Anti-Diabetic Effect of a Shihunine-Rich Extract of Dendrobium loddigesii on 3T3-L1 Cells and db/db Mice by Up-Regulating AMPK–GLUT4–PPARα


Abstract
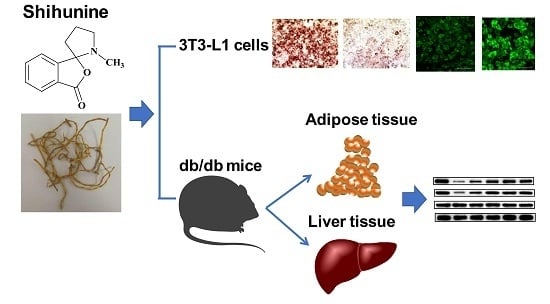
:
1. Introduction
2. Results
2.1. Composition of a Shihunine-Rich Extract of D. loddigesii
2.2. Effect of a Shihunine-Rich Extract of D. loddigesii on the 3T3-L1 Preadipocyte
2.2.1. Inhibition of Preadipocyte Differentiation and Lipogenesis
2.2.2. Improvement of Glucose Uptake
2.3. Effect of a Shihunine-Rich Extract of D. loddigesii on Db/db Mice
2.3.1. Effects of a Shihunine-Rich Extract of D. loddigesii on Body Weight, Blood Glucose Level, and Oral Glucose Tolerance Test
2.3.2. Effect of a Shihunine-Rich Extract of D. loddigesii on Serum Insulin and Lipid Levels
2.3.3. Effect of a Shihunine-Rich Extract of D. loddigesii on Tissue Forms of Adipose/Pancreas
2.4. Effect of a Shihunine-Rich Extract of D. loddigesii on Cleaved Caspase-3 Protein Expression in the Pancreas
2.5. Effect of a Shihunine-Rich Extract of D. loddigesii on p-AMPK, GLUT4, and PPARα Protein Expressions in Tissue of the Adipose/Liver
2.6. Toxic Effect of a Shihunine-Rich Extract of D. loddigesii on C57BL/6 Mice
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Materials
4.3. Preparation and Identification of the Shihunine-Rich Extract of D. Loddigesii
4.4. Cell Experiments
4.4.1. T3-l1 Cell Culture, Differentiation Induction, and Insulin-Resistant Adipocyte Model
4.4.2. Determination of Intracellular Lipid Content, Triglyceride Content, and Glucose Uptake
4.5. Animal Experiments
4.6. Blood Glucose and Serum Biochemical Indices Analysis
4.7. Histologic Analysis and Immunohistochemical Analysis
4.8. Western Blot Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AMPK | adenosine monophosphate-activated protein kinase |
| ALT | alanine aminotransferase |
| AST | aspartate aminotransferase |
| control | no-treatment C57 mice |
| cleaved caspase-3 | cleaved cysteine aspartic acid-specific protease 3 |
| DLS | shihunine-rich extract of Dendrobium loddigesii |
| DLS200 | DLS-treatment C57 mice, in a dose of 200 mg/kg |
| DM | no-treatment diabetes db/db mice |
| DMDLS100 | DLS-treatment diabetes db/db mice, in a dose of 100 mg/kg |
| DMDLS25 | DLS-treatment diabetes db/db mice, in a dose of 25 mg/kg |
| DMDLS50 | DLS-treatment diabetes db/db mice, in a dose of 50 mg/kg |
| DMMET130 | MET-treatment diabetes db/db mice, in a dose of 130 mg/kg |
| FBG | fasting blood glucose |
| GLUT4 | glucose transporter 4 |
| HDL-C | high-density lipoprotein cholesterol |
| INS | insulin |
| IOD | integrated optical density |
| LDL-C | low-density lipoprotein cholesterol |
| MET | metformin |
| OGTT | oral glucose tolerance test |
| p-AMPK | adenosine monophosphate-activated protein kinase phosphorylation |
| PPARα | peroxisome proliferator-activated receptor α |
| T2DM | type 2 diabetes mellitus |
| TC | total cholesterol |
| TG | triglyceride |
| UA | uric acid |
| CRE | creatinine |
References
- Ogurtsova, K.; da Rocha Fernandes, J.D.; Huang, Y.; Linnenkamp, U.; Guariguata, L.; Cho, N.H.; Cavan, D.; Shaw, J.E.; Makaroff, L.E. IDF diabetes atlas: Global estimates for the prevalence of diabetes for 2015 and 2040. Diabetes Res. Clin. Pract. 2017, 128, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Boussageon, R.; Roustit, M.; Gueyffier, F.; Tudrej, B.V.; Rehman, M.B. Type 2 diabetes. Lancet 2018, 391, 1261. [Google Scholar] [CrossRef]
