Biotechnological Advances in Resveratrol Production and its Chemical Diversity



Abstract
:1. Introduction
2. Biological Significance in Humans
2.1. Biosynthesis of Resveratrol in Plants
2.2. Alternative Sources of Resveratrol
2.3. Microbial Biosynthesis of Resveratrol
2.4. Biosynthesis of Resveratrol in Non-E. coli Hosts
2.5. Biosynthesis of Resveratrol in E. coli Host
2.6. Pathway Engineering
2.7. Steps for Increasing the Precursor Pool
2.8. Protein Engineering
2.9. De novo Pathway Engineering
2.10. Central Carbon-Flux Redirection
2.11. Optimized Conditions for Higher Resveratrol Production
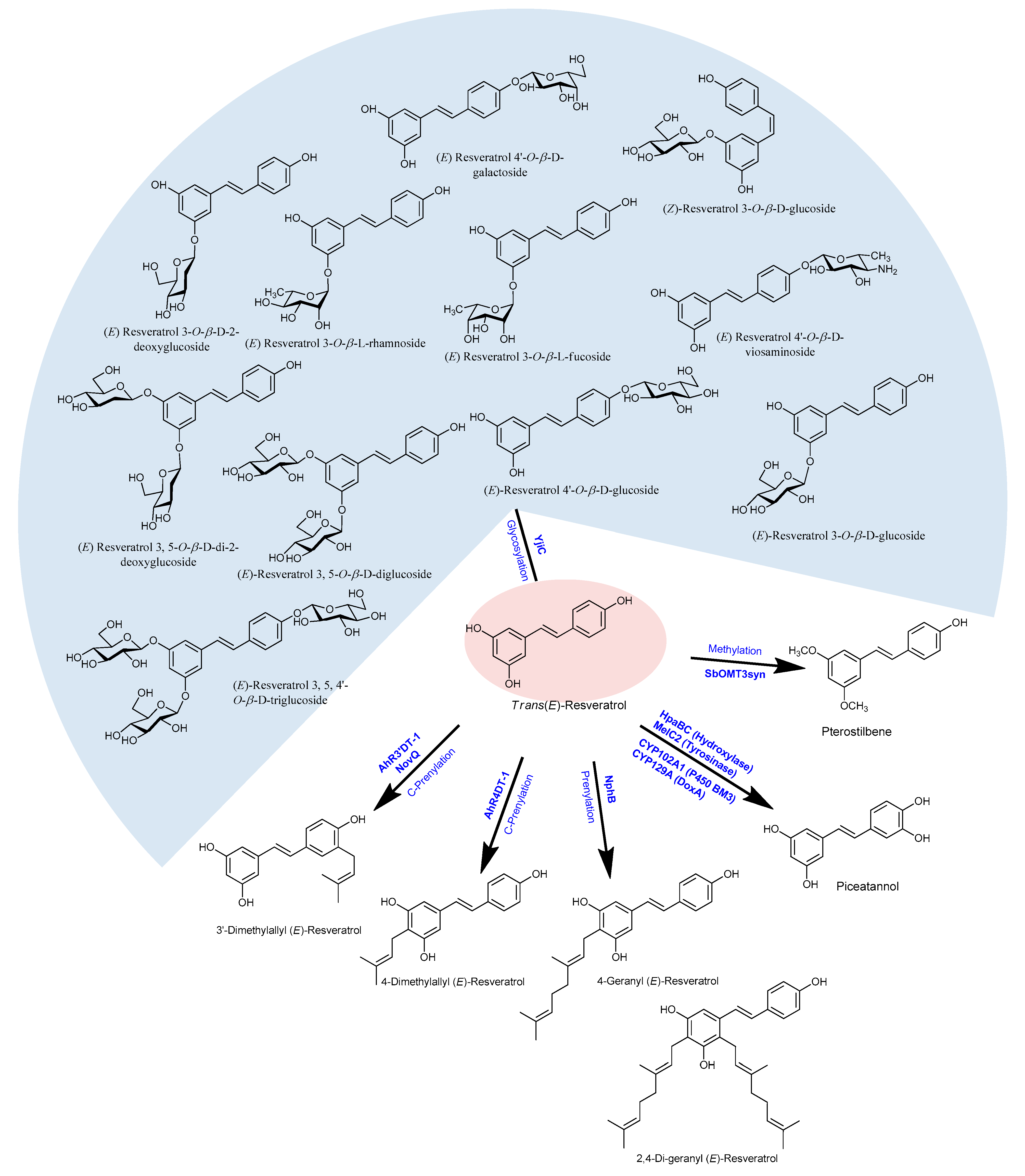
2.12. Microbial Production of Resveratrol Derivatives
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Pandey, R.P.; Parajuli, P.; Shin, J.Y.; Lee, J.; Lee, S.; Hong, Y.S. Enzymatic biosynthesis of novel resveratrol glucoside and glycoside derivatives. Appl. Environ. Microbiol. 2014, 80, 7235–7243. [Google Scholar] [CrossRef] [PubMed]
- Gambini, J.; Inglés, M.; Olaso, G.; Lopez-Grueso, R.; Bonet-Costa, V.; Gimeno-Mallench, L.; Borras, C. Properties of resveratrol: In vitro and in vivo studies about metabolism, bioavailability, and biological effects in animal models and humans. Oxid. Med. Cell. Longev. 2015. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Mar, M.I.; Mateos, R.; García-Parrilla, M.C.; Puertas, B.; Cantos-Villar, E. Bioactive compounds in wine: Resveratrol, hydroxytyrosol and melatonin: A review. Food Chem. 2012, 130, 797–813. [Google Scholar] [CrossRef]
- Mikulski, D.; Go´rniak, R.; Molski, M. A theoretical study of the structure-radical scavenging activity of trans-resveratrol analogues and cis-resveratrol in gas phase and water environment. Eur. J. Med. Chem. 2010, 45, 1015–1027. [Google Scholar] [CrossRef] [PubMed]
- Savoia, D. Plant-derived antimicrobial compounds: Alternatives to antibiotics. Future Microbiol. 2012, 7, 979–990. [Google Scholar] [CrossRef] [PubMed]
- Camont, L.; Cottart, C.H.; Rhayem, Y.; Nivet-Antoine, V.; Djelidi, R.; Collin, F.; Beaudeux, J.L.; Bonnefont-Rousselot, D. Simple spectrophotometric assessment of the trans-/cis-resveratrol ratio in aqueous solutions. Anal. Chim. Acta 2009, 634, 121–128. [Google Scholar] [CrossRef]
- Burns, J.; Yokota, T.; Ashihara, H.; Lean, M.E.; Crozier, A. Plant foods and herbal sources of resveratrol. J. Agric. Food Chem. 2002, 50, 3337–3340. [Google Scholar] [CrossRef] [PubMed]
- Roldán, A.; Palacios, V.; Caro, I.; Pérez, L. Resveratrol content of Palomino fino grapes: Influence of vintage and fungal infection. J. Agric. Food Chem. 2003, 51, 1464–1468. [Google Scholar] [CrossRef]
- Chen, X.; He, H.; Wang, G.; Yang, B.; Ren, W.; Ma, L.; Yu, Q. Stereospecific determination of cis- and trans-resveratrol in rat plasma by HPLC: Application to pharmacokinetic studies. Biomed. Chromatogr. 2007, 21, 257–265. [Google Scholar] [CrossRef]
- Takaoka, M. Resveratrol, a new phenolic compound, from Veratrum grandiflorum. J. Chem. Soc. Japan. 1939, 60, 1090–1100. [Google Scholar]
- Bauer, J.H.; Goupil, S.; Garber, G.B.; Helfand, S.L. An accelerated assay for the identification of lifespan-extending interventions in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2004, 101, 12980–12985. [Google Scholar] [CrossRef] [PubMed]
- Navindra, P.S.; Vishal, V.K.; Subhash, P. Sources and chemistry of resveratrol. In Resveratrol in Health and Disease, 1st ed.; Bharat, B.A., Shishir, S., Eds.; CRC Press: Boca Raton, FL, USA, 2005; pp. 17–32. [Google Scholar]
- Langcake, P.; Pryce, R.J. A new class of phytoalexins from grapevines. Experientia 1977, 33, 151–152. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lin, Y.; Li, L.; Linhardt, R.J.; Yan, Y. Regulating malonyl-CoA metabolism via synthetic antisense RNAs for enhanced biosynthesis of natural products. Metab. Eng. 2015, 29, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Fu, Z.D.; Wang, F.; Liu, H.Y.; Han, R. Anti-angiogenic activity of resveratrol, a natural compound from medicinal plants. J. Asian Nat. Prod. Res. 2005, 7, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Bru, R.; Selles, S.; Casado-Vela, J. Modified cyclodextrins are chemically defined glucan inducers of defense responses in grapevine cell cultures. J. Agric. Food Chem. 2006, 54, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Baur, J.A.; Pearson, K.J.; Price, N.L.; Jamieson, H.A.; Lerin, C.; Kalra, A.; Prabhu, V.V.; Lewis, K.; Poosala, S.; Pistell, P.J. Resveratrol improves health and survival of mice on a high-calorie diet. Nature 2006, 444, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Bavaresco, L.; Petegolli, D.; Canto, E.; Fregoni, M.; Chiusa, G.; Trevisan, M. Elicitation and accumulation of stilbene phytoalexins in grapevine berries infected by Botrytis cinerea. Vitis 1997, 36, 77–84. [Google Scholar]
- Dixon, R.A. Natural products and plant disease resistance. Nature 2001, 411, 843–847. [Google Scholar] [CrossRef]
- Donnez, D.; Jeandet, P.; Clement, C. Bioproduction of resveratrol and stilbene derivatives by plant cells and microorganisms. Trends Biotechnol. 2009, 27, 706–713. [Google Scholar] [CrossRef]
- Nonomura, S.; Kanagawa, H.; Makimoto, A. Chemical constituents of polygonaceous plants. i. studies on the components of ko-j o-kon.(polygonum cuspidatum sieb. et zucc.). Yakugaku zasshi: J. Pharm. Soc. Japan 1963, 83, 988–990. [Google Scholar] [CrossRef]
