
Flowering Stage and Daytime Affect Scent Emission of Malus ioensis “Prairie Rose”


Abstract
:1. Introduction
2. Results
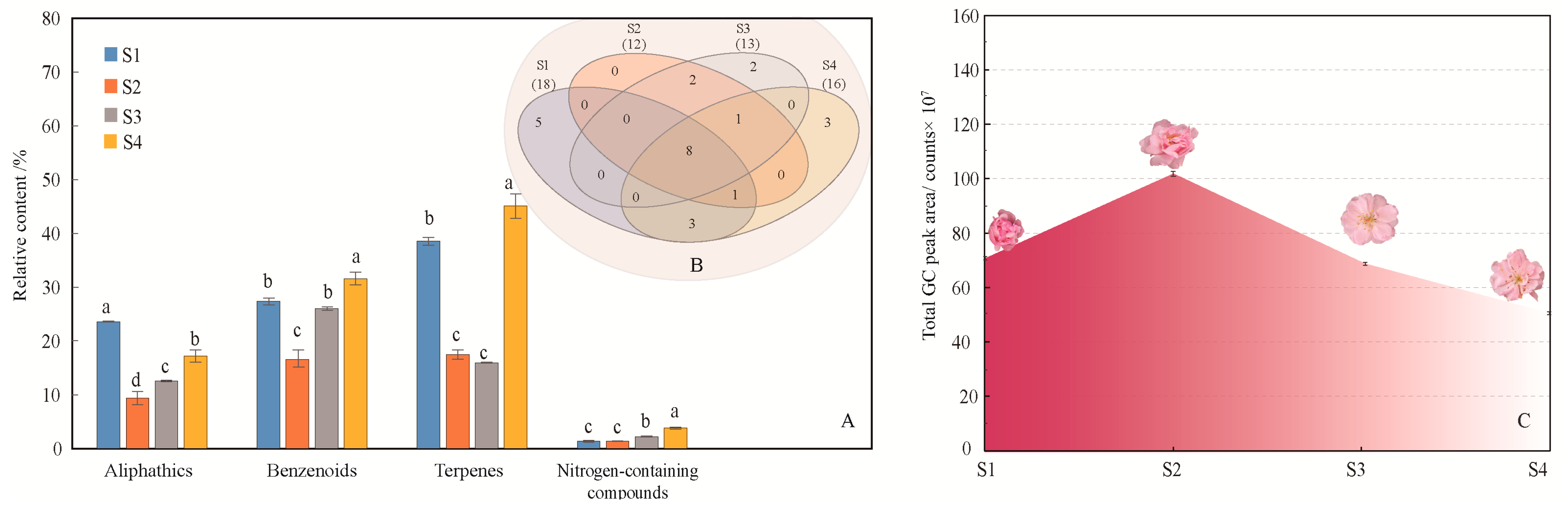
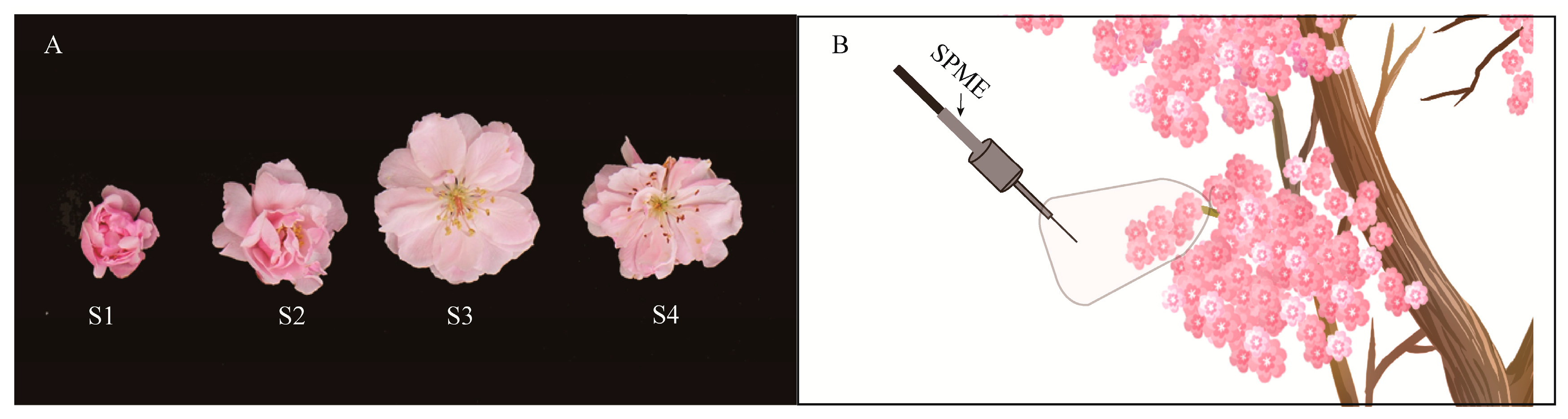
2.1. Volatile Compounds Emission Characteristic of Different Flowering Stages
2.1.1. Identification and Comparison of Volatile Compounds of Flowers at Different Flowering Stages
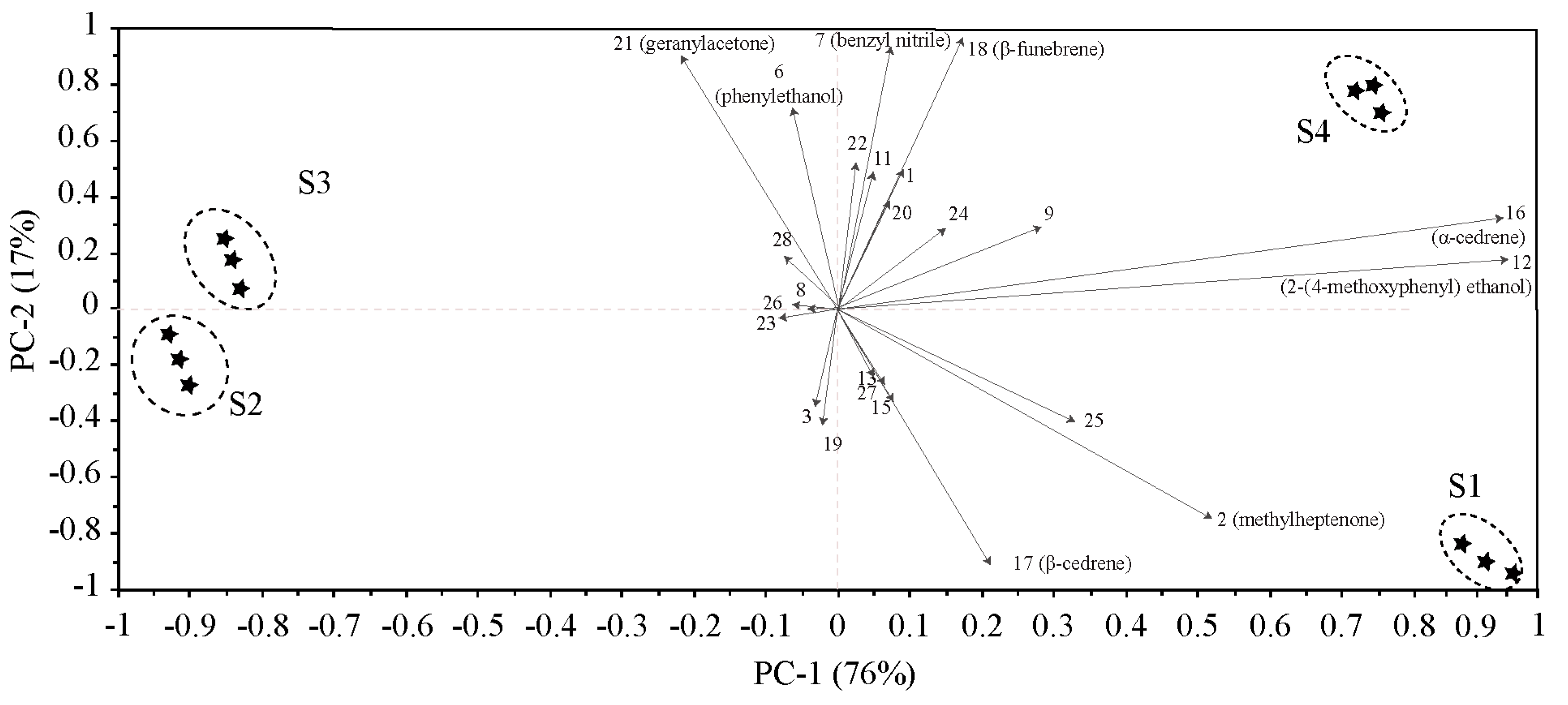
2.1.2. Difference Analysis of Flower Volatiles Based on Bray-Curtis Dissimilarity Analysis and Principal Component Analysis (PCA)
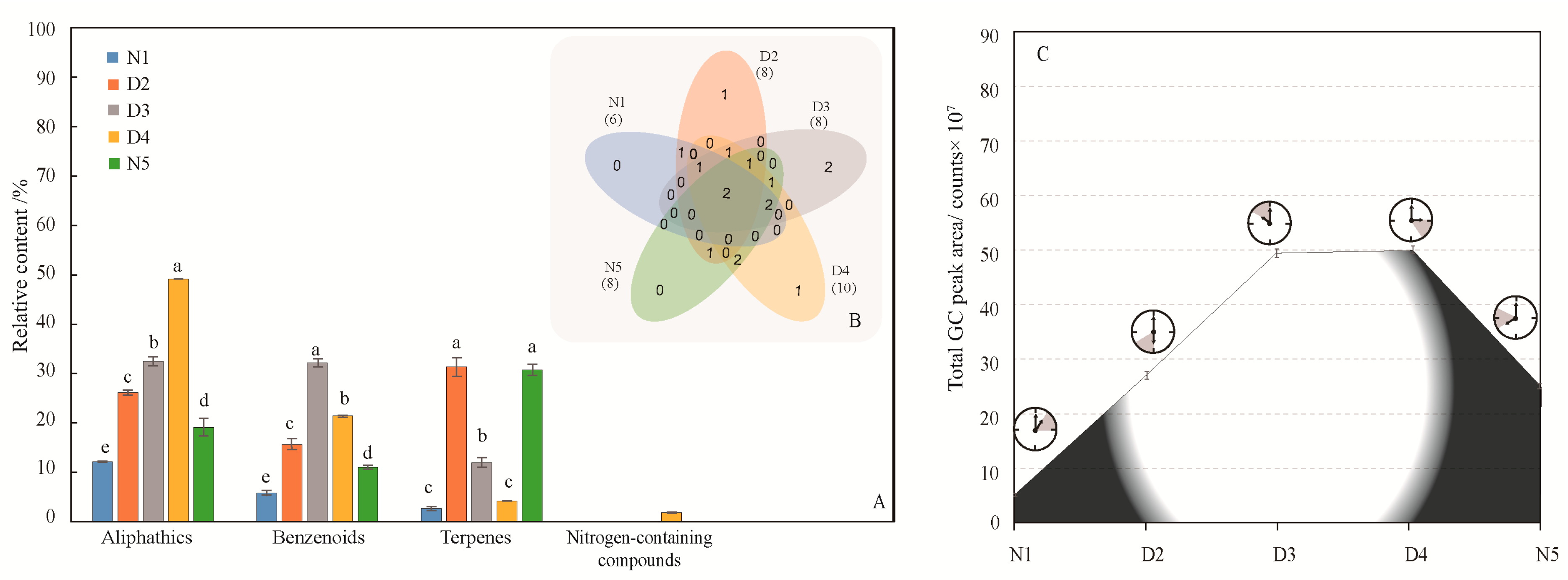
2.2. Diurnal and Nocturnal Release Rhythm of M. ioensis “Prairie Rose”
2.2.1. Identification and Comparison of Volatile Compounds at Different Times
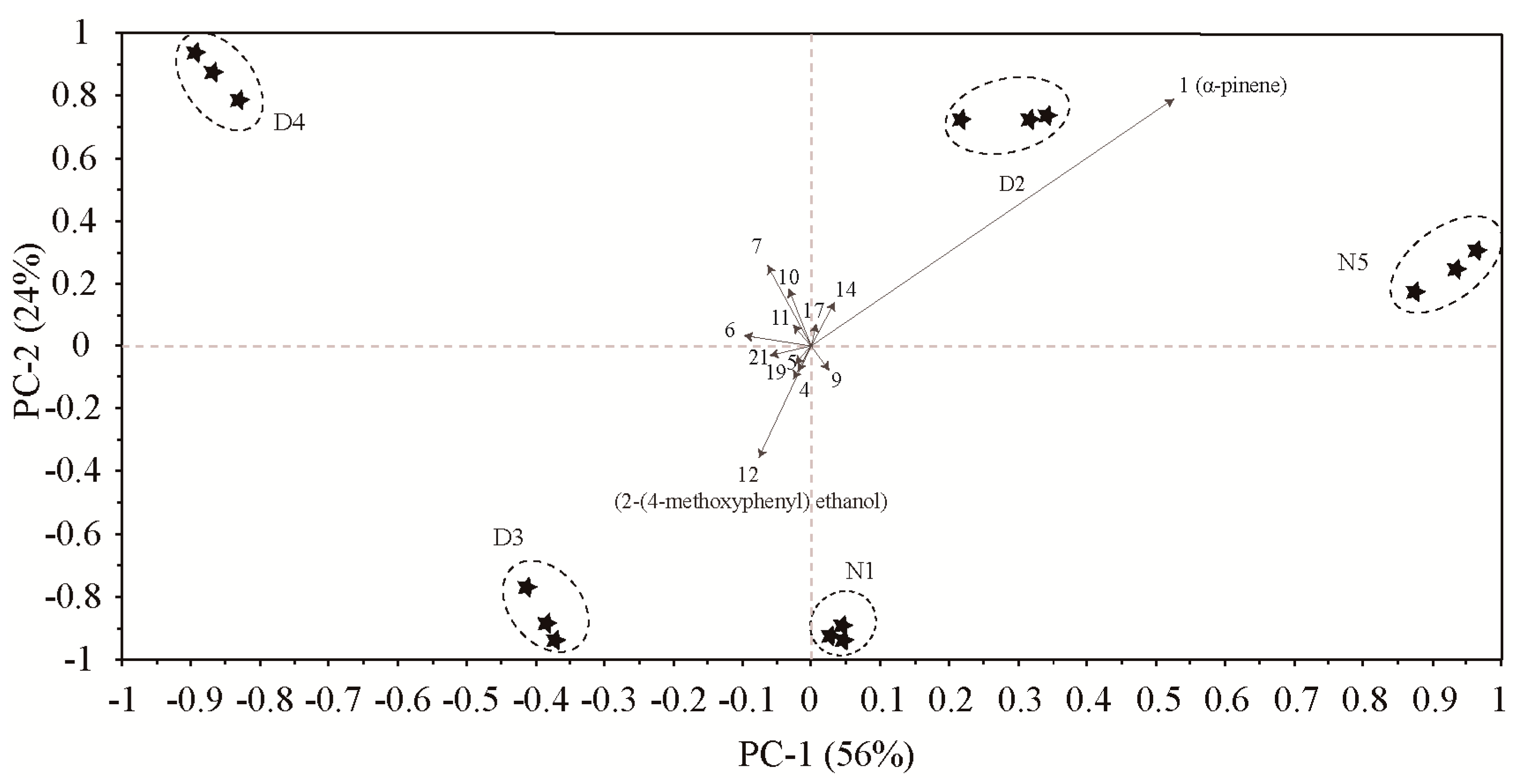
2.2.2. Difference Analysis of Volatiles in Different Flower Parts Based on Bray-Curtis Dissimilarity Analysis and Principal Component Analysis
2.3. Floral Scent Properties of M. ioensis “Prairie Rose”
3. Discussion
3.1. Flower Characteristics of M. ioensis “Prairie Rose”
3.2. The Release Dynamics of the Floral Scent of M. ioensis “Prairie Rose”
3.3. Limitations of Floral Collection Methods
4. Materials and Methods
4.1. Plant Materials
4.2. Experimental Design
4.3. SPME-GC-MS Analysis
4.4. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Knudsen, J.T.; Eriksson, R.; Gershenzon, J.; Stahl, B. Diversity and distribution of floral scent. Bot. Rev. 2006, 72, 1–120. [Google Scholar] [CrossRef]
- Dudareva, N.; Piechulla, B.; Pichersky, E. Biogenesis of floral scents. In Horticultural Reviews; Janick, J., Ed.; John Wiley & Sons: New York, NY, USA, 1999; Volume 24, pp. 31–54. [Google Scholar]
- Sexton, R.; Stopford, A.P.; Moodie, W.T.; Porter, A.E.A. Aroma production from cut sweet pea flowers (Lathyrus odoratus): The role of ethylene. Physiol. Plant. 2005, 124, 381–389. [Google Scholar] [CrossRef]
- Dudareva, N.; Negre, F.; Nagegowda, D.A.; Orlova, I. Plant volatiles: Recent advances and future perspectives. Crit. Rev. Plant Sci. 2006, 25, 417–440. [Google Scholar] [CrossRef]