- Chatterjee, S.; Khunti, K.; Davies, M.J. Type 2 diabetes. Lancet 2017, 389, 2239–2251. [Google Scholar] [CrossRef]
- Kahn, S.E.; Hull, R.L.; Utzschneider, K.M. Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 2006, 444, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Reaven, G.M. Role of insulin resistance in human disease. In Multiple Risk Factors in Cardiovascular Disease. Medical Science Symposia Series; Gotto, A.M., Lenfant, C., Paoletti, R., Soma, M., Eds.; Springer: Dordrecht, The Netherlands, 1992; Volume 1, pp. 91–97. [Google Scholar]
- Kahn, S.E. Clinical review 135: The importance of β-cell failure in the development and progression of type 2 diabetes. J. Clin. Endocrinol. Metab. 2001, 86, 4047–4058. [Google Scholar] [PubMed]
- Petersen, M.C.; Shulman, G.I. Mechanisms of insulin action and insulin resistance. Physiol. Rev. 2018, 98, 2133–2223. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.B.; Zhou, G.; Li, C. AMPK: an emerging drug target for diabetes and the metabolic syndrome. Cell Metab. 2009, 9, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Coughlan, K.A.; Valentine, R.J.; Ruderman, N.B.; Saha, A.K. AMPK activation: A therapeutic target for type 2 diabetes? Diabetes Metab. Syndr. Obes. 2014, 7, 241–253. [Google Scholar] [PubMed]
- Gual, P.; Le Marchand-Brustel, Y.; Tanti, J.F. Positive and negative regulation of insulin signaling through IRS-1 phosphorylation. Biochimie 2005, 87, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Kola, B.; Boscaro, M.; Rutter, G.A.; Grossman, A.B.; Korbonits, M. Expanding role of AMPK in endocrinology. Trends Endocrinol. Metab. 2006, 17, 205–215. [Google Scholar] [CrossRef]
- Cool, B.; Zinker, B.; Chiou, W.; Kifle, L.; Cao, N.; Perham, M.; Dickinson, R.; Adler, A.; Gagne, G.; Iyengar, R.; et al. Identification and characterization of a small molecule AMPK activator that treats key components of type 2 diabetes and the metabolic syndrome. Cell Metab. 2006, 3, 403–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantó, C.; Auwerx, J. AMP-activated protein kinase and its downstream transcriptional pathways. Cell Mol. Life Sci. 2010, 67, 3407–3423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Czech, M.P. The GLUT4 glucose transporter. Cell Metab. 2007, 5, 237–352. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, P.R.; Gnudi, L.; Tozzo, E.; Yang, H.; Leach, F.; Kahn, B.B. Adipose cell hyperplasia and enhanced glucose disposal in transgenic mice overexpressing GLUT4 selectively in adipose tissue. J. Biol. Chem. 1993, 268, 22243–22246. [Google Scholar] [PubMed]
- Tozzo, E.; Shepherd, P.R.; Gnudi, L.; Kahn, B.B. Metabolism, transgenic GLUT-4 over expression in fat enhances glucose metabolism: Preferential effect on fatty acid synthesis. Am. J. Physiol. 1995, 268, E956–E964. [Google Scholar]
- Poulsen, L.; Siersbaek, M.; Mandrup, S. PPARs: Fatty acid sensors controlling metabolism. Semin. Cell Dev. Biol. 2012, 23, 631–639. [Google Scholar] [CrossRef]
- Berge, R.K.; Tronstad, K.J.; Berge, K.; Rost, T.H.; Wergedahl, H.; Gudbrandsen, O.A.; Skorve, J. The metabolic syndrome and the hepatic fatty acid drainage hypothesis. Biochimie 2005, 87, 15–20. [Google Scholar] [CrossRef]