- Laza-Knoerr, A.L.; Gref, R.; Couvreur, P. Cyclodextrins for drug delivery. J. Drug Target 2010, 18, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Knutson, M.D.; Leeuwenburgh, C. Resveratrol and novel potent activators of SIRT1: Effects on aging and age-related diseases. Nutr. Rev. 2008, 66, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Ahmad, F.; Philp, A.; Baar, K.; Williams, T.; Luo, H.; Ke, H.; Rehmann, H.; Taussig, R.; Brown, A.L.; et al. Resveratrol ameliorates aging-related metabolic phenotypes by inhibiting cAMP phosphodiesterases. Cell 2012, 148, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Pangeni, R.; Sahni, J.K.; Ali, J.; Sharma, S.; Baboota, S. Resveratrol: Review on therapeutic potential and recent advances in drug delivery. Expert. Opin. Drug. Deliv. 2014, 11, 1285–1298. [Google Scholar] [CrossRef] [PubMed]
- Okawara, M.; Katsuki, H.; Kurimoto, E.; Shibata, H.; Kume, T.; Akaike, A. Resveratrol protects dopaminergic neurons in midbrain slice culture from multiple insults. Biochem. Pharm. 2007, 73, 550–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenz, M.; Hauser, A.C.; Püspök-Schwarz, M.; Kotanko, P.; Arias, I.; Zodl, H.; Sunder-Plassmann, G. Anderson-Fabry disease in Austria. Wien Klin Wochenschr 2003, 115, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Bradamante, S.; Barenghi, L.; Villa, A. Cardiovascular protective effects of resveratrol. Cardiovasc. Drug Rev. 2004, 22, 169–188. [Google Scholar] [CrossRef]
- Berrougui, H.; Grenier, G.; Loued, S.; Drouin, G.; Khalil, A. A new insight into resveratrol as an atheroprotective compound: Inhibition of lipid peroxidation and enhancement of cholesterol efflux. Atherosclerosis 2009, 207, 420–427. [Google Scholar] [CrossRef]
- Jang, M.; Cai, L.; Udeani, G.O.; Slowing, K.V.; Thomas, C.F.; Beecher, C.W.; Fong, H.H.; Farnsworth, N.R.; Kinghorn, A.D.; Mehta, R.G.; et al. Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. Science 1997, 275, 218–220. [Google Scholar] [CrossRef]
- Bai, X.; Yu, W.; Ji, W.; Lin, Z.; Tan, S.; Duan, K.; Li, N. Early versus delayed administration of norepinephrine in patients with septic shock. Crit. Care 2014. [Google Scholar] [CrossRef]
- Alarcon De La Lastra, C.; Villegas, I. Resveratrol as an anti-inflammatory and anti-aging agent: Mechanisms and clinical implications. Mol. Nutr. Food Res. 2005, 49, 405–430. [Google Scholar] [CrossRef]
- Holthoff, J.H.; Woodling, K.A.; Doerge, D.R. Resveratrol, a dietary polyphenolic phytoalexin, is a functional scavenger of peroxynitrite. Biochem. Pharmacol. 2010, 80, 1260–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lancon, A.; Kaminski, J.; Tili, E. Control of microRNA expression as a new way for resveratrol to deliver its beneficial effects. J. Agric. Food Chem. 2012, 60, 8783–8789. [Google Scholar] [CrossRef]
- Stervbo, U.; Vang, O.; Bonnesen, C. A review of the content of the putative chemopreventive phytoalexin resveratrol in red wine. Food Chem. 2007, 101, 449–457. [Google Scholar] [CrossRef]
- Whitlock, N.C.; Baek, S.J. The anticancer effects of resveratrol: Modulation of transcription factors. Nutr. Cancer 2012, 64, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Szkudelski, T. Resveratrol inhibits insulin secretion from rat pancreatic islets. Eur. J. Pharmacol. 2006, 552, 176–181. [Google Scholar] [CrossRef]
- Sun, C.; Zhang, F.; Ge, X.; Yan, T.; Chen, X.; Shi, X.; Zhai, Q. SIRT1improves insulin sensitivity under insulin-resistant conditions by repressing PTP1B. Cell Metab. 2007, 6, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Petrovski, G.; Gurusamy, N.; Das, D.K. Resveratrol in cardiovascular health and disease. Ann. NY Acad. Sci. 2011, 1215, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Pezzuto, J.M. Grapes and human health: A perspective. J. Agric. Food Chem. 2008, 56, 6777–6784. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.R.; Lokhandwala, M.F.; Banday, A.A. Resveratrol prevents endothelial nitric oxide synthase uncoupling and attenuates development of hypertension in spontaneously hypertensive rats. Eur. J. Pharmacol. 2011, 667, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Jeandet, P.; Sobarzo-Sanchez, E.; Clement, C.; Nabavi, S.F.; Habtemariam, S.; Nabavi, S.M.; Cordelier, S. Engineering stilbene metabolic pathways in microbial cells. Biotechnol. Adv. 2018, 36, 2264–2283. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, A.; Pandey, R.P.; Sohng, J.K. Biosynthesis of resveratrol and piceatannol in engineered microbial strains: Achievements and perspectives. Appl. Microbiol. Biotechnol. 2019, 103, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Braga, A.; Ferreira, P.; Oliveira, J.; Rocha, I.; Faria, N. Heterologous production of resveratrol in bacterial hosts: Current status and perspectives. World J. Microbiol. Biotechnol. 2018. [Google Scholar] [CrossRef]
- Light, S.H.; Halavaty, A.S.; Minasov, G. Structural analysis of a 3-deoxy-D-arabino-heptulosonate 7-phosphate synthase with an N-terminal chorismate mutase-like regulatory domain. Protein Sci. 2012, 21, 887–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, H.; Dudareva, N. The Shikimate pathway and aromatic amino acid biosynthesis in plants. Annu. Rev. Plant Biol. 2012, 63, 73–105. [Google Scholar] [CrossRef]
- Rosler, J.; Krekel, F.; Amrhein, N. Maize phenylalanine ammonia-lyase has tyrosine ammonia-lyase activity. Plant Physiol. 1997, 113, 175–179. [Google Scholar] [CrossRef]
- Ferrer, J.L.; Austin, M.B.; Stewart, C., Jr. Structure and function of enzymes involved in the biosynthesis of phenylpropanoids. Plant Physiol. Biochem. 2008, 46, 356–370. [Google Scholar] [CrossRef] [Green Version]