- Vainstein, A.; Lewinsohn, E.; Pichersky, E.; Weiss, D. Floral fragrance. New inroads into an old commodity. Plant. Physiol. 2001, 127, 1383–1389. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, J.T.; Tollsten, L.; Bergström, L.G. Floral scents—A checklist of volatile compounds isolated by head-space techniques. Phytochemistry 1993, 33, 253–280. [Google Scholar] [CrossRef]
- Fiala, J.L. Flowering Crabapples: The Genus Malus; Timber Press: Portland, OR, USA, 1994; pp. 105–273. [Google Scholar]
- Li, Y.N. Researches of Germplasm Resources of Malus Mill; China Agriculture Press: Beijing, China, 2001; pp. 3–9, 181–183, 315–335. [Google Scholar]
- Zhao, J.; Wang, R.; Huang, C.X.; Mao, Z.Q.; Guo, L.; Shen, X. Taxonomic analysis of volatiles emitted by ornamental crabapple flowers. Acta Ecol. Sin. 2014, 34, 213–218. [Google Scholar] [CrossRef]
- Chu, A.X.; Tang, G.G. Cultivar investigation and classification of Malus halliana Koehne in Henan Province. Acta Agric. Univ. Jiangxiensis 2008, 30, 1090–1096. [Google Scholar] [CrossRef]
- Muzher, B.M.; Younis, R.A.A.; El-Halabi, O.; Ismail, O.M. Genetic identification of some Syrian local apple (Malus sp.) cultivars using molecular markers. Res. J. Agric. Biol. Sci. 2007, 3, 704–713. [Google Scholar]
- Ulukan, H. The evolution of cultivated plant species: Classical plant breeding versus genetic engineering. Plant. Sys. Evol. 2009, 280, 133–142. [Google Scholar] [CrossRef]
- Li, W. HS-SPME-GC-MS analysis of volatile constituents from the flowers and leaves of Malus baccata (Linn.) Borkh. Nat. Prod. Res. Dev. 2012, 24, 490–493. [Google Scholar] [CrossRef]
- Fan, J.; Zhang, W.; Zhou, T.; Zhang, D.; Zhang, D.; Zhang, L.; Wang, G.; Cao, F. Discrimination of Malus taxa with different scent intensities using electronic nose and gas chromatography–mass spectrometry. Sensors 2018, 18, 3429. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.X.; Wei, H.L.; Jiang, Z.H.; Cao, F.L.; Tang, G.G. Studies on flowering phenological characteristics of ornamental crabapple cultivar group. Acta Hortic. Sin. 2014, 41, 713–725. [Google Scholar]