- Martin, G.; Schoonjans, K.; Lefebvre, A.M.; Staels, B.; Auwerx, J. Coordinate regulation of the expression of the fatty acid transport protein and Acyl-CoA synthetase genes by PPARα and PPARγ activators. J. Biol. Chem. 1997, 272, 28210–28217. [Google Scholar] [CrossRef]
- Brandt, J.M.; Djouadi, F.; Kelly, D.P. Fatty acids activate transcription of the muscle carnitine palmitoyl transferase I gene in cardiac myocytes via the peroxisome proliferator-activated receptor α. J. Biol. Chem. 1998, 273, 23786–23792. [Google Scholar] [CrossRef]
- Houten, S.M.; Chegary, M.; Te Brinke, H.; Wijnen, W.J.; Glatz, J.F.; Luiken, J.J.; Wijburg, F.A.; Wanders, R.J. Pyruvate dehydrogenase kinase 4 expression is synergistically induced by AMP-activated protein kinase and fatty acids. Cell Mol. Life Sci. 2009, 66, 1283–1294. [Google Scholar] [CrossRef]
- Bronner, M.; Hertz, R.; Bar-Tana, J. Kinase-independent transcriptional co-activation of peroxisome proliferator-activated receptor α by AMP-activated protein kinase. Biochem. J. 2004, 384, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Editorial Committee of Flora of China, CAS. Flora of China; Science Press: Beijing, China, 1999; Volume 19, pp. 104–106. [Google Scholar]
- Wang, H.Y.; Li, Q.M.; Yu, N.J.; Chen, W.D.; Zha, X.Q.; Wu, D.L.; Pan, L.H.; Duan, J.; Luo, J.P. Dendrobium huoshanense polysaccharide regulates hepatic glucose homeostasis and pancreatic beta-cell function in type 2 diabetic mice. Carbohydr. Polym. 2019, 211, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Shi, H.; Cui, Y.; Wang, X.; Zhang, J.; Yu, W.; Wei, M. Dendrobium mixture regulates hepatic gluconeogenesis in diabetic rats via the phosphoinositide-3-kinase/protein kinase B signaling pathway. Exp. Ther. Med. 2018, 16, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, D.; Dou, M.; Li, Z.; Zhang, J.; Zhao, X. Dendrobium officinale Kimura et Migo attenuates diabetic cardiomyopathy through inhibiting oxidative stress, inflammation and fibrosis in streptozotocin-induced mice. Biomed. Pharmacother. 2016, 84, 1350–1358. [Google Scholar] [CrossRef]
- Xu, J.; Han, Q.B.; Li, S.L.; Chen, X.J.; Wang, X.N.; Zhao, Z.Z.; Chen, H.B. Chemistry, bioactivity and quality control of Dendrobium, a commonly used tonic herb in traditional Chinese medicine. Phytochem. Rev. 2013, 12, 341–367. [Google Scholar] [CrossRef]
- Chen, H.; Li, X.; Xu, Y.; Lo, K.; Zheng, H.; Hu, H.; Wang, J.; Lin, Y. Study on the polar extracts of Dendrobium nobile, D. officinale, D. loddigesii, and Flickingeria fimbriata: metabolite identification, content evaluation, and bioactivity assay. Molecules 2018, 23, 1185. [Google Scholar] [CrossRef] [PubMed]
- Li, X.W.; Chen, H.P.; He, Y.Y.; Chen, W.L.; Chen, J.W.; Gao, L.; Hu, H.Y.; Wang, J. Effects of rich-polyphenols extract of Dendrobium loddigesii on anti-diabetic, anti-inflammatory, anti-oxidant, and gut microbiota modulation in db/db mice. Molecules 2018, 23, 3245. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Qin, X.; Zhang, T.; Li, Q.; Zhang, J.; Zhao, J. Astragalus polysaccharide improves insulin sensitivity via AMPK activation in 3T3-L1 adipocytes. Molecules 2018, 23, 2711. [Google Scholar] [CrossRef]
