Emissive Enhancement of the Singlet Oxygen Chemiluminescence Probe after Binding to Bovine Serum Albumin

 , and
, and 
Abstract
:
1. Introduction
2. Results
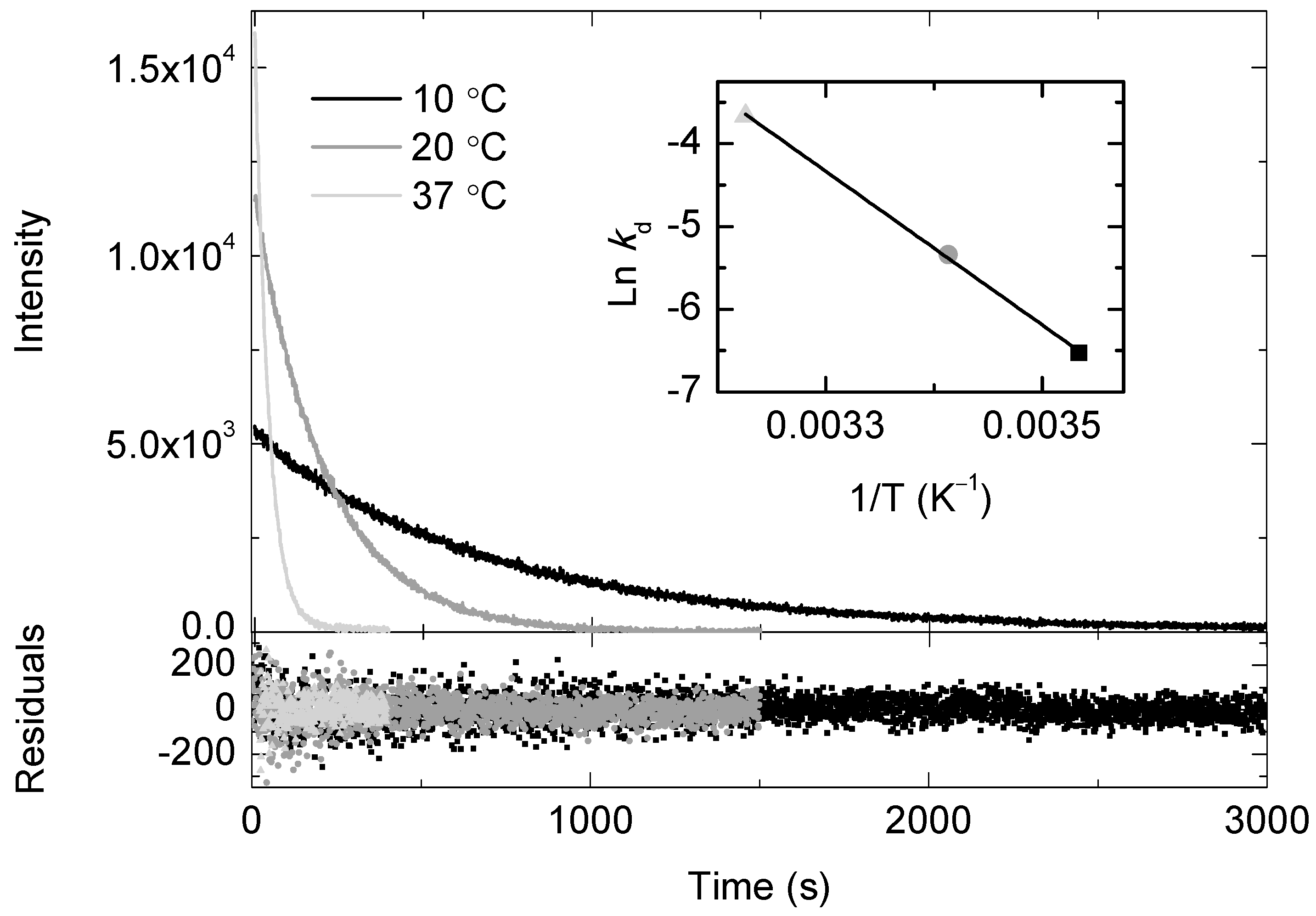
2.1. Temperature and Molecular Oxygen Effects on the Chemiluminescence Signal of the SOCL Phenoxy-Dioxetane
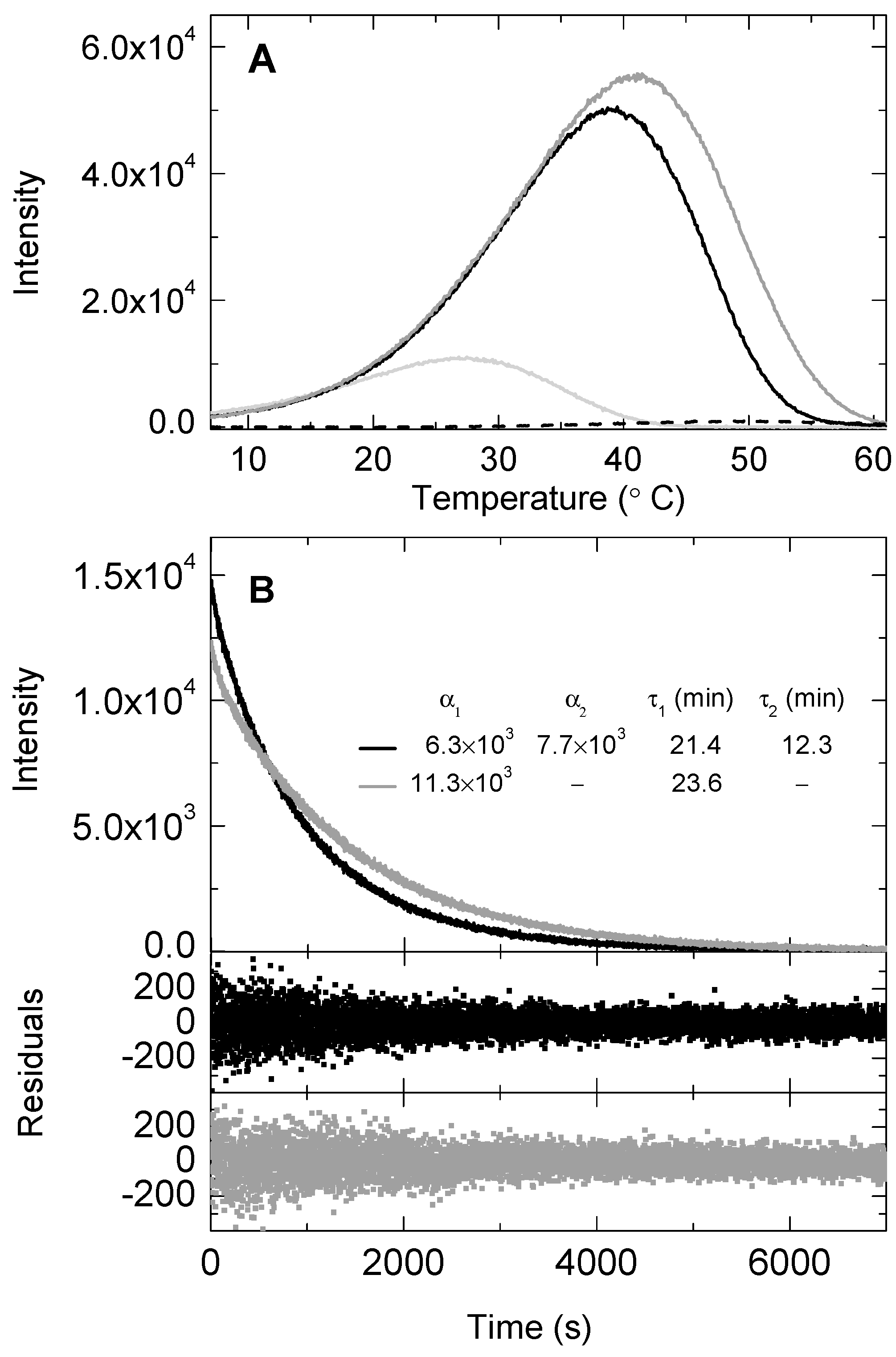
2.2. BSA Effect on the Chemiluminescence Signal of the SOCL Phenoxy-Dioxetane
2.3. SOCL and BSA Binding Energetics
3. Discussion
4. Material and Methods
4.1. Chemical Reagents and Stock Solutions
4.2. Oxygen Consumption Experiments
4.3. Chemiluminescence Experiments
4.4. ITC Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Schweitzer, C.; Schmidt, R. Physical mechanisms of generation and deactivation of singlet oxygen. Chem. Rev. 2003, 103, 1685–1757. [Google Scholar] [CrossRef] [PubMed]
- Losev, A.P.; Byteva, I.M.; Gurinovich, G.P. Singlet oxygen luminescence quantum yields in organic solvents and water. Chem. Phys. Lett. 1988, 143, 127–129. [Google Scholar] [CrossRef]
- Li, H.; Melø, T.B.; Arellano, J.B.; Naqvi, K.R. Temporal profile of the singlet oxygen emission endogenously produced by photosystem II reaction centre in an aqueous buffer. Photosynth. Res. 2012, 112, 75–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorman, A.A.; Rodgers, M.A.J. Current perspectives of singlet oxygen detection in biological environments. J. Photochem. Photobiol. B Biol. 1992, 14, 159–176. [Google Scholar] [CrossRef]
- Bisby, R.H.; Morgan, C.G.; Hamblett, I.; Gorman, A.A. Quenching of singlet oxygen by Trolox C, ascorbate, and amino acids: Effects of pH and temperature. J. Phys. Chem. A 1999, 103, 7454–7459. [Google Scholar] [CrossRef]
- Da Silva, E.F.F.; Pedersen, B.W.; Breitenbach, T.; Toftegaard, R.; Kuimova, M.K.; Arnaut, L.G.; Ogilby, P.R. Irradiation- and sensitizer-dependent changes in the lifetime of intracellular singlet oxygen produced in a photosensitized process. J. Phys. Chem. B 2012, 116, 445–461. [Google Scholar] [CrossRef] [PubMed]
- Maisch, T.; Baier, J.; Franz, B.; Maier, M.; Landthaler, M.; Szeimies, R.-M.; Baeumler, W. The role of singlet oxygen and oxygen concentration in photodynamic inactivation of bacteria. Proc. Natl. Acad. Sci. USA 2007, 104, 7223–7228. [Google Scholar] [CrossRef] [Green Version]
- Hideg, E. A comparative study of fluorescent singlet oxygen probes in plant leaves. Cent. Eur. J. Biol. 2008, 3, 273–284. [Google Scholar] [CrossRef]