- Zhang, W.X.; Qiu, J.; Jiang, Z.H.; Wei, H.L.; Cao, F.L.; Tang, G.G. Studies on dynamic characteristics of the pigment components of ornamental crabapple cultivars groups in flowering process. Acta Hortic. Sin. 2014, 41, 1145–1156. [Google Scholar]
- Zhang, W.X.; Jiang, Z.H.; Qiu, J.; Wei, H.L.; Cao, F.L. Studies on sequence dynamic distribution pattern of flower color parameters of ornamental crabapple. Acta Hortic. Sin. 2013, 40, 505–514. [Google Scholar]
- Zhang, W.X.; Zhao, M.M.; Fan, J.J.; Zhou, T.; Chen, Y.X.; Cao, F.L. Study on relationship between pollen exine ornamentation pattern and germplasm evolution in flowering crabapple. Sci. Rep. 2017, 7, 39759. [Google Scholar] [CrossRef] [Green Version]
- Dudareva, N.; Pichersky, E.; Gershenzon, J. Biochemistry of plant volatiles. Plant. Physiol. 2004, 135, 1893–1902. [Google Scholar] [CrossRef]
- Li, Y.; Ma, H.; Wan, Y.; Li, T.; Liu, X.; Sun, Z.; Li, Z. Volatile organic compounds emissions from Luculia pinceana flower and its changes at different stages of flower development. Molecules 2016, 21, 531. [Google Scholar] [CrossRef] [PubMed]
- Deng, B.; Cao, Y.; Fang, S.; Shang, X.; Yang, W.; Qian, C. Variation and stability of growth and leaf flavonoid content in Cyclocarya paliurus across environments. Ind Crop. Prod. 2015, 76, 386–393. [Google Scholar] [CrossRef]
- Pichersky, E.; Raguso, R.A.; Lewinsohn, E.; Croteau, R. Floral scent production in Clarkia (Onagraceae) (I. localization and developmental modulation of monoterpene emission and linalool synthase activity). Plant. Physiol. 1994, 106, 1533–1540. [Google Scholar] [CrossRef]
- Effmert, U.; Große, J.; Röse, U.S.; Ehrig, F.; Kägi, R.; Piechulla, B. Volatile composition, emission pattern, and localization of floral scent emission in Mirabilis jalapa (Nyctaginaceae). Am. J. Bot. 2005, 92, 2–12. [Google Scholar] [CrossRef]
- Van Gemert, L.J. Odour Thresholds: Compilations of Odour Threshold Values in Air, Water and Other Media, 2nd ed.; Oliemans Punter & Partners BA: Utrecht, The Netherlands, 2011. [Google Scholar]
- Bicchi, C.; Joulain, D. Review headspace-gas chromatographic analysis of medicinal and aromatic plants and flowers. Flavour Fragr. 1990, 5, 131–145. [Google Scholar] [CrossRef]