- Zhang, R.X.; Jia, Z.P.; Li, M.X.; Guo, L.M.; Zhang, X.H. The establishment of insulin resistant model in vitro and preliminary application for screening drugs. Chin. Pharmacol. Bull. 2008, 24, 971–976. [Google Scholar]
- Zhang, W.Y.; Lee, J.J.; Kim, Y.; Kim, I.S.; Han, J.H.; Lee, S.G.; Ahn, M.J.; Jung, S.H.; Myung, C.S. Effect of eriodictyol on glucose uptake and insulin resistance in vitro. J. Agric. Food Chem. 2012, 60, 7652–7658. [Google Scholar] [CrossRef]
- Bahmani, M.; Shirzad, H.; Mirhosseini, M.; Mesripour, A.; Rafieian-Kopaei, M. A review on ethnobotanical and therapeutic uses of Fenugreek (Trigonella foenum-graceum L). J. Evid. Based Complementary Altern. Med. 2016, 21, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, W.; McRae, S.; Marek, G.; Wymer, D.; Pannu, V.; Baylis, C.; Johnson, R.J.; Sautin, Y.Y. Hyperuricemia as a mediator of the proinflammatory endocrine imbalance in the adipose tissue in a murine model of the metabolic syndrome. Diabetes 2011, 60, 1258–1269. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Z.; Chen, B.X.; Xu, Y.G.; Xu, J.Y.; Zhu, X.Y. Clinical analysis of relationship between type 2 diabetes and gastric mucosal lesions. Tianjin Med. J. 2008, 36, 863–865. [Google Scholar]
- Liadis, N.; Murakami, K.; Eweida, M.; Elford, A.R.; Sheu, L.; Gaisano, H.Y.; Hakem, R.; Ohashi, P.S.; Woo, M. Caspase-3-dependent-cell apoptosis in the initiation of autoimmune diabetes mellitus. Mol. Cell. Biol. 2005, 25, 3620–3629. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.M. Protective effects of geniposide on diabetes via inhibiting apoptosis in diabetic rats Islets. J. Clin. Exp. Med. 2017, 16, 741–743. [Google Scholar]
- Huang, M.Q.; Zhou, C.J.; Zhang, Y.P.; Zhang, X.Q.; Xu, W.; Lin, J.; Wang, P.J. Salvianolic acid B ameliorates hyperglycemia and dyslipidemia in db/db mice through the AMPK pathway. Cell Physiol. Biochem. 2016, 40, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Penumathsa, S.V.; Thirunavukkarasu, M.; Zhan, L.; Maulik, G.; Menon, V.P.; Bagchi, D.; Maulik, N. Resveratrol enhances GLUT-4 translocation to the caveolar lipid raft fractions through AMPK/Akt/eNOS signalling pathway in diabetic myocardium. J. Cell Mol. Med. 2008, 12, 2350–2361. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.S.; Kim, W.S.; Kim, K.H.; Yoon, M.J.; Cho, H.J.; Shen, Y.; Ye, J.M.; Lee, C.H.; Oh, W.K.; Kim, C.T.; et al. Berberine, a natural plant product, activates AMP-activated protein kinase with beneficial metabolic effects in diabetic and insulin-resistant states. Diabetes 2006, 55, 2256–2264. [Google Scholar] [CrossRef]
- Pu, P.; Gao, D.M.; Mohamed, S.; Chen, J.; Zhang, J.; Zhou, X.Y.; Zhou, N.J.; Xie, J.; Jiang, H. Naringin ameliorates metabolic syndrome by activating AMP-activated protein kinase in mice feda high-fat diet. Arch. Biochem. Biophys. 2012, 518, 61–70. [Google Scholar] [CrossRef]
- Ko, J.H.; Nam, D.; Um, J.Y.; Jung, S.H.; Ahn, K.S. Bergamottin inhibits adipogenesis in 3T3-L1 Cells and weight regulation in diet-induced obese mice. Am. J. Chin. Med. 2018, 46, 601–615. [Google Scholar] [CrossRef]