- Ruiz-González, R.; Zanocco, A.L. Singlet oxygen fluorescent probes. In Singlet Oxygen: Applications in Biosciences and Nanosciences; Nonell, S., Flors, C., Eds.; The Royal Society of Chemistry: Cambridge, UK, 2016; Volume 2, pp. 103–120. [Google Scholar] [CrossRef]
- Chen, X.Q.; Wang, F.; Hyun, J.Y.; Wei, T.W.; Qiang, J.; Ren, X.T.; Shin, I.; Yoon, J. Recent progress in the development of fluorescent, luminescent and colorimetric probes for detection of reactive oxygen and nitrogen species. Chem. Soc. Rev. 2016, 45, 2976–3016. [Google Scholar] [CrossRef]
- Nardi, G.; Manet, I.; Monti, S.; Miranda, M.A.; Lhiaubet-Vallet, V. Scope and limitations of the TEMPO/EPR method for singlet oxygen detection: The misleading role of electron transfer. Free Radic. Biol. Med. 2014, 77, 64–70. [Google Scholar] [CrossRef]
- Tanaka, K.; Miura, T.; Umezawa, N.; Urano, Y.; Kikuchi, K.; Higuchi, T.; Nagano, T. Rational design of fluorescein-based fluorescence probes, mechanism-based design of a maximum fluorescence probe for singlet oxygen. J. Am. Chem. Soc. 2001, 123, 2530–2536. [Google Scholar] [CrossRef] [PubMed]
- Nagano, T. Bioimaging probes for reactive oxygen species and reactive nitrogen species. J. Clin. Biochem. Nutr. 2009, 45, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Hananya, N.; Green, O.; Blau, R.; Satchi-Fainaro, R.; Shabat, D. A highly efficient chemiluminescence probe for the detection of singlet oxygen in living cells. Angew. Chem. Int. Ed. Engl. 2017, 56, 11793–11796. [Google Scholar] [CrossRef] [PubMed]
- Schaap, A.P.; Gagnon, S.D. Chemiluminescence from a phenoxide-substituted 1,2-dioxetane: A model for firefly bioluminescence. J. Am. Chem. Soc. 1982, 104, 3504–3506. [Google Scholar] [CrossRef]
- Gnaim, S.; Green, O.; Shabat, D. The emergence of aqueous chemiluminescence: New promising class of phenoxy 1,2-dioxetane luminophores. Chem. Commun. 2018, 54, 2073–2085. [Google Scholar] [CrossRef] [PubMed]
- Green, O.; Gnaim, S.; Blau, R.; Eldar-Boock, A.; Satchi-Fainaro, R.; Shabat, D. Near-infrared dioxetane luminophores with direct chemiluminescence emission mode. J. Am. Chem. Soc. 2017, 139, 13243–13248. [Google Scholar] [CrossRef] [PubMed]
- Bruemmer, K.J.; Green, O.; Su, T.A.; Shabat, D.; Chang, C.J. Chemiluminescent probes for activity-based sensing of formaldehyde released from folate degradation in living mice. Angew. Chem. Int. Ed. Engl. 2018, 57, 7508–7512. [Google Scholar] [CrossRef]