- Yoshioka, P.M. Misidentification of the Bray-Curtis similarity index. Mar. Ecol. Prog. Ser. 2008, 368, 309–310. [Google Scholar] [CrossRef] [Green Version]
- Bult, J.H.; Schifferstein, H.N.; Roozen, J.P.; Boronat, E.D.; Voragen, A.G.; Kroeze, J.H. Sensory evaluation of character impact components in an apple model mixture. Chem. Senses 2002, 27, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Laing, D.G.; Eddy, A.; Francis, G.W.; Stephens, L. Evidence for the temporal processing of odor mixtures in humans. Brain Res. 1994, 651, 317–328. [Google Scholar] [CrossRef]
- Wang, L.M.; Li, M.T.; Jin, W.W.; Li, S.; Zhang, S.Q.; Yu, L.J. Variations in the components of Osmanthus fragrans lour. essential oil at different stages of flowering. Food Chem. 2009, 114, 233–236. [Google Scholar] [CrossRef]
- Zhang, J.J.; Kang, W. Volatiles from Flowers of Lagerstroemia caudata by HS-SPME-GC-MS. Chem. Nat. Compd. 2014, 50, 933–934. [Google Scholar] [CrossRef]
- Li, Z.G.; Lee, M.R.; Shen, D.L. Analysis of volatile compounds emitted from fresh Syringa oblata flowers in different florescence by headspace solid-phase microextraction–gas chromatography–mass spectrometry. Anal. Chim. Acta 2006, 576, 43–49. [Google Scholar] [CrossRef]
- Scalliet, G.; Piola, F.; Douady, C.J.; Réty, S.; Raymond, O.; Baudino, S.; Bordji, K.; Bendahmane, M.; Dumas, C.; Cock, J.M.; et al. Scent evolution in Chinese roses. Proc. Natl. Acad. Sci. USA 2008, 105, 5927–5932. [Google Scholar] [CrossRef] [Green Version]
- Nowak, R. Chemical composition of hips essential oils of some Rosa L. species December 13, 2004. Z. Naturforsch. C 2005, 60, 369–378. [Google Scholar] [CrossRef]
- Kong, Y.; Sun, M.; Pan, H.T.; Zhang, Q.X. Advances in metabolism and regulation of floral scent. J. Beijing For. Univ. 2012, 34, 149–154. [Google Scholar]
- Baldermann, S.; Yang, Z.; Sakai, M.; Fleischman, P.; Watanabe, N. Volatile constituents in the scent of roses. Floric. Ornam. Biotechnol. 2009, 3, 89–97. [Google Scholar]