- Camacho-Zaragoza, J.M.; Hernández-Chávez, G.; Moreno-Avitia, F.; Ramírez-Iñiguez, R.; Martínez, A.; Bolívar, F.; Gosset, G. Engineering of a microbial coculture of Escherichia coli strains for the biosynthesis of resveratrol. Microb. Cell Fact. 2016. [Google Scholar] [CrossRef]
- Wang, W.; Wan, S.B.; Zhang, P.; Wang, H.L.; Zhan, J.C.; Huang, W.D. Prokaryotic expression, polyclonal antibody preparation of the stilbene synthase gene from grape berry and its different expression in fruit development and under heat acclimation. Plant Physiol. Biochem. 2008, 46, 1085–1092. [Google Scholar] [CrossRef]
- Guo, Y.W.; Guo, H.L.; Li, X.; Huang, L.L.; Zhang, B.N.; Pang, X.B.; Wang, H. Two type III polyketide synthases from Polygonum cuspidatum: Gene structure, evolutionary route and metabolites. Plant Biotechnol. Rep. 2013, 7, 371–381. [Google Scholar] [CrossRef]
- Raiber, S.; Schröder, G.; Schröder, J. Molecular and enzymatic characterization of two stilbene synthases from Eastern white pine (Pinus strobus) A single Arg/His difference determines the activity and the pH dependence of the enzymes. FEBS Lett. 1995, 361, 299–302. [Google Scholar] [CrossRef]
- Kodan, A.; Kuroda, H.; Sakai, F. A stilbene synthase from Japanese red pine (Pinus densiflora): Implications for phytoalexin accumulation and down-regulation of flavonoid biosynthesis. Proc. Natl. Acad. Sci. USA 2002, 99, 3335–3339. [Google Scholar] [CrossRef] [PubMed]
- Condori, J.; Medrano, G.; Sivakumar, G.; Nair, V.; Cramer, C.; Medina-Bolivar, F. Functional characterization of a stilbene synthase gene using a transient expression system in planta. Plant Cell Rep. 2009, 28, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Liu, S.J.; Ma, S.S.; Yang, W.; Hu, Y.L.; Wu, Q.; Lin, Z.P. Effect of Matrix Attachment Regions on Resveratrol Production in Tobacco with Transgene of Stilbene Synthase from Parthenocissus henryan. Acta Bot. Sin. 2004, 46, 948–954. [Google Scholar]
- Wang, C.; Zhi, S.; Liu, C.; Xu, F.; Zhao, A.; Wang, X.; Yu, M. Characterization of stilbene synthase genes in mulberry (Morus atropurpurea) and metabolic engineering for the production of resveratrol in Escherichia coli. J. Agri. Food Chem. 2017, 65, 1659–1668. [Google Scholar] [CrossRef]
- Samappito, S.; Page, J.E.; Schmidt, J.; De-Eknamkul, W.; Kutchan, T.M. Aromatic and pyrone polyketides synthesized by a stilbene synthase from Rheum tataricum. Phytochemistry 2003, 62, 313–323. [Google Scholar] [CrossRef]
- Mei, Y.Z.; Liu, R.X.; Wang, D.P.; Wang, X.; Dai, C.C. Biocatalysis and biotransformation of resveratrol in microorganisms. Biotechnol. Lett. 2015, 37, 9–18. [Google Scholar] [CrossRef]
- Nopo-Olazabal, C.; Hubstenberger, J.; Nopo-Olazabal, L. Antioxidant activity of selected stilbenoids and their bioproduction in hairy root cultures of muscadine grape (Vitis rotundifolia Michx.). J. Agric. Food Chem. 2013, 61, 11744–11758. [Google Scholar] [CrossRef]
- Almagro, L.; Belchí-Navarro, S.; Sabater-Jara, A.B.; Vera-Urbina, J.C.; SellésMarchart, S.; Bru, R.; Pedreño, M.A. Bioproduction of trans-resveratrol from grapevine cell cultures. In Natural Products; Ramawat, K., Mérillon, J.M., Eds.; Springer: Berlin, Germany, 2013; pp. 1683–1713. [Google Scholar]
- Liu, C.C.; Zhang, L.L.; An, J.; Chen, B.; Yang, H. Recent strategies for efficient production of polyhydroxyalkanoates by micro-organisms. Lett. Appl. Microbiol. 2016, 62, 9–15. [Google Scholar] [CrossRef]
- Quideau, S.; Deffieux, D.; Douat-Casassus, C.; Pouységu, L. Plant polyphenols: Chemical properties, biological activities, and synthesis. Angew. Chem.-Int. Ed. 2011, 50, 586–621. [Google Scholar] [CrossRef]
- Solladie, G.; Pasturel-Jacope, Y.; Maignan, J. A re-investigation of resveratrol synthesis by Perkins reaction. Application to the synthesis of aryl cinnamic acids. Tetrahedron 2003, 59, 3315–3321. [Google Scholar] [CrossRef]
- Saraswati, S.V.; Thomas, N.F.; Weber, J.F. Strategies and methods for the syntheses of natural oligomeric stilbenoids and analogues. Curr. Org. Chem. 2012, 16, 605–662. [Google Scholar] [CrossRef]
- Nandagopal, K.; Halder, M.; Dash, B.; Nayak, S.; Jha, S. Biotechnological approaches for production of anti-cancerous compounds resveratrol, podophyllotoxin and zerumbone. Curr. Med. Chem. 2018, 25, 4693–4717. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Shao, Z.; Zhao, H. Engineering microbial factories for synthesis of value-added products. J. Ind. Microbiol. Biotechnol. 2013, 38, 873–890. [Google Scholar] [CrossRef] [PubMed]
- Halls, C.; Yu, O. Potential for metabolic engineering of resveratrol biosynthesis. Trends Biotechnol. 2008, 26, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Beekwilder, J.; Wolswinkel, R.; Jonker, H.; Hall, R.; de Vos, C.R.; Bovy, A. Production of resveratrol in recombinant microorganisms. Appl. Environ. Microbiol. 2006, 72, 5670–5672. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Schneider, K.; Kristensen, M. Engineering yeast for high-level production of stilbenoid antioxidants. Sci. Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.Y.; Han, N.S.; Park, Y.C.; Kim, M.D.; Seo, J.H. Production of resveratrol from p-coumaric acid in recombinant Saccharomyces cerevisiae expressing 4-coumarate: Coenzyme A ligase and stilbene synthase genes. Enzyme Microb. Technol. 2011, 48, 48–53. [Google Scholar] [CrossRef]
- Lim, C.G.; Fowler, Z.L.; Hueller, T. High-yield resveratrol production in engineered Escherichia coli. Appl. Environ. Microbiol. 2011, 77, 3451–3460. [Google Scholar] [CrossRef] [PubMed]