- Hananya, N.; Reid, J.P.; Green, O.; Sigman, M.S.; Shabat, D. Rapid chemiexcitation of phenoxy-dioxetane luminophores yields ultrasensitive chemiluminescence assays. Chem. Sci. 2019, 10, 1380–1385. [Google Scholar] [CrossRef]
- Roth-Konforti, M.E.; Bauer, C.R.; Shabat, D. Unprecedented sensitivity in a probe for monitoring cathepsin B: Chemiluminescence microscopy cell-imaging of a natively expressed enzyme. Angew. Chem. Int. Ed. Engl. 2017, 56, 15633–15638. [Google Scholar] [CrossRef]
- Hananya, N.; Shabat, D. Recent advances and challenges in luminescent imaging: Bright outlook for chemiluminescence of dioxetanes in water. ACS Cent. Sci. 2019, 5, 949–959. [Google Scholar] [CrossRef]
- Gollmer, A.; Arnbjerg, J.; Blaikie, F.H.; Pedersen, B.W.; Breitenbach, T.; Daasbjerg, K.; Glasius, M.; Ogilby, P.R. Singlet Oxygen Sensor Green®: Photochemical behavior in solution and in a mammalian cell. Photochem. Photobiol. 2011, 87, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-González, R.; Bresolí-Obach, R.; Gulías, Ò.; Agut, M.; Savoie, H.; Boyle, R.W.; Nonell, S.; Giuntini, F. NanoSOSG: A nanostructured fluorescent probe for the detection of intracellular singlet oxygen. Angew. Chem. Int. Ed. Engl. 2017, 56, 2885–2888. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J. Singlet oxygen-mediated damage to proteins and its consequences. Biochem. Biophys. Res. Commun. 2003, 305, 761–770. [Google Scholar] [CrossRef]
- Jensen, R.L.; Arnbjerg, J.; Birkedal, H.; Ogilby, P.R. Singlet oxygen’s response to protein dynamics. J. Am. Chem. Soc. 2011, 133, 7166–7173. [Google Scholar] [CrossRef] [PubMed]
- Leitner, D.M.; Gruebele, M.; Havenith, M. Solvation dynamics of biomolecules: Modeling and terahertz experiments. HFSP J. 2008, 2, 314–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, W.; Jiang, G.; Zhou, Q.; Zhang, B.; Wang, X. Greatly enhanced binding of a cationic porphyrin towards bovine serum albumin by cucurbit [8] uril. Phys. Chem. Chem. Phys. 2010, 12, 13255–13260. [Google Scholar] [CrossRef] [PubMed]
- Alarcón, E.; Maria Edwards, A.; Aspee, A.; Moran, F.E.; Borsarelli, C.D.; Lissi, E.A.; Gonzalez-Nilo, D.; Poblete, H.; Scaiano, J.C. Photophysics and photochemistry of dyes bound to human serum albumin are determined by the dye localization. Photochem. Photobiol. Sci. 2010, 9, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Baier, J.; Maisch, T.; Regensburger, J.; Loibl, M.; Vasold, R.; Baeumler, W. Time dependence of singlet oxygen luminescence provides an indication of oxygen concentration during oxygen consumption. J. Biomed. Opt. 2007, 12. [Google Scholar] [CrossRef] [PubMed]