- Berglund, B.; Olsson, M.J. Odor-intensity interaction in binary and ternary mixtures. Percept. Psychophys. 1993, 53, 475–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudareva, N.; Pichersky, E. Biochemical and molecular genetic aspects of floral scents. Plant. Physiol. 2000, 122, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Yakir, E.; Hilman, D.; Harir, Y.; Green, R.M. Regulation of output from the plant circadian clock. FEBS J. 2007, 274, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Mc Tavish, H.S.; Davies, N.W.; Menary, R.C. Emission of Volatiles from brown boronia flowers: Some comparative observations. Ann. Bot. 2000, 86, 347–354. [Google Scholar] [CrossRef]
- Dudareva, N.; Murfitt, L.M.; Mann, C.J.; Gorenstein, N.; Kolosova, N.; Kish, C.M.; Bonham, C.; Wood, K. Developmental regulation of methyl benzoate biosynthesis and emission in snapdragon flowers. Plant Cell Online 2000, 12, 949–961. [Google Scholar] [CrossRef] [PubMed]
- Kolosova, N.; Gorenstein, N.; Kish, C.M.; Dudareva, N. Regulation of circadian methyl benzoate emission in diurnally and nocturnally emitting plants. Plant Cell 2001, 13, 2333–2347. [Google Scholar] [CrossRef]
- Shi, T.; Yang, X.; Wang, L. Study on the aroma component emission pattern of Osmanthus fragrans ‘Boye Jingui’. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2018, 42, 97–104. [Google Scholar] [CrossRef]
- Fu, L.S. Study on the Biological Characteristics of High-Efficiency Pollination Tree in Apple Flowering Stage. Ph.D. Thesis, Shangdong Agriculture University, Shandong, China, 2010. [Google Scholar]
- Zhao, X.; Yuan, Z.; Fang, Y.; Yin, Y.; Feng, L. Characterization and evaluation of major anthocyanins in pomegranate (Punica granatum L.) peel of different cultivars and their development phases. Eur. Food Res. Technol. 2013, 236, 109–117. [Google Scholar] [CrossRef]
- Wang, S.A.; Yang, R.; Wang, P.; Wang, Q.; Li, L.; Li, Y.; Yin, Z. ‘Jinhuang’Yellow-leaf Crape Myrtle. HortScience 2016, 51, 595–596. [Google Scholar] [CrossRef]
- Delano-Frier, J.P.; Pearce, G.; Huffaker, A.; Stratmann, J.W. Systemic Wound Signaling in Plants. In Long-Distance Systemic Signaling and Communication in Plants; František, B., Ed.; Springer: Berlin, Germany, 2013; Volume 19, pp. 323–362. [Google Scholar]
