 2011, 1229, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Liang, X. Food therapy and medicinal diet therapy of traditional Chinese medicine. Clin. Nutr. Exp. 2018, 18, 1–5. [Google Scholar] [CrossRef]
- Komesaroff, P.A.; Black, C.V.; Cable, V.; Sudhir, K. Effects of wild yam extract on menopausal symptoms, lipids and sex hormones in healthy menopausal women. Climacteric 2001, 4, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Dumont, N.A.; Bentzinger, C.F.; Sincennes, M.C.; Rudnicki, M.A. Satellite Cells and Skeletal Muscle Regeneration. Compr. Physiol. 2015, 5, 1027–1059. [Google Scholar] [PubMed]
- Cole, N.J.; Hall, T.E.; Martin, C.L.; Chapman, M.A.; Kobiyama, A.; Nihei, Y.; Watabe, S.; Johnston, I.A. Temperature and the expression of myogenic regulatory factors (MRFs) and myosin heavy chain isoforms during embryogenesis in the common carp Cyprinus carpio L. J. Exp. Biol. 2004, 207, 4239–4248. [Google Scholar] [CrossRef] [PubMed]
- Saltiel, A.R.; Kahn, C.R. Insulin signalling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Massimiliano, G.; Francesca, G.; Jose, M.; Alvarez, S.; Mazzoni, L.; Tamara, Y.F.H.; Quiles, J.L.; Bullon, P.; Battino, M. AMPK as a New Attractive Therapeutic Target for Disease Prevention: The Role of Dietary Compounds AMPK and Disease Prevention. Curr. Drug Targets 2016, 17, 865–889. [Google Scholar]
- Pistollato, F.; Battino, M. Role of plant-based diets in the prevention and regression of metabolic syndrome and neurodegenerative diseases. Trends Food Sci. Technol. 2014, 40, 62–81. [Google Scholar] [CrossRef]
- Hardie, D.G.; Ross, F.A.; Hawley, S.A. AMPK: A nutrient and energy sensor that maintains energy homeostasis. Nat. Rev. Mol. Cell Biol. 2012, 13, 251–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, J.; Bruckbauer, A.; Zemel, M.B. Activation of the AMPK/Sirt1 pathway by a leucine–metformin combination increases insulin sensitivity in skeletal muscle, and stimulates glucose and lipid metabolism and increases life span in Caenorhabditis elegans. Metabolism 2016, 65, 1679–1691. [Google Scholar] [CrossRef] [PubMed]
- Cantó, C.; Auwerx, J. PGC-1α, SIRT1 and AMPK, an energy sensing network that controls energy expenditure. Curr. Opin. Lipidol. 2009, 20, 98–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taherzadeh-FardE, S.C.; Akkad, D.A.; Wieczorek, S.; Haghikia, A.; Chan, A.; Epplen, J.T.; Arning, L. PGC-1alpha downstream transcription factors NRF-1 and TFAM are genetic modifiers of Huntington disease. Mol. Neurodegener. 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Vytla, V.S.; Ochs, R.S. Metformin increases mitochondrial energy formation in L6 muscle cell culture. J. Biol. Chem. 2013, 288, 20369–20377. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Kim, M.A.; Moon, E.; Kim, S.Y.; Choi, S.Z.; Son, M.W.; Lee, K.R. Furostanolsaponins from the rhizomes of Dioscorea japonica and their effects on NGF induction. Bioorg. Med. Chem. Lett. 2011, 21, 2075–2078. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.W.; Um, S.H.; Kwak, J.H.; Park, H.J.; Kim, K.H.; Moon, E.Y.; Kwon, S.T.; Pyo, S. Suppression of adhesion molecule expression by phenanthrene-containing extract of bulbils of Chinese Yam in vascular smooth muscle cells through inhibition of MAPK, Akt and NF-κB. Food Chem. Toxicol. 2012, 50, 2792–2804. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Zhang, Y.; Su, C.; Zie, J. Determination of allantoin and adenosine in Rhizoma dioscoreae by HILIC-double wavelength HPLC. Northwest Pharm. J. 2018, 33, 334–336. [Google Scholar]