- Naveenraj, S.; Anandan, S. Binding of serum albumins with bioactive substances—Nanoparticles to drugs. J. Photochem. Photobiol. C Photochem. Rev. 2013, 14, 53–71. [Google Scholar] [CrossRef]
- Arellano, J.B.; Yousef, Y.A.; Melø, T.B.; Mohamad, S.B.B.; Cogdell, R.J.; Naqvi, K.R. Formation and geminate quenching of singlet oxygen in purple bacterial reaction center. J. Photochem. Photobiol. B Biol. 2007, 87, 105–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appiani, E.; Ossola, R.; Latch, D.E.; Erickson, P.R.; McNeill, K. Aqueous singlet oxygen reaction kinetics of furfuryl alcohol: Effect of temperature, pH, and salt content. Environ. Sci. Process. Impacts 2017, 19, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Scully, F.E.; Hoigne, J. Rate constants for reactions of singlet oxygen with phenols and other compounds in water. Chemosphere 1987, 16, 681–694. [Google Scholar] [CrossRef]
- Yabuta, Y.; Tamoi, M.; Yamamoto, K.; Tomizawa, K.-i.; Yokota, A.; Shigeoka, S. Molecular design of photosynthesis-elevated chloroplasts for mass accumulation of a foreign protein. Plant. Cell Physiol. 2008, 49, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Arellano, J.B.; Naqvi, K.R. Endogenous singlet oxygen photosensitizers in plants. In Singlet Oxygen: Applications in Biosciences and Nanosciences; Nonell, S., Flors, C., Eds.; The Royal Society of Chemistry: Cambridge, UK, 2016; Volume 1, pp. 239–269. [Google Scholar] [CrossRef]
- Hu, Y.J.; Liu, Y.; Zhao, R.M.; Dong, J.X.; Qu, S.S. Spectroscopic studies on the interaction between methylene blue and bovine serum albumin. J. Photochem. Photobiol. A Chem. 2006, 179, 324–329. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Y.; Sun, S.; Zhang, A.; Liu, Y. Binding investigation on the interaction between Methylene Blue (MB)/TiO2 nanocomposites and bovine serum albumin by resonance light-scattering (RLS) technique and fluorescence spectroscopy. J. Photochem. Photobiol. B Biol. 2013, 128, 12–19. [Google Scholar] [CrossRef] [PubMed]
- He, L.-L.; Wang, Y.-X.; Wu, X.-X.; Liu, X.-P.; Wang, X.; Liu, B.; Wang, X. Enhancement of the binding affinity of methylene blue to site I in human serum albumin by cupric and ferric ions. Luminescence 2015, 30, 1380–1388. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.J.; Li, W.; Liu, Y.; Dong, J.X.; Qu, S.S. Fluorometric investigation of the interaction between methylene blue and human serum albumin. J. Pharm. Biomed. Anal. 2005, 39, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Jensen, R.L.; Arnbjerg, J.; Ogilby, P.R. Reaction of singlet oxygen with tryptophan in proteins: A pronounced effect of the local environment on the reaction rate. J. Am. Chem. Soc. 2012, 134, 9820–9826. [Google Scholar] [CrossRef] [PubMed]