- Stashenko, E.E.; Martínez, J.R. Sampling flower scent for chromatographic analysis. J. Sep. Sci. 2008, 31, 2022–2031. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Pawliszyn, J. Solid-phase microextraction field sampler. Anal. Chem. 2004, 76, 6823–6828. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Time/min | Compounds | CAS | LRI | LRI a | Probability/% | ODT b/ppm | Odor Characteristic c | Relative Content/% | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bud | Initial Flowering | Flowering | End Flowering | 1:00–3:00 | 6:00–8:00 | 10:00–12:00 | 15:00–17:00 | 20:00–22:00 | |||||||||
| 1 | 6.93 | α-pinene | 80-56-8 | 877 | 907 | 87.5–92.1 | 0.12–1.01 | fruity, sweet, pine | -- | -- | -- | 2.62 ± 0.21 | -- | 19.89 ± 1.12b | -- | -- | 28.53 ± 1.38a |
| 2 | 9.43 | methylheptenone | 110-93-0 | 937 | 942 | 81.6–93.4 | 0.0189 | mushroom, earthy, woody | 16.36 ± 0.23a | 6.92 ± 0.9d | 8.08 ± 0.26c | 11.72 ± 0.61b | 2.35 ± 0.24d | 14.65 ± 0.97b | 11.5 ± 1.06c | 31.54 ± 1.34a | -- |
| 3 | 9.92 | 4-methylanisole | 104-93-8 | 972 | 989 | 84.8–91.9 | 0.0029 | 1.83 ± 0.06a | 1.39 ± 0.29c | 1.48 ± 0.09b | -- | ||||||
| 4 | 12.15 | linalool | 78-70-6 | 1079 | 1082 | 83.8 | 0.0015 | floral, woody, fresh | -- | -- | 3.61 ± 0.34 | -- | -- | ||||
| 5 | 12.32 | nonanal | 124-19-6 | 1098 | 1098 | 76.4–89.2 | 0.0031 | floral, sweet, citrus | 1.46 ± 0.13 | -- | -- | -- | 4.83 ± 0.19b | -- | 5.6 ± 0.5b | 6.88 ± 0.76a | 7.34 ± 0.66a |
| 6 | 12.58 | phenylethanol | 60-12-8 | 1165 | 1150 | 78.2–91.5 | 0.012 | rose, honey, fragrant | 6.02 ± 0.33c | 3.76 ± 0.2d | 10.59 ± 0.68a | 7.51 ± 0.48b | -- | 5.36 ± 0.9b | 8.68 ± 1.5a | 5.21 ± 0.13b | -- |
| 7 | 12.77 | benzyl nitrile | 140-29-4 | 1178 | 1160 | 82.8–93.6 | 1 | herbal, floral | 2.83 ± 0.06c | 2.81 ± 0.31c | 5.18 ± 0.24b | 7.41 ± 0.47a | -- | -- | -- | 7.24 ± 0.7a | 2.62 ± 0.51b |
| 8 | 15.57 | cuminaldehyde | 122-03-2 | 1212 | 1214 | 94.7 | 0.14 | sharp, woody | -- | -- | 1.2 ± 0.09 | -- | |||||