- Choi, O.; Wu, C.-Z.; Kang, S.Y. Biosynthesis of plant-specifc phenylpropanoids by construction of an artifcial biosynthetic pathway in Escherichia coli. J. Ind. Microbiol. Biotechnol. 2011, 38, 1657–1665. [Google Scholar] [CrossRef]
- Wu, J.; Liu, P.; Fan, Y.; Bao, H.; Du, G.; Zhou, J.; Chen, J. Multivariate modular metabolic engineering of Escherichia coli to produce resveratrol from L-tyrosine. J. Biotechnol. 2013, 167, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Makrides, S.C. Strategies for achieving high-level expression of genes in Escherichia coli. Microbiol. Rev. 1996, 60, 512–538. [Google Scholar] [PubMed]
- Rosano, G.L.; Ceccarelli, E.A. Recombinant protein expression in Escherichia coli: Advances and challenges. Front. Microbiol. 2014, 5, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Wood, K.V.; Morgan, J. Metabolic engineering of the phenylpropanoid pathway in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2005, 71, 2962–2969. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lin, J.; Hu, H. De novo biosynthesis of resveratrol by site-specific integration of heterologous genes in Escherichia coli. FEMS Microbiol. Lett. 2016, 363, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.L.; Prather, K.L.J.; Kluskens, L.D.; Rodrigues, L.R. Heterologous production of curcuminoids. Microbiol. Mol. Biol. Rev. 2015, 79, 39–60. [Google Scholar] [CrossRef] [PubMed]
- Yesilirmak, F.; Sayers, Z. Heterelogous expression of plant genes. Int. J. Plant Genom. 2009. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.P.; Parajuli, P.; Koffas, M.A.G.; Sohng, J.K. Microbial production of natural and non-natural flavonoids: Pathway engineering, directed evolution and systems/synthetic biology. Biotechnol. Adv. 2016, 34, 634–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krivoruchko, A.; Nielsen, J. Production of natural products through metabolic engineering of Saccharomyces cerevisiae. Curr. Opin. Biotechnol. 2015, 35, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Kildegaard, K.R.; Chen, Y.; Rodriguez, A.; Borodina, I.; Nielsen, J. De novo production of resveratrol from glucose or ethanol by engineered Saccharomyces cerevisiae. Metab. Eng. 2015, 32, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sydor, T.; Schaffer, S.; Boles, E. Considerable increase in resveratrol production by recombinant industrial yeast strains with use of rich medium. Appl. Environ. Microbiol. 2010, 76, 3361–3363. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yu, O. Synthetic scaffolds increased resveratrol biosynthesis in engineered yeast cells. J. Biotechnol 2012, 157, 258–260. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.L.; Xue, Z.; Zhu, Q.Q. Inventors; EI du Pont de Nemours and Co, assignee. Method for the production of resveratrol in a recombinant oleaginous microorganism. U.S. Patent No. 7772444B2, 19 May 2005. [Google Scholar]
- Zhang, W.; Yang, Y.; Liu, X.; Liu, C.; Bai, Z. Development of a secretory expression system with high compatibility between expression elements and an optimized host for endoxylanase production in Corynebacterium glutamicum. Microb. Cell. Fact. 2019. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Xu, J.Z.; Zhang, W.G. Metabolic engineering of l-leucine production in Escherichia coli and Corynebacterium glutamicum: A review. Crit. Rev. Biotechnol. 2019, 6, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Zhang, F.; Xu, J.Z.; Zhang, W.G.; Chen, X.L.; Liu, L.M. Improvement of l-Leucine Production in Corynebacterium glutamicum by Altering the Redox Flux. Int. J. Mol. Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Kallscheuer, N.; Vogt, M.; Stenzel, A.; Gätgens, J.; Bott, M.; Marienhagen, J. Construction of a Corynebacterium glutamicum platform strain for the production of stilbenes and (2S)-flavanones. Metab. Eng. 2016, 38, 47–55. [Google Scholar] [CrossRef]
- Braga, A.; Oliveira, J.; Silva, R.; Ferreira, P.; Rocha, I.; Kallscheuer, N.; Marienhagen, J.; Faria, N. Impact of the cultivation strategy on resveratrol production from glucose in engineered Corynebacterium glutamicum. J. Biotechnol. 2018, 265, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Van Summeren-Wesenhagen, P.V.; Marienhagen, J. Metabolic engineering of Escherichia coli for the synthesis of the plant polyphenol pinosylvin. Appl. Environ. Microbiol. 2015, 81, 840–849. [Google Scholar] [CrossRef]
- Park, S.R.; Yoon, J.A.; Paik, J.H.; Park, J.W.; Jung, W.S.; Ban, Y.H.; Kim, E.J.; Yoo, Y.J.; Han, A.R.; Yoon, Y.J. Engineering of plant-specific phenylpropanoids biosynthesis in Streptomyces venezuelae. J. Biotechnol. 2009, 141, 181–188. [Google Scholar] [CrossRef]
- Katz, M.; Smits, H.P.; Förster, J.; Nielsen, J.B.; Evola, S.A. Metabolically engineered cells for the production of resveratrol or an oligomeric or glycosidically-bound derivative thereof. U.S. Patent No. 9,404,129, 4 June 2015. [Google Scholar]
- Marienhagen, J.; Bott, M. Metabolic engineering of microorganisms for the synthesis of plant natural products. J. Biotechnol. 2013, 163, 166–178. [Google Scholar] [CrossRef]
- Kang, S.Y.; Lee, J.K.; Choi, O.; Kim, C.Y.; Jang, J.H.; Hwang, B.Y.; Hong, Y.S. Biosynthesis of methylated resveratrol analogs through the construction of an artificial biosynthetic pathway in E. coli. BMC Biotechnol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Guleria, S.; Koffas, M.A.; Yan, Y. Microbial production of value-added nutraceuticals. Curr. Opin. Biotechnol. 2016, 37, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Lin, Y.; Yan, Y. Caffeic acid production enhancement by engineering a phenylalanine over-producing Escherichia coli strain. Biotechnol. Bioeng. 2013, 110, 3188–3196. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yang, Y.; Yan, Y. Bioproduction of resveratrol. In Biotechnology of Natural Products; Schwab, W., Lange, B., Wüst, M., Eds.; Springer: Cham, Switzerland, 2018; pp. 61–79. [Google Scholar]