- Mazur, S.; Foote, C.S. Chemistry of singlet oxygen. IX. A stable dioxetane from photooxygenation of tetramethoxyethylene. J. Am. Chem. Soc. 1970, 92, 3225–3226. [Google Scholar] [CrossRef]
- MacManus-Spencer, L.A.; Latch, D.E.; Kroncke, K.M.; McNeill, K. Stable dioxetane precursors as selective trap-and-trigger chemiluminescent probes for singlet oxygen. Anal. Chem. 2005, 77, 1200–1205. [Google Scholar] [CrossRef] [PubMed]
- Adam, W.; Matsumoto, M.; Trofimov, A.V. Viscosity dependence of the chemically induced electron-exchange chemiluminescence triggered from a bicyclic dioxetane. J. Am. Chem. Soc. 2000, 122, 8631–8634. [Google Scholar] [CrossRef]
- Adam, W.; Bronstein, I.; Trofimov, A.V.; Vasil’ev, R.F. Solvent-cage effect (viscosity dependence) as a diagnostic probe for the mechanism of the intramolecular chemically initiated electron-exchange luminescence (CIEEL) triggered from a spiroadamantyl-substituted dioxetane. J. Am. Chem. Soc. 1999, 121, 958–961. [Google Scholar] [CrossRef]
- Schaap, A.P.; Handley, R.S.; Giri, B.P. Chemical and enzymatic triggering of 1,2-dioxetanes. 1: Aryl esterase-catalyzed chemiluminescence from a naphthyl acetate-substituted dioxetane. Tetrahedron Lett. 1987, 28, 935–938. [Google Scholar] [CrossRef]
- Schaap, A.P.; Chen, T.S.; Handley, R.S.; Desilva, R.; Giri, B.P. Chemical and enzymatic triggering of 1,2-dioxetanes 2: Fluoride-induced chemiluminescence from tert-butyldimethylsilyloxy-substituted dioxetanes. Tetrahedron Lett. 1987, 28, 1155–1158. [Google Scholar] [CrossRef]
- Geddes, C.D. Optical halide sensing using fluorescence quenching: Theory, simulations and applications—A review. Meas. Sci. Technol. 2001, 12, R53–R88. [Google Scholar] [CrossRef]
- Velazquez-Campoy, A.; Todd, M.J.; Freire, E. HIV-1 protease inhibitors: Enthalpic versus entropic optimization of the binding affinity. Biochemistry 2000, 39, 2201–2207. [Google Scholar] [CrossRef] [PubMed]
- Jelesarov, I.; Bosshard, H.R. Isothermal titration calorimetry and differential scanning calorimetry as complementary tools to investigate the energetics of biomolecular recognition. J. Mol. Recognit. 1999, 12, 3–18. [Google Scholar] [CrossRef]