| 9 | 15.6 | dodecane | 112-40-3 | 1231 | 1200 | 83.2–86.5 | 0.77 | fusel-like | 3.06 ± 0.09b | -- | -- | 4.4 ± 0.37a | 5.02 ± 0.32c | 7.59 ± 0.53b | 9.64 ± 1.64a | 4.25 ± 0.71c | 7.49 ± 0.39b |
| 10 | 15.75 | decanal | 112-31-2 | 1219 | 1220 | 79.9–95.0 | 0.003 | green, floral, lemon, | -- | -- | -- | 5.03 ± 0.43a | 2.48 ± 0.39b | ||||
| 11 | 18.87 | (2-nitroethyl) benzene | 6125-24-2 | 1321 | 1304 | 80.6–91.2 | 0.002 | sweet, floral, spicy | 1.36 ± 0.19c | 1.41 ± 0.03c | 2.23 ± 0.24b | 3.83 ± 0.18a | -- | -- | -- | 1.84 ± 0.15 | -- |
| 12 | 21.49 | 2-(4-methoxyphenyl) ethanol | 702-23-8 | 1381 | 1374 | 79.8–87.9 | -- | fresh citrus | 16.56 ± 0.56a | 3.58 ± 0.43c | 5.25 ± 0.62b | 16.36 ± 1.21a | 6.56 ± 0.32d | 9.16 ± 0.22b | 21.96 ± 1.08a | 7.84 ± 0.41c | 8.1 ± 0.41c |
| 13 | 21.79 | texanol | 77-68-9 | 1380 | 1380 | 90.3 | -- | 1.26 ± 0.04 | -- | -- | -- | ||||||
| 14 | 22.19 | (Z)-3-hexenyl hexanoate | 31501-11-8 | 1389 | 1380 | 84.9–89.5 | 0.0052 | fruity, green | 2.66 ± 0.45b | 9.1 ± 0.25a | -- | -- | -- | ||||
| 15 | 22.32 | β-elemene | 515-13-9 | 1443 | 1429 | 89.4 | -- | herbal | 1.70 ± 0.20 | -- | -- | -- | |||||
| 16 | 23.48 | α-cedrene | 469-61-4 | 1453 | 1444 | 88.4–90.2 | 0.00003 | woody | 11.71 ± 0.36a | -- | -- | 12.41 ± 1.29a | |||||
| 17 | 23.82 | β-cedrene | 546-28-1 | 1456 | 1447 | 82.6–88.4 | 0.00003 | woody | 5.02 ± 0.14 | -- | -- | -- | -- | 2.62 ± 0.59 | -- | -- | -- |
| 18 | 23.96 | β-funebrene | 79120-98-2 | 1457 | 1434 | 92.6 | -- | -- | -- | -- | 5.00 ± 0.19 | ||||||
| 19 | 24.1 | α-ionone | 127-41-3 | 1434 | 1434 | 83.3–95.1 | 0.001–0.006 | violet, woody, fruity | 4.39 ± 0.5a | 3.8 ± 0.56a | 3.64 ± 0.09a | 2.32 ± 0.16b | -- | -- | 2.61 ± 0.52 | -- | -- |
| 20 | 24.45 | cis-thujopsene | 470-40-6 | 1462 | 1456 | 88.2 | -- | -- | -- | -- | 2.05 ± 0.29 | ||||||
| 21 | 25.2 | geranylacetone | 689-67-8 | 1456 | 1456 | 79.3–89.8 | 0.06 | fresh, floral, rose, fruity | 4.92 ± 0.4b | 10.7 ± 1.26a | 9.64 ± 0.33a | 10.19 ± 0.76a | 5.77 ± 0.81a | 4.16 ± 0.21b | 2.24 ± 0.23c | ||
| 22 | 27.09 | β-ionone | 79-77-6 | 1493 | 1491 | 88.1–93.9 | 0.001–0.006 | earthy, woody | 1.3 ± 0.09c | 1.7 ± 0.03c | 2.71 ± 0.08b | 3.95 ± 0.74a | |||||