- Jia, Y.; Bhuiyan, M.J.; Jun, H.J.; Lee, J.H.; Hoang, M.H.; Lee, H.J.; Kim, N.; Lee, D.; Hwang, K.Y.; Hwang, B.Y.; et al. Ursolic acid is a PPAR-α agonist that regulates hepatic lipid metabolism. Bioorg. Med. Chem. Lett. 2011, 21, 5876–5880. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Kim, S.; Kim, J.; Kim, B.; Wu, C.; Lee, J.H.; Jun, H.J.; Kim, N.; Lee, D.; Lee, S.J. Ursolic acid improves lipid and glucose metabolism in high-fat-fed C57BL/6J mice by activating peroxisome proliferator-activated receptor alpha and hepatic autophagy. Mol. Nutr. Food Res. 2015, 59, 344–354. [Google Scholar] [CrossRef] [PubMed]
- Bak, E.J.; Kim, J.; Choi, Y.H.; Kim, J.H.; Lee, D.E.; Woo, G.H.; Cha, J.H.; Yoo, Y.J. Wogonin ameliorates hyperglycemia and dyslipidemia via PPARα activation in db/db mice. Clin. Nutr. 2014, 33, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Y.; Tsai, S.I.; Damu, A.G.; Wu, T.S. A rapid and simple determination of protoberberine alkaloids in Rhizoma Coptidis by 1H-NMR and its application for quality control of commercial prescriptions. J. Pharm. Biomed. Anal. 2009, 49, 1272–1276. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; He, T.; Han, Y.; Sheng, J.Z.; Jin, S.; Jin, M.W. Pentamethylquercetin improves adiponectin expression in differentiated 3T3-L1 cells via a mechanism that implicates PPARgamma together with TNF-alpha and IL-6. Molecules 2011, 16, 5754–5768. [Google Scholar] [CrossRef] [PubMed]
- Luan, G.; Wang, Y.; Wang, Z.; Zhou, W.; Hu, N.; Li, G.; Wang, H. Flavonoid glycosides from Fenugreek seeds regulate glycolipid metabolism by improving mitochondrial function in 3T3-L1 adipocytes in vitro. J. Agric. Food Chem. 2018, 66, 3169–3178. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Flores, L.M.; Lopez-Briones, S.; Macias-Cervantes, M.H.; Ramirez-Emiliano, J.; Perez-Vazquez, V. A PPARgamma, NF-kappaB and AMPK-dependent mechanism may be involved in the beneficial effects of curcumin in the diabetic db/db mice liver. Molecules 2014, 19, 8289–8302. [Google Scholar] [CrossRef]
Sample Availability: Not available. |



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | TG | TC | HDL-C | LDL-C | INS |
|---|---|---|---|---|---|
| mmol/L | mIU/L | ||||
| DM | 3.34 ± 1.25 | 7.42 ± 1.49 | 2.27 ± 0.44 | 0.79 ± 0.29 | 13.12 ± 2.81 |
| DMMET130 | 2.49 ± 0.73 | 7.08 ± 1.10 | 2.53 ± 0.63 | 1.14 ± 0.48 | 21.29 ± 7.68 * |
| Control | 1.06 ± 0.33 | 3.52 ± 0.64 | 2.11 ± 0.17 | 0.22 ± 0.03 | 25.12 ± 4.81 |
| DMDLS25 | 2.13 ± 0.40 * | 6.11 ± 1.25 | 2.64 ± 0.48 | 0.97 ± 0.38 | 20.06 ± 4.94 * |
| DMDLS50 | 1.88 ± 0.31 ** | 5.19 ± 1.68 ** | 2.19 ± 0.51 | 0.83 ± 0.34 | 22.21 ± 4.39 ** |
| DMDLS100 | 1.90 ± 0.23 ** | 6.73 ± 0.71 | 2.20 ± 0.23 | 1.00 ± 0.34 | 17.55 ± 2.62 |
| Group | ALT | AST | UA | CRE |
|---|---|---|---|---|
| U/grot | μmol/L | |||
| Control | 21.33 ± 11.10 | 33.27 ± 14.82 | 230.23 ± 77.76 | 17.01 ± 7.67 |
| DLS200 | 28.21 ± 11.86 | 27.00 ± 11.88 | 305.51 ± 39.20 * | 20.38 ± 5.90 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.-W.; Huang, M.; Lo, K.; Chen, W.-L.; He, Y.-Y.; Xu, Y.; Zheng, H.; Hu, H.; Wang, J. Anti-Diabetic Effect of a Shihunine-Rich Extract of Dendrobium loddigesii on 3T3-L1 Cells and db/db Mice by Up-Regulating AMPK–GLUT4–PPARα. Molecules 2019, 24, 2673. https://doi.org/10.3390/molecules24142673
Li X-W, Huang M, Lo K, Chen W-L, He Y-Y, Xu Y, Zheng H, Hu H, Wang J. Anti-Diabetic Effect of a Shihunine-