- Claveria-Gimeno, R.; Vega, S.; Abian, O.; Velazquez-Campoy, A. A look at ligand binding thermodynamics in drug discovery. Expert Opin. Drug Discov. 2017, 12, 363–377. [Google Scholar] [CrossRef]
- Bastos, E.L.; Da Silva, S.M.; Baader, W.J. Solvent cage effects: Basis of a general mechanism for efficient chemiluminescence. J. Org. Chem. 2013, 78, 4432–4439. [Google Scholar] [CrossRef]
- Signorelli, S.; Arellano, J.B.; Melø, T.B.; Borsani, O.; Monza, J. Proline does not quench singlet oxygen: Evidence to reconsider its protective role in plants. Plant. Physiol. Biochem. 2013, 64, 80–83. [Google Scholar] [CrossRef] [Green Version]
- Buey, R.M.; Arellano, J.B.; López-Maury, L.; Galindo-Trigo, S.; Velázquez-Campoy, A.; Revuelta, J.L.; De Pereda, J.M.; Florencio, F.J.; Schürmann, P.; Buchanan, B.B.; et al. Unprecedented pathway of reducing equivalents in a diflavin-linked disulfide oxidoreductase. Proc. Natl. Acad. Sci. USA 2017, 114, 12725–12730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Sample Availability: Samples of the compound Singlet Oxygen Chemiluminescence probe (SOCL) is available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ka (M−1) | Kd (μM) | ΔH (kJ mol−1) | n |
|---|---|---|---|
| (5.9 ± 0.9) × 105 | 1.7 ± 0.3 | 7.9 ± 1.2 | 2.1 ± 0.1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miranda-Apodaca, J.; Hananya, N.; Velázquez-Campoy, A.; Shabat, D.; Arellano, J.B. Emissive Enhancement of the Singlet Oxygen Chemiluminescence Probe after Binding to Bovine Serum Albumin. Molecules 2019, 24, 2422. https://doi.org/10.3390/molecules24132422
Miranda-Apodaca J, Hananya N, Velázquez-Campoy A, Shabat D, Arellano JB. Emissive Enhancement of the Singlet Oxygen Chemiluminescence Probe after Binding to Bovine Serum Albumin. Molecules. 2019; 24(13):2422. https://doi.org/10.3390/molecules24132422
Chicago/Turabian StyleMiranda-Apodaca, Jon, Nir Hananya, Adrián Velázquez-Campoy, Doron Shabat, and Juan B. Arellano. 2019. "Emissive Enhancement of the Singlet Oxygen Chemiluminescence Probe after Binding to Bovine Serum Albumin" Molecules 24, no. 13: 2422. https://doi.org/10.3390/molecules24132422
APA StyleMiranda-Apodaca, J., Hananya, N., Velázquez-Campoy, A., Shabat, D., & Arellano, J. B. (2019). Emissive Enhancement of the Singlet Oxygen Chemiluminescence Probe after Binding to Bovine Serum Albumin. Molecules, 24(13), 2422. https://doi.org/10.3390/molecules24132422