- Lin, K.C.; Yeh, L.R.; Chen, L.J.; Wen, Y.J.; Cheng, K.C.; Cheng, J.T. Plasma glucose-lowing action of allantoin is induced by activation of imidazoline I-2 receptors in streptozotocin-induced diabetic rats. Horm. Metab. Res. 2012, 44, 41–46. [Google Scholar] [PubMed]
- Umegaki, H. Sarcopenia and diabetes: Hyperglycemia is a risk factor for age-associated muscle mass and functional reduction. J. Diabetes Investig. 2015, 6, 623–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Accession No. | Sequence (5′→3′) | |
|---|---|---|---|
| MyHC | Forward | NM 001039545.2 | TGA ACT GGA GGG TGA GGT AG |
| Reverse | NM 001039545.2 | TTC GGT CTT CTT CTG TCT GG | |
| PGC1α | Forward | XM 006503779.3 | CAC CAA ACC CAC AGA AAA CAG |
| Reverse | XM 006503779.3 | GGG TCA GAG GAA GAG ATA AAG TTG | |
| NRF-1 | Forward | XM 017321445.1 | ACC CTC AGT CTC ACG ACT AT |
| Reverse | XM 017321445.1 | GAA CAC TCC TCA GAC CCT TAA C | |
| TFAM | Forward | XM 017313918.1 | CAC CCA GAT GCA AAA CTT TCA G |
| Reverse | XM 017313918.1 | CTG CTC TTT ATA CTT GCT CAC AG | |
| Sirt-1 | Forward | NM 001159589.2 | GAT CCT TCA GTG TCA TGG TT |
| Reverse | NM 001159589.2 | GAA GAC AAT CTC TGG CTT CA | |
| Gapdh | Forward | XM_017321385.1 | CAG CCT CGT CCC GTA GAC A |
| Reverse | XM_017321385.1 | CGC TCC TGG AAG ATG GTG AT |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, J.; Kang, S.Y.; Meng, X.; Kang, A.N.; Park, J.H.; Park, Y.-K.; Jung, H.W. Effects of Rhizome Extract of Dioscorea batatas and Its Active Compound, Allantoin, on the Regulation of Myoblast Differentiation and Mitochondrial Biogenesis in C2C12 Myotubes. Molecules 2018, 23, 2023. https://doi.org/10.3390/molecules23082023
Ma J, Kang SY, Meng X, Kang AN, Park JH, Park Y-K, Jung HW. Effects of Rhizome Extract of Dioscorea batatas and Its Active Compound, Allantoin, on the Regulation of Myoblast Differentiation and Mitochondrial Biogenesis in C2C12 Myotubes. Molecules. 2018; 23(8):2023. https://doi.org/10.3390/molecules23082023
Chicago/Turabian StyleMa, Junnan, Seok Yong Kang, Xianglong Meng, An Na Kang, Jong Hun Park, Yong-Ki Park, and Hyo Won Jung. 2018. "Effects of Rhizome Extract of Dioscorea batatas and Its Active Compound, Allantoin, on the Regulation of Myoblast Differentiation and Mitochondrial Biogenesis in C2C12 Myotubes" Molecules 23, no. 8: 2023. https://doi.org/10.3390/molecules23082023
APA StyleMa, J., Kang, S. Y., Meng, X., Kang, A. N., Park, J. H., Park, Y. -K., & Jung, H. W. (2018). Effects of Rhizome Extract of Dioscorea batatas and Its Active Compound, Allantoin, on the Regulation of Myoblast Differentiation and Mitochondrial Biogenesis in C2C12 Myotubes. Molecules, 23(8), 2023. https://doi.org/10.3390/molecules23082023