
Identification and Characterization of trans-Isopentenyl Diphosphate Synthases Involved in Herbivory-Induced Volatile Terpene Formation in Populus trichocarpa


 , and
, and
Abstract
:1. Introduction
2. Results
2.1. The trans-IDS Gene Family in P. trichocarpa Possesses a Single FPPS Gene and Four Putative GPPS Genes
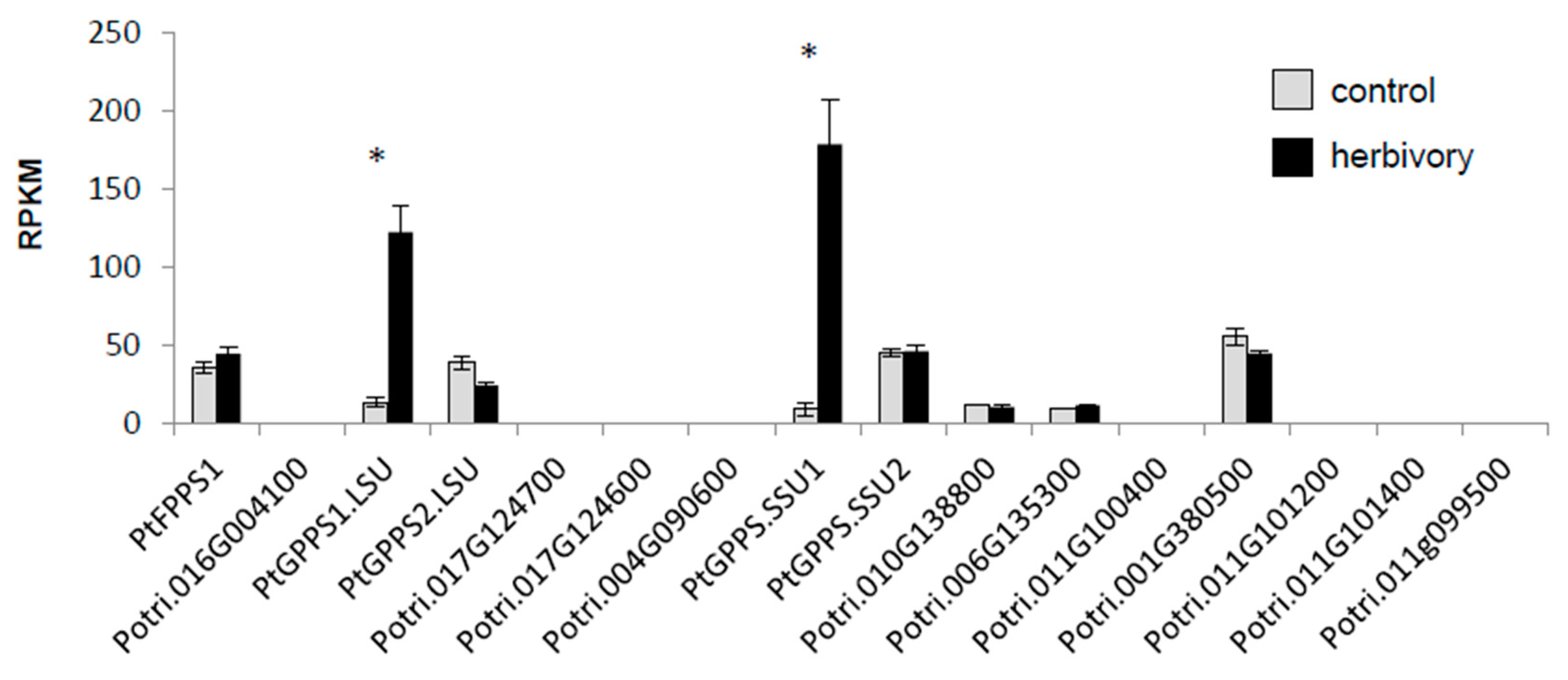
2.2. PtGPPS1.LSU and PtGPPS.SSU1 are Induced Upon Herbivory
2.3. PtFPPS1 Produces FPP While PtGPPS1.LSU and PtGPPS2.LSU Have GPPS and GGPPS Activity In Vitro
2.4. The Small Subunit PtGPPS.SSU1 Enhances the GPPS Activity of PtGPPS1.LSU and PtGPPS2.LSU In Vitro
3. Discussion
4. Material and Methods
4.1. Plants and Insects
4.2. RNA Extraction and Reverse Transcription
4.3. Identification and Isolation of trans-IDS Genes
4.4. Heterologous Expression and Enzyme Assays
4.5. LC-MS/MS Analysis of GPP, FPP, and GGPP
4.6. Gene Expression Analysis (RNAseq and qRT-PCR)
4.7. Sequence Analysis and Tree Reconstruction
4.8. Statistical Analysis
4.9. Accession Numbers
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Unsicker, S.B.; Kunert, G.; Gershenzon, J. Protective perfumes: The role of vegetative volatiles in plant defense against herbivores. Curr. Opin. Plant Biol. 2009, 12, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Riedlmeier, M.; Ghirardo, A.; Wenig, M.; Knappe, C.; Koch, K.; Georgii, E.; Dey, S.; Parker, J.E.; Schnitzler, J.P.; Vlot, A.C. Monoterpenes support systemic acquired resistance within and between plants. Plant Cell 2017, 29, 1440–1459. [Google Scholar] [CrossRef] [PubMed]
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Nagel, R.; Schmidt, A.; Peters, R.J. Isoprenyl diphosphate synthases: The chain length determining step in terpene biosynthesis. Planta 2018. [Google Scholar] [CrossRef] [PubMed]
- Closa, M.; Vranova, E.; Bortolotti, C.; Bigler, L.; Arro, M.; Ferrer, A.; Gruissem, W. The Arabidopsis thaliana FPP synthase isozymes have overlapping and specific functions in isoprenoid biosynthesis, and complete loss of FPP synthase activity causes early developmental arrest. Plant J. 2010, 63, 512–525. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Schalk, M.; Clark, A.; Miles, R.B.; Coates, R.; Chappell, J. Redirection of cytosolic or plastidic isoprenoid precursors elevates terpene production in plants. Nat. Biotechnol. 2006, 24, 1441–1447. [Google Scholar] [CrossRef]
- Liang, P.-H.; Ko, T.-P.; Wang, A.H.J. Structure, mechanism and function of prenyltransferases. Eur. J. Biochem. 2002, 269, 3339–3354. [Google Scholar] [CrossRef]
- Vandermoten, S.; Haubruge, E.; Cusson, M. New insights into short-chain prenyltransferases: Structural features, evolutionary history and potential for selective inhibition. Cell. Mol. Life Sci. 2009, 66, 3685–3695. [Google Scholar] [CrossRef]