- Watts, K.T.; Lee, P.C.; Schmidt-Dannert, C. Biosynthesis of plant-specific stilbene polyketides in metabolically engineered Escherichia coli. BMC Biotechnol. 2006, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, E.; Guo, X.; Meng, Z.; Wang, J.; Sun, J.; Yao, X.; Xun, H. Construction, expression, and characterization of Arabidopsis thaliana 4-CL and Arachis hypogaea RS fusion gene 4-CL: RS in Escherichia coli. World J. Microbiol. Biotechnol. 2015, 31, 1379–1385. [Google Scholar] [CrossRef] [PubMed]
- Katsuyama, Y.; Funa, N.; Horinouchi, S. Precursor-directed biosynthesis of stilbene methyl ethers in Escherichia coli. Biotechnol. J. 2007, 2, 1286–1293. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, S.; Xiao, A.; Rasmussen, M.; Skidmore, C.; Zhan, J. Metabolic engineering of Escherichia coli for the biosynthesis of various phenylpropanoid derivatives. Metab. Eng. 2015, 29, 153–159. [Google Scholar] [CrossRef]
- Afonso, M.S.; Ferreira, S.; Domingues, F.C.; Silva, F. Resveratrol production in bioreactor: Assessment of cell physiological states and plasmid segregational stability. Biotechnol. Rep. 2014, 5, 7–13. [Google Scholar] [CrossRef]
- Katsuyama, Y.; Funa, N.; Miyahisa, I.; Horinouchi, S. Synthesis of unnatural flavonoids and stilbenes by exploiting the plant biosynthetic pathway in Escherichia coli. Chem. Biol. 2007, 14, 613–621. [Google Scholar] [CrossRef]
- Bhan, N.; Xu, P.; Khalidi, O.; Koffas, M.A.G. Redirecting carbon flux into malonyl-CoA to improve resveratrol titers: Proof of concept for genetic interventions predicted by OptForce computational framework. Chem. Eng. Sci. 2013, 103, 109–114. [Google Scholar] [CrossRef]
- Wu, J.; Zhou, P.; Zhang, X.; Dong, M. Efcient de novo synthesis of resveratrol by metabolically engineered Escherichia coli. J. Ind. Microbiol. Biotechnol. 2017, 44, 1083–1095. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, S.Z.; Li, J.; Pan, X.; Cahoon, R.E.; Jaworski, J.G.; Wang, X.; Jez, J.M.; Chen, F.; Yu, O. Using unnatural protein fusions to engineer resveratrol biosynthesis in yeast and mammalian cells. J. Am. Chem. Soc. 2006, 128, 13030–13031. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Halls, C.; Zhang, J.; Matsuno, M.; Zhang, Y.; Yu, O. Stepwise increase of resveratrol biosynthesis in yeast Saccharomyces cerevisiae by metabolic engineering. Metab. Eng. 2011, 13, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, P.; Dudnik, A.; Neves, A.R.; Förster, J. Engineering Lactococcus lactis for stilbene production. In Proceedings of the 28th International Conference on Polyphenols, Vienna, Austria, 11 July 2016. [Google Scholar]
- Choi, O.; Lee, J.K.; Kang, S.Y.; Pandey, R.P.; Sohng, J.K.; Ahn, J.S. Construction of artificial biosynthetic pathways for resveratrol glucoside derivatives. J. Microbiol. Biotechnol. 2014, 24, 614–618. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.J.; An, C.H.; Woo, S.G.; Jeong, H.J.; Kim, Y.M.; Park, S.J.; Yoon, B.D.; Kim, Y. Production of pinostilbene compounds by the expression of resveratrol O-methyltransferase genes in Escherichia coli. Enzyme Microb. Tech. 2014, 54, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Salas-Navarrete, C.; Hernandez-Chavez, G.; Flores, N.; Martinez, L.M.; Martinez, A.; Bolivar, F.; Barona-Gomez, F.; Gosset, G. Increasing pinosylvin production in Escherichia coli by reducing the expression level of the gene fabI-encoded enoyl-acid carrier protein reductase. Electron. J. Biotechnol. 2018, 33, 11–16. [Google Scholar] [CrossRef]
- Kallscheuer, N.; Vogt, M.; Marienhagen, J. Functional expression of plant-derived methyltransferase, flavanone 3-hydroxylase, and flavonol synthase in Corynebacterium glutamicum for production of pterostilbene, kaempferol, and quercetin. J. Biotechnol. 2017, 258, 190–196. [Google Scholar] [CrossRef]
- Wang, Y.; Bhuiya, M.W.; Zhou, R.; Yu, O. Pterostilbene production by microorganisms expressing resveratrol O-methyltransferase. Ann. Microbiol. 2014, 65, 817–826. [Google Scholar] [CrossRef]
- Takamura, Y.; Nomura, G. Changes in the intracellular concentration of Acetyl-CoA and Malonyl-CoA in relation to the carbon and energy metabolism of Escherichia coli K12. J. Gen. Microbiol. 1988, 134, 2249–2253. [Google Scholar] [CrossRef]
- Zha, W.; Rubin-Pitel, S.B.; Shao, Z.; Zhao, H. Improving cellular malonyl-CoA level in Escherichia coli via metabolic engineering. Metab. Eng. 2009, 11, 92–198. [Google Scholar] [CrossRef]
- Finzel, K.; Lee, D.J.; Burkart, M.D. Using modern tools to probe the structure–function relationship of fatty acid synthases. ChemBioChem 2015, 16, 528–547. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Shao, D.; Shi, J.; Huang, Q.; Yang, H.; Jin, M. Strategies for enhancing resveratrol production and the expression of pathway enzymes. Appl. Microbiol. Biotechnol. 2016, 100, 7407–7421. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.N.S.; Koffas, M.; Stephanopoulos, G. Optimization of a heterologous pathway for the production of flavonoids from glucose. Metab. Eng. 2011, 13, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Subrahmanyam, S., Jr.; Cronan, J.E. Overproduction of a functional fatty acid biosynthetic enzyme blocks fatty acid synthesis in Escherichia coli. J. Bacteriol. 1998, 180, 4596–4602. [Google Scholar] [PubMed]
- de Fouchecour, F.; Sanchez-Castaneda, A.K.; Saulou-Berion, C.; Spinnler, H.E. Process engineering for microbial production of 3-hydroxypropionic acid. Biotechnol. Adv. 2018, 36, 1207–1222. [Google Scholar] [CrossRef] [PubMed]
- Pickens, L.B.; Tang, Y.; Chooi, Y.H. Metabolic engineering for the production of natural products. Annu. Rev. Chem. Biomol. Eng. 2011, 2, 211–236. [Google Scholar] [CrossRef]
- Becker, J.V.; Armstrong, G.O.; van der Merwe, M.J.; Lambrechts, M.G.; Vivier, M.A.; Pretorius, I.S. Metabolic engineering of Saccharomyces cerevisiae for the synthesis of the wine-related antioxidant resveratrol. FEMS Yeast Res. 2003, 4, 79–85. [Google Scholar] [CrossRef]
- Juminaga, D.; Baidoo, E.E.; Redding-Johanson, A.M.; Batth, T.S.; Burd, H.; Mukhopadhyay, A.; Petzold, C.J.; Keasling, J.D. Modular engineering of L-tyrosine production in Escherichia coli. Appl. Environ. Microbiol. 2012, 78, 89–98. [Google Scholar] [CrossRef]
- Zhang, H.; Stephanopoulos, G. Engineering, E. coli for caffeic acid biosynthesis from renewable sugars. Appl. Microbiol. Biotechnol. 2013, 97, 3333–3341. [Google Scholar] [CrossRef]
- Bulter, T.; Bernstein, J.R.; Liao, J.C. A perspective of metabolic engineering strategies: Moving up the systems hierarchy. Biotechnol. Bioeng. 2003, 84, 815–821. [Google Scholar] [CrossRef]
- Lütke-Eversloh, T.; Stephanopoulos, G. L-Tyrosine production by deregulated strains of Escherichia coli. Appl. Microbiol. Biotechnol. 2007, 75, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Na, D.; Yoo, S.M.; Chung, H.; Park, H.; Park, J.H.; Lee, S.Y. Metabolic engineering of Escherichia coli using synthetic small regulatory RNAs. Nat. Biotechnol. 2013, 31, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wu, B.H.; Liu, Z.N.; Qiao, J.; Zhao, G.R. Combinatorial Optimization of Resveratrol Production in Engineered, E. coli. J. Agric. Food Chem. 2018, 66, 13444–13453. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Ranganathan, S.; Fowler, Z.L. Genome-scale metabolic network modeling results in minimal interventions that cooperatively force carbon flux towards malonyl-CoA. Metab. Eng. 2011, 13, 578–587. [Google Scholar] [CrossRef] [PubMed]
- Milke, L.; Aschenbrenner, J.; Marienhagen, J.; Kallscheuer, N. Production of plant-derived polyphenols in microorganisms: Current state and perspectives. Appl. Microbiol. Biotechnol. 2018, 102, 1575–1585. [Google Scholar] [CrossRef]
- Berg, J.; Tymoczko, J.; Stryer, L. Biochemistry; W. H. Freeman and Company: New York, NY, USA, 2002; p. 10010. [Google Scholar]
- Fowler, Z.L.; Gikandi, W.W.; Koffas, M.A.G. Increased malonyl coenzyme A biosynthesis by tuning the Escherichia coli metabolic network and its application to flavanone production. Appl. Environ. Microbiol. 2015, 75, 5831–5839. [Google Scholar] [CrossRef]
- Shrestha, A.; Pandey, R.P.; Pokhrel, A.R.; Dhakal, D.; Chu, L.L.; Sohng, J.K. Modular pathway engineering for resveratrol and piceatannol production in engineered Escherichia coli. Appl. Microbiol. Biotechnol. 2018, 102, 9691–9706. [Google Scholar] [CrossRef]
- An, J.H.; Kim, Y.S. A gene cluster encoding malonyl-CoA decarboxylase (MatA), malonyl-CoA synthetase (MatB) and a putative dicarboxylate carrier protein (MatC) in Rhizobium trifolii: Cloning, sequencing, and expression of the enzymes in Escherichia coli. Eur. J. Biochem. 1998, 257, 395–402. [Google Scholar] [CrossRef]
- Braga, A.; Silva, M.; Oliveira, J.; Silva, A.R.; Ferreira, P.; Ottens, M.; Rocha, I.; Faria, N. An adsorptive bioprocess for production and recovery of resveratrol with Corynebacterium glutamicum. J. Chem. Technol. Biotechnol. 2018, 93, 1661–1668. [Google Scholar] [CrossRef]
- Lucas-Abellán, C.; Fortea, I.; López-Nicolás, J.M.; Núñez-Delicado, E. Cyclodextrins as resveratrol carrier system. Food Chem. 2007, 104, 39–44. [Google Scholar] [CrossRef]
- Trela, B.C.; Waterhouse, A.L. Resveratrol: Isomeric molar absorptivities and stability. J. Agric. Food Chem. 1996, 44, 1253–1257. [Google Scholar] [CrossRef]
- Zhou, K.; Qiao, K.; Edgar, S.; Stephanopoulo, G. Distributing a metabolic pathway among a microbial consortium enhances production of natural products. Nat. Biotechnol. 2015, 33, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Hua, D.; Ma, C.; Song, L. Enhanced vanillin production from ferulic acid using adsorbent resin. Appl. Microbiol. Biotechnol. 2007, 74, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Jin, Y.; Ho, C.T. Evaluation of resveratrol derivatives as potential antioxidants and identification of a reaction product of resveratrol and 2, 2-diphenyl-1-picryhydrazyl radical. J. Agric. Food Chem. 1999, 47, 3974–3977. [Google Scholar] [CrossRef] [PubMed]
- Fauconneau, B.; Waffo-Teguo, P.; Huguet, F.; Barrier, L.; Decendit, A.; Merillon, J.M. Comparative study of radical scavenger and antioxidant properties of phenolic compounds from Vitis vinifera cell cultures using in vitro tests. Life Sci. 1997, 61, 2103–2110. [Google Scholar] [CrossRef]
- Lee, S.K.; Nam, K.A.; Hoe, Y.H.; Min, H.Y.; Kim, E.Y.; Ko, H.; Song, S.; Lee, T.; Kim, S. Synthesis and evaluation of cytotoxicity of stilbene analogues. Arch. Pharm. Res. 2003, 26, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Hall, D.; De Luca, V. Mesocarp localization of a bi-functional resveratrol/hydroxycinnamic acid glucosyltransferase of Concord grape (Vitis labrusca). Plant J. 2007, 49, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Schmidlin, L.; Poutaraud, A.; Claudel, P.; Mestre, P.; Prado, E.; SantosRosa, M.; Wiedemann-Merdinoglu, S.; Karst, F.; Merdinoglu, D.; Hugueney, P.A. Stress-inducible resveratrol O-methyltransferase involved in the biosynthesis of pterostilbene in grapevine. Plant. Physiol. 2008, 148, 1630–1639. [Google Scholar] [CrossRef]
- Jeandet, P.; Douillet-Breuil, A.C.; Bessis, R.; Debord, S.; Sbaghi, M.; Adrian, M. Phytoalexins from the Vitaceae: Biosynthesis, phytoalexin gene expression in transgenic plants, antifungal activity, and metabolism. J. Agric. Food. Chem. 2002, 50, 2731–2741. [Google Scholar] [CrossRef]
- Yu, C.K.; Lam, C.N.; Springob, K.; Schmidt, J.; Chu, I.K.; Lo, C. Constitutive accumulation of cis-piceid in transgenic Arabidopsis overexpressing a sorghum stilbene synthase gene. Plant. Cell Physiol. 2006, 47, 1017–1021. [Google Scholar] [CrossRef]
- Jeong, Y.J.; Woo, S.G.; An, C.H.; Jeong, H.J.; Hong, Y.S.; Kim, Y.M.; Ryu, Y.B.; Rho, M.C.; Lee, W.S.; Kim, C.Y. Metabolic engineering for resveratrol derivative biosynthesis in Escherichia coli. Mol. Cells 2015, 38, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.Y.; Choi, O.; Lee, J.K.; Hwang, B.Y.; Uhm, T.B.; Hong, Y.S. Artificial biosynthesis of phenylpropanoic acids in a tyrosine overproducing Escherichia coli strain. Microb. Cell Fact. 2012. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Yan, Y. Biotechnological production of plant-specific hydroxylated phenylpropanoids. Biotechnol. Bioeng. 2014, 111, 1895–1899. [Google Scholar] [CrossRef] [PubMed]
- Furuya, T.; Miura, M.; Kino, K. A Coenzyme-Independent Decarboxylase/Oxygenase Cascade for the Efficient Synthesis of Vanillin. ChemBioChem 2014, 15, 2248–2254. [Google Scholar] [CrossRef] [PubMed]
- Rimal, H.; Yu, S.C.; Lee, J.H.; Yamaguchi, T.; Oh, T.J. Hydroxylation of resveratrol with DoxA in vitro: An enzyme with the potential for the bioconversion of a bioactive stilbene. J. Microbiol. Biotechnol. 2018, 28, 561–565. [Google Scholar] [PubMed]
- Yang, T.; Fang, L.; Sanders, S.; Jayanthi, S.; Rajan, G.; Podicheti, R.; Medina-Bolivar, F. Stilbenoid prenyltransferases define key steps in the diversification of peanut phytoalexins. J. Biolog. Chem. 2018, 293, 28–46. [Google Scholar] [CrossRef] [Green Version]
- Araya-Cloutier, C.; Martens, B.; Schaftenaar, G.; Leipoldt, F.; Gruppen, H.; Vincken, J.P. Structural basis for non-genuine phenolic acceptor substrate specificity of Streptomyces roseochromogenes prenyltransferase CloQ from the ABBA/PT-barrel superfamily. PLoS ONE 2017. [Google Scholar] [CrossRef]
- Botta, B.; Delle Monache, G.; Menendez, P.; Boffi, A. Novel prenyltransferase enzymes as a tool for flavonoid prenylation. Trends Pharmacol. Sci. 2005, 26, 606–608. [Google Scholar] [CrossRef]
- Kuzuyama, T.; Noel, J.P.; Richard, S.B. Structural basis for the promiscuous biosynthetic prenylation of aromatic natural products. Nature 2005, 435, 983–987. [Google Scholar] [CrossRef] [Green Version]
- Li, S.M. Evolution of aromatic prenyltransferases in the biosynthesis of indole derivatives. Phytochemistry 2009, 70, 1746–1757. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Name | Source | Reference |
|---|---|---|
| VvSTS | Vitis vinifera | [50] |
| Pcu1STS | Polygonum cuspidatum | [51] |
| Pcu3STS | Polygonum cuspidatum | [51] |
| PsSTS | Pinus strobus | [52] |
| PdSTS | Pinus densiflora | [53] |
| AhSTS | Arachis hypogaea | [54] |
| PhSTS | Parthenocissus henryana | [55] |
| MaSTS | Morus atropurpurea | [56] |
| RtSTS | Rheum tataricum | [57] |
| Microbial Host | Incorporated Genes | Host Engineered | Substrate | Titer (mg/L) | References |
|---|---|---|---|---|---|
| Escherichia coli(Resveratrol) | |||||
| E. coli BW27784 | 4CL(A. thaliana) STS (A. hypogaea) | p-Coumaric acid | 0.16 | [103] | |
| E. coli BL21(DE3) | 4CL (N. tabacum) STS (V. vinifera) | p-Coumaric acid | 16 | [68] | |
| E. coli BL21(DE3) | Pal (R. rubra) 4CL (L. erythrorhizon) STS (A. hypogaea) | Tyrosine | 37 | [101] | |
| E. coli BRB | 4CL (L. erythrorhizon) STS (A. hypogaea) ACC (C. glutamicum) F3H and FLS (Citrus) | Cinnamic acid | 155 | [104] | |
| p-Coumaric acid | 171 | ||||
| E. coli BW27784 | 4CL (A. thaliana) STS (A. hypogaea) | p-Coumaric acid | 105 | [99] | |
| E. coli BW27784 | 4CL (A. thaliana) STS (A. hypogaea) | p-Coumaric acid | 404 | [71] | |
| 4CL (A. thaliana) STS (V. vinífera) | 1380 | ||||
| 4CL (P. crispum) STS (A. hypogaea) | 142 | ||||
| 4CL (P. crispum) STS (V. vinífera) | 610 | ||||
| 4CL (A. thaliana) STS (V. vinífera) | p-Coumaric acid and cerulenin | 2340 | [71] | ||
| E. coli C41 (DE3) | TAL (S. espanaensis) 4CL (S. coelicolor) STS (A. hypogaea) | p-Coumaric acid | 104 | [72] | |
| E. coli BW25113 (DE3) | TAL (R. glutinis) 4CL (P. crispum) STS (V. vinifera) | Inactivation of tyrR and deletion of trpED by chromosomal integration | Glucose | 4.6 | [77] |
| E. coli BW27784 (DE3) | 4CL (A. thaliana) STS (V. vinífera) | p-Coumaric acid | 1600 | [105] | |
| E. coli BL21 (DE3) | TAL (R. glutinis) 4CL (P. crispum) STS (V. vinifera) matB and matC (R. trifolii) | L-Tyrosine | 35.02 | [73] | |
| E. coli BL21 (DE3) | 4CL::STS, 4CL (A. thaliana)- STS (A. hypogaea) fusion enzyme | p-Coumaric acid | 80.5 | [100] | |
| E. coli BL21 (DE3) | TAL (T. cutaneum) 4CL (P. crispum) STS (V. vinifera) matB and matC (R. trifolii) tyrAfbr and aroGfbr (E. coli K12) | Down-regulation of fabD, fabH, fabB, fabF, fabI | Glucose | 304.5 | [106] |
| E. coli BW25113 | 4CL2 (P.crispum) STS (V. vinifera) | p-Coumaric acid | 268.2 | [14] | |
| E. coli C41 (DE3) | TAL (S. espanaensis) 4CL (S. coelicolor) STS (A. hypogaea) | Glucose | 5.2 | [95] | |
| E. coli BL21(DE3) | TAL (S. espanaensis) 4CL (A. thaliana) STS (A. hypogaea) | Tyrosine | 114.2 | [102] | |
| E. coli W(pheA-) Rg E.coli W-Vv | TAL (R. glutinis) tktAfbr and aroG fbr (E. coli) 4CL (S. coelicolor) STS (V. vinífera) | Deletion of pheA | Glycerol | 22.58 | [49] |
| Non- E. coli (Resveratrol) | |||||
| C. glutamicum DelAro3 | STS (A. hypogaea) 4CL (P. crispum) | Deletion of phdB, pcaF and pobA | p-Coumaric acid | 12 | [89] |
| p-coumaric acid + cerulenin | 158 | ||||
| C. glutamicum DelAro3 | TAL (F. johnsoniae) 4CL (P. crispum) STS (A. hypogaea) aroH (E. coli) | Deletion of phdB, pcaF, qsuB and pobA | Glucose | 12 | [90] |
| Glucose+ cerulenin | 59 | ||||
| Glucose(40 g/L) | 4 | ||||
| Glucose(80 g/L) | 12 | ||||
| Glucose (Fed-batch) | 7 | ||||
| S. cerevisiae W303-1A | 4CL (A. thaliana) STS (A. hypogaea) | p-Coumaric acid | 3.1 | [70] | |
| S. cerevisiae WAT11 | 4CL (A. thaliana):STS (V. vinifera) | p-Coumaric acid | 5.25 | [107] | |
| S. cerevisiae WAT11 | TAL (R. sphaeroides) 4CL::STS, 4CL (A. thaliana)-STS V. vinifera) fusionenzyme | Tyrosine | 1.9 | [108] | |
| S. cerevisiae CEN. PK102-5B | TAL (H. aurantiacus) 4CL (A. thaliana) VST (V. vinifera) | Overexpression of aro4, aro7, and acc1 | Glucose (fed-batch) | 415.65 | [82] |
| Ethanol (fed-batch) | 531.41 | ||||
| S. cerevisiae ST4990 | PAL (A. thaliana) C4H (A. thaliana) 4CL (A. thaliana) VST (V. vinifera) ACS (S. enterica) Overexpression of atr2 (A. thaliana) | Overexpression of aro4, aro7, and acc1 and deletion of aro10 | Glucose (Fed-batch) | 812 | [69] |
| S. cerevisiae DHS2001 | STS (A. hypogaea) 4CL (S. coelicolor) | Deletion of pks | p-Coumaric acid | 0.4 | [92] |
| L.lactis | TAL, 4CL, STS, ACC (different sources) | L-Tyrosine | 0.45-1.37 | [109] | |
| Industrial Brazilian S. cerevisiae strain | STS (V. vinifera) 4CL (A. thaliana) | p-Coumaric acid | 391 | [83] | |
| Escherichiacoli (Resveratrol derivatives) | Substrate | Derivatives | |||
| E. coli C41 (DE3) | TAL (S. espanaensis) 4CL (S. coelicolor) STS (A. hypogea) Glycosyltransferase YjiC (Bacillus spp.) | Glucose | 3-O-resveratrol β-D-glucoside | 2.5 | [110] |
| 4′-O-resveratrol β-D-glucoside | 7.5 | ||||
| E. coli BL21-CodonPlus (DE3)-RIPL | VvROMT (V. riparia) | Resveratrol | Pinostilbene | 0.16 | [111] |
| Pterostilbene | 0.04 | ||||
| SbROMT (S. bicolor) | Pinostilbene | 34 | |||
| Pterostilbene | 0.16 | ||||
| E. coli C41(DE3) | TAL (S. espanaensis) 4CL (S. coelicolor) STS (A. hypogea) SbOMT3, SbOMT1 | Glucose | 3,5-Dihydroxy-4′-methoxystilbene | 2.50.2 | [95] |
| 5-Hydroxy-3,4′-dimethoxystilbene | |||||
| E. coli W3110 | 4CL(S.coelicolor A3), STS (V.vinifera) | Cinnamic acid | Pinosylvin | 34.89 | [112] |
| 4CL(S. coelicolor A3), STS (V. vinifera) + reduced expression level of fabI gene | Cinnamic acid | Pinosylvin | 52.67 | ||
| E. coli BW27784 | 4CL1(A. thaliana) STS(A. hypogea) | Caffieic acid | Piceatannol | 13 | [99] |
| E. coli BLR(DE3) | PAL(Rhodotorula rubra) 4CL(Lithospermum erythrorhizon) STS(A. hypogea) ACC(Corynebacterium glutamicum) | Phenylalanine | Pinosylvin | 20 | [104] |
| Tyrosine | Pinostilbene | 18 | |||
| Pterositibene | 5.8 | ||||
| PAL(Rhodotorula rubra) 4CL(Lithospermum erythrorhizon) STS(A. hypogea) ACC(Corynebacterium glutamicum) +OsPMT (Oryza sativa) | Phenylalanine | Pinosylvin monomethyether | 27 | ||
| +Pinosylvin dimethyl ether | 27 | ||||
| E. coli BL21(DE3) | PAL (P. crispum) 4CL(S.coelicolor) STS [Pinus strobus (two mutated T248A; Q361R)] Cerulenin | Glucose/phenylalanine | Pinosylvin | 91 | [91] |
| Non-E. coli hosts (Resveratrol derivatives) | |||||
| C. glutamicum DelAro4 | 4CL (P. crispum), STS (A. hypogea) OMT (V. vinifera):MalE (E. coli) metK (E. coli) | p-Coumaric acid | Pterostilbene | 42 | [113] |
| S. cerevisiae WAT11 | 4CL (A. thaliana), STS (V. vinifera) ROMT (V. vinifera) | Resveratrol | Pterostilbene | 150 | [114] |
| p-Coumaric acid | Pterostilbene | 2 | |||
| S. cerevisiae ST4993 | thaliana (PAL, C4H, 4CL2) STS (V. vinifera) + feedback insensitive alleles of DHAP synthase (ScARO4K229L) and chorismate mutase (ScArO7G141S) + shikimate kinase aroL + deregulated variant of ACC1 + non-regulated version of acetyl-CoA synthase (ACS) + deletion of phenylpyruvate decarboxylase ARO10 + cytochrome P450 reductases ATR2 and CYB5 + SbROMT (Sorghum bicolor) | Glucose | Pterostilbene | 5.5 | [69] |
| S. cerevisiae ST4994 | All the components of Strain ST4993 along with VvROMT(V.vinifera) instead of SbROMT | Pterostilbene | 3.5 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thapa, S.B.; Pandey, R.P.; Park, Y.I.; Sohng, J.K. Biotechnological Advances in Resveratrol Production and its Chemical Diversity. Molecules 2019, 24, 2571. https://doi.org/10.3390/molecules24142571
Thapa SB, Pandey RP, Park YI, Sohng JK. Biotechnological Advances in Resveratrol Production and its Chemical Diversity. Molecules. 2019; 24(14):2571. https://doi.org/10.3390/molecules24142571
Chicago/Turabian StyleThapa, Samir Bahadur, Ramesh Prasad Pandey, Yong Il Park, and Jae Kyung Sohng. 2019. "Biotechnological Advances in Resveratrol Production and its Chemical Diversity" Molecules 24, no. 14: 2571. https://doi.org/10.3390/molecules24142571
APA StyleThapa, S. B., Pandey, R. P., Park, Y. I., & Sohng, J. K. (2019). Biotechnological Advances in Resveratrol Production and its Chemical Diversity. Molecules, 24(14), 2571. https://doi.org/10.3390/molecules24142571