| 23 | 27.88 | pentadecane | 629-62-9 | 1492 | 1500 | 84.6–91.5 | 0.5 | mild green, fusel-like | -- | 1.16 ± 0.16a | 1.03 ± 0.06a | -- | |||||
| 24 | 28.36 | α-farnesene | 502-61-4 | 1496 | 1496 | 80.2–87.9 | 2 | woody | 1.96 ± 0.37b | 1.3 ± 0.14c | -- | 3.58 ± 0.16a | |||||
| 25 | 32.09 | cedrol | 77-53-2 | 1528 | 1573 | 85.4–90.5 | 0.00013 | sweet | 5.42 ± 0.31a | -- | -- | 2.99 ± 0.30b | |||||
| 26 | 38.95 | octacosane | 630-02-4 | 2802 | 2800 | 85.9 | -- | -- | -- | 1.80 ± 0.12 | -- | ||||||
| 27 | 39.14 | heneicosane | 629-94-7 | 2082 | 2100 | 90.3 | -- | 1.46 ± 0.33 | -- | -- | -- | ||||||
| 28 | 39.58 | methyl hexadecanoate | 112-39-0 | 1920 | 1925 | 86.8–93.1 | >2 | faint, sweet | -- | 1.33 ± 0.16b | 1.69 ± 0.02a | 1.10 ± 0.12b | 20.51 ± 0.95a | 5.25 ± 0.23c | -- | 5.35 ± 0.31c | 9.73 ± 0.47b |
| Flowering Stage | S1 | S2 | S3 | S4 |
|---|---|---|---|---|
| S1 | 0 | |||
| S2 | 0.52 | 0 | ||
| S3 | 0.52 | 0.21 | 0 | |
| S4 | 0.27 | 0.48 | 0.41 | 0 |
| Times of Day | N1 | D2 | D3 | D4 | N5 |
|---|---|---|---|---|---|
| N1 | 0 | ||||
| D2 | 0.65 | 0 | |||
| D3 | 0.61 | 0.49 | 0 | ||
| D4 | 0.63 | 0.55 | 0.47 | 0 | |
| N5 | 0.60 | 0.44 | 0.63 | 0.59 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, J.; Zhang, W.; Zhang, D.; Wang, G.; Cao, F. Flowering Stage and Daytime Affect Scent Emission of Malus ioensis “Prairie Rose”. Molecules 2019, 24, 2356. https://doi.org/10.3390/molecules24132356
Fan J, Zhang W, Zhang D, Wang G, Cao F. Flowering Stage and Daytime Affect Scent Emission of Malus ioensis “Prairie Rose”. Molecules. 2019; 24(13):2356. https://doi.org/10.3390/molecules24132356
Chicago/Turabian StyleFan, Junjun, Wangxiang Zhang, Donglin Zhang, Guibin Wang, and Fuliang Cao. 2019. "Flowering Stage and Daytime Affect Scent Emission of Malus ioensis “Prairie Rose”" Molecules 24, no. 13: 2356. https://doi.org/10.3390/molecules24132356
APA StyleFan, J., Zhang, W., Zhang, D., Wang, G., & Cao, F. (2019). Flowering Stage and Daytime Affect Scent Emission of Malus ioensis “Prairie Rose”. Molecules, 24(13), 2356. https://doi.org/10.3390/molecules24132356