- Wang, K.C.; Ohnuma, S.-I. Isoprenyl diphosphate synthases. Biochim. Biophys. Acta 2000, 1529, 33–48. [Google Scholar] [CrossRef]
- Tarshis, L.C.; Yan, M.; Poulter, C.D.; Sacchettini, J.C. Crystal structure of recombinant farnesyl diphosphate synthase at 2.6-Ang. Resolution. Biochemistry 1994, 33, 10871–10877. [Google Scholar] [CrossRef]
- Hosfield, D.J.; Zhang, Y.; Dougan, D.R.; Broun, A.; Tari, L.W.; Swanson, R.V.; Finn, J. Structural basis for bisphosphonate-mediated inhibition of isoprenoid biosynthesis. J. Biol. Chem. 2004, 279, 8526–8529. [Google Scholar] [CrossRef]
- Kavanagh, K.L.; Dunford, J.E.; Bunkoczi, G.; Russell, R.G.; Oppermann, U. The crystal structure of human geranylgeranyl pyrophosphate synthase reveals a novel hexameric arrangement and inhibitory product binding. J. Biol. Chem. 2006, 281, 22004–22012. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.H.; Guo, R.T.; Ko, T.P.; Wang, A.H.; Liang, P.H. Crystal structure of Type-III geranylgeranyl pyrophosphate synthase from Saccharomyces cerevisiae and the mechanism of product chain length determination. J. Biol. Chem. 2006, 281, 14991–15000. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.H.; Hsieh, F.L.; Ko, T.P.; Teng, K.H.; Liang, P.H.; Wang, A.H. Structure of a heterotetrameric geranyl pyrophosphate synthase from mint (Mentha piperita) reveals intersubunit regulation. Plant Cell 2010, 22, 454–467. [Google Scholar] [CrossRef] [PubMed]
- Dickschat, J.S. Isoprenoids in three-dimensional space: The stereochemistry of terpene biosynthesis. Nat. Prod. Rep. 2011, 28, 1917–1936. [Google Scholar] [CrossRef] [PubMed]
- Tholl, D.; Kish, C.M.; Orlova, I.; Sherman, D.; Gershenzon, J.; Pichersky, E.; Dudareva, N. Formation of monoterpenes in Antirrhinum majus and Clarkia breweri flowers involves heterodimeric geranyl diphosphate synthases. Plant Cell 2004, 16, 977–992. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Dixon, R.A. Heterodimeric geranyl(geranyl)diphosphate synthase from hop (Humulus lupulus) and the evolution of monoterpene biosynthesis. Proc. Natl. Acad. Sci. USA 2009, 106, 9914–9919. [Google Scholar] [CrossRef] [PubMed]
- Orlova, I.; Nagegowda, D.A.; Kish, C.M.; Gutensohn, M.; Maeda, H.; Varbanova, M.; Fridman, E.; Yamaguchi, S.; Hanada, A.; Kamiya, Y.; et al. The small subunit of snapdragon geranyl diphosphate synthase modifies the chain length specificity of tobacco geranylgeranyl diphosphate synthase in planta. Plant Cell 2009, 21, 4002–4017. [Google Scholar] [CrossRef] [PubMed]
- Nagegowda, D.A. Plant volatile terpenoid metabolism: Biosynthetic genes, transcriptional regulation and subcellular compartmentation. FEBS Lett. 2010, 584, 2965–2973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danner, H.; Boeckler, G.A.; Irmisch, S.; Yuan, J.S.; Chen, F.; Gershenzon, J.; Unsicker, S.B.; Köllner, T.G. Four terpene synthases produce major compounds of the gypsy moth feeding-induced volatile blend of Populus trichocarpa. Phytochemistry 2011, 72. [Google Scholar] [CrossRef] [PubMed]
- Irmisch, S.; Jiang, Y.; Chen, F.; Gershenzon, J.; Köllner, T.G. Terpene synthases and their contribution to herbivore-induced volatile emission in western balsam poplar (Populus trichocarpa). BMC Plant Biol. 2014, 14. [Google Scholar] [CrossRef] [PubMed]
- Irmisch, S.; McCormick, A.C.; Boeckler, G.A.; Schmidt, A.; Reichelt, M.; Schneider, B.; Block, K.; Schnitzler, J.P.; Gershenzon, J.; Unsicker, S.B.; et al. Two herbivore-induced cytochrome P450 enzymes CYP79d6 and CYP79d7 catalyze the formation of volatile aldoximes involved in poplar defense. Plant Cell 2013, 25. [Google Scholar] [CrossRef] [PubMed]
- Irmisch, S.; McCormick, A.C.; Günther, J.; Schmidt, A.; Boeckler, G.A.; Gershenzon, J.; Unsicker, S.B.; Köllner, T.G. Herbivore-induced poplar cytochrome P450 enzymes of the CYP71 family convert aldoximes to nitriles which repel a generalist caterpillar. Plant J. 2014, 80, 1095–1107. [Google Scholar] [CrossRef] [PubMed]
- McCormick, A.C.; Irmisch, S.; Reinecke, A.; Boeckler, G.A.; Veit, D.; Reichelt, M.; Hansson, B.S.; Gershenzon, J.; Köllner, T.G.; Unsicker, S.B. Herbivore-induced volatile emission in black poplar: Regulation and role in attracting herbivore enemies. Plant Cell Environ. 2014, 37, 1909–1923. [Google Scholar] [CrossRef] [PubMed]
- Lackus, N.D.; Lackner, S.; Gershenzon, J.; Unsicker, S.B.; Köllner, T.G. The occurrence and formation of monoterpenes in herbivore-damaged poplar roots. Sci. Rep. 2018, 8, 17936. [Google Scholar] [CrossRef] [PubMed]
- Vranova, E.; Coman, D.; Gruissem, W. Structure and dynamics of the isoprenoid pathway network. Mol. Plant 2012, 5, 318–333. [Google Scholar] [CrossRef] [PubMed]
- Günther, J.; Lackus, N.D.; Schmidt, A.; Huber, M.; Stödler, H.-J.; Reichelt, M.; Gershenzon, J.; Köllner, T.G. Separate pathways contribute to the herbivore-induced formation of 2-phenylethanol in poplar. Plant Physiol. 2019, 180, 767–782. [Google Scholar] [CrossRef]
- Jia, Q.; Chen, F. Catalytic functions of the isoprenyl diphosphate synthase superfamily in plants: A growing repertoire. Mol. Plant 2016, 9, 189–191. [Google Scholar] [CrossRef]
- Cervantes-Cervantes, M.; Gallagher, C.E.; Zhu, C.; Wurtzel, E.T. Maize cDNAs expressed in endosperm encode functional farnesyl diphosphate synthase with geranylgeranyl diphosphate synthase activity. Plant Physiol. 2006, 141, 220–231. [Google Scholar] [CrossRef]
- Schmidt, A.; Gershenzon, J. Cloning and characterization of two different types of geranyl diphosphate synthases from norway spruce (Picea abies). Phytochemistry 2008, 69, 49–57. [Google Scholar] [CrossRef]
- Schmidt, A.; Wächtler, B.; Temp, U.; Krekling, T.; Séguin, A.; Gershenzon, J. A bifunctional geranyl and geranylgeranyl diphosphate synthase is involved in terpene oleoresin formation in Picea abies. Plant Physiol. 2010, 152, 639–655. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, Y.Y.; Jeng, M.F.; Tsai, W.C.; Chuang, Y.C.; Li, C.Y.; Wu, T.S.; Kuoh, C.S.; Chen, W.H.; Chen, H.H. A novel homodimeric geranyl diphosphate synthase from the orchid Phalaenopsis bellina lacking a DD(x)2-4D motif. Plant J. 2008, 55, 719–733. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.; Smita, S.S.; Singh, A.K.; Shanker, K.; Nagegowda, D.A. Heteromeric and homomeric geranyl diphosphate synthases from Catharanthus roseus and their role in monoterpene indole alkaloid biosynthesis. Mol. Plant 2013, 6, 1531–1549. [Google Scholar] [CrossRef] [PubMed]
- Cunillera, N.; Arró, M.; Delourme, D.; Karst, F.; Boronat, A.; Ferrer, A. Arabidopsis thaliana contains two differentially expressed farnesyl-diphosphate synthase genes. J. Biol. Chem. 1996, 271, 7774–7780. [Google Scholar] [CrossRef] [PubMed]
- Dhar, M.K.; Koul, A.; Kaul, S. Farnesyl pyrophosphate synthase: A key enzyme in isoprenoid biosynthetic pathway and potential molecular target for drug development. New Biotechnol. 2013, 30, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Richter, A.; Seidl-Adams, I.; Köllner, T.G.; Schaff, C.; Tumlinson, J.H.; Degenhardt, J. A small, differentially regulated family of farnesyl diphosphate synthases in maize (Zea mays) provides farnesyl diphosphate for the biosynthesis of herbivore-induced sesquiterpenes. Planta 2015, 241, 1351–1361. [Google Scholar] [CrossRef] [PubMed]
- Hemmerlin, A.; Rivera, S.B.; Erickson, H.K.; Poulter, C.D. Enzymes encoded by the farnesyl diphosphate synthase gene family in the big sagebrush Artemisia tridentata ssp. Spiciformis. J. Biol. Chem. 2003, 278, 32132–32140. [Google Scholar] [CrossRef]
- Wang, L.; Jing, T.; Li, T.; Du, H.; Wuyun, T.-N. Identification and expression analysis of the Eucommia ulmoides farnesyl diphosphate synthase gene family to reveal the key gene involved in rubber biosynthesis. Acta Physiol. Plant. 2017, 40, 11. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Z.X.; Yu, X.D.; Fan, J.; Pickett, J.A.; Jones, H.D.; Zhou, J.J.; Birkett, M.A.; Caulfield, J.; Napier, J.A.; et al. Molecular characterization of two isoforms of a farnesyl pyrophosphate synthase gene in wheat and their roles in sesquiterpene synthesis and inducible defence against aphid infestation. New Phytol. 2015, 206, 1101–1115. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. Mega6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
Data Availability: All data generated or analyzed during this study are included in the main text or supplement of this published article. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence | Usage |
|---|---|---|
| PtFPPS1-fwd | ATGGTAGGTCTCAGCGCATGGCAGATCTGAAGTCAACGTTC | cloning |
| PtFPPS1-rev | ATGGTAGGTCTCATATCATTTCTGCCTCTTGTAAATTTTAGCC | cloning |
| PtGPPS1.LSU-fwd | ATGGTAGGTCTCAAATGCCCACTTTTGATTTCAAGTCTTACA | cloning |
| PtGPPS1.LSU-rev | ATGGTAGGTCTCAGCGCTGTTTTGCCTGTAAGCAATGTAATTAG | cloning |
| PtGPPS2.LSU-fwd | ATGGTAACCTGCATTAAATGCCCACTTTTGATTTCAAGTCTTATAT | cloning |
| PtGPPS2.LSU-rev | ATGGTAACCTGCATTAGCGCTGTTTTGCCTGTAAGCAATGTAATTAG | cloning |
| PtGPPS.SSU1-fwd | ATGGTAGGTCTCAAATGGCAACTTCCAATGGCACTACTTAC | cloning |
| PtGPPS.SSU1-rev | ATGGTAGGTCTCAGCGCTAACATTGCCGGAAGTGGTCCCT | cloning |
| PtGPPS.SSU2-fwd | ATGGTAGGTCTCAAATGTCAAAAACACCCCAGTTTGATTTAAA | cloning |
| PtGPPS.SSU2-rev | ATGGTAGGTCTCAGCGCTACTTGACTCACCAAAACTGAAACC | cloning |
| PtFPPS1-fwd1 | GGTTGCCCAAGGTTGGTCTTATTGC | qRT-PCR |
| PtFPPS1-rev1 | TGAGTAGTAGGCGGTCTTGTACTG | qRT-PCR |
| PtGPPS1.LSU-fwd1 | TGGCGAAAGCTATTGG | qRT-PCR |
| PtGPPS1.LSU-rev1 | CGGTTCCTCCACCTAATATG | qRT-PCR |
| PtGPPS2.LSU-fwd1 | GTTGCTGGACAAGTTGTG | qRT-PCR |
| PtGPPS2.LSU-rev1 | TCCAATACTCCTCGCGTATC | qRT-PCR |
| PtGPPS.SSU1-fwd1 | TGCTCCAGCCTTGTGCATAG | qRT-PCR |
| PtGPPS.SSU1-rev1 | ACCATCCCATCTCCTGTTAG | qRT-PCR |
| PtGPPS.SSU2-fwd1 | GATTGCTAGCCGGTGCCAAG | qRT-PCR |
| PtGPPS.SSU2-rev1 | CCTCCGCTACCTCTATAGCC | qRT-PCR |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lackus, N.D.; Petersen, N.P.; Nagel, R.; Schmidt, A.; Irmisch, S.; Gershenzon, J.; Köllner, T.G. Identification and Characterization of trans-Isopentenyl Diphosphate Synthases Involved in Herbivory-Induced Volatile Terpene Formation in Populus trichocarpa. Molecules 2019, 24, 2408. https://doi.org/10.3390/molecules24132408
Lackus ND, Petersen NP, Nagel R, Schmidt A, Irmisch S, Gershenzon J, Köllner TG. Identification and Characterization of trans-Isopentenyl Diphosphate Synthases Involved in Herbivory-Induced Volatile Terpene Formation in Populus trichocarpa. Molecules. 2019; 24(13):2408. https://doi.org/10.3390/molecules24132408
Chicago/Turabian StyleLackus, Nathalie D., Nora P. Petersen, Raimund Nagel, Axel Schmidt, Sandra Irmisch, Jonathan Gershenzon, and Tobias G. Köllner. 2019. "Identification and Characterization of trans-Isopentenyl Diphosphate Synthases Involved in Herbivory-Induced Volatile Terpene Formation in Populus trichocarpa" Molecules 24, no. 13: 2408. https://doi.org/10.3390/molecules24132408
APA StyleLackus, N. D., Petersen, N. P., Nagel, R., Schmidt, A., Irmisch, S., Gershenzon, J., & Köllner, T. G. (2019). Identification and Characterization of trans-Isopentenyl Diphosphate Synthases Involved in Herbivory-Induced Volatile Terpene Formation in Populus trichocarpa. Molecules, 24(13), 2408. https://doi.org/10.3390/molecules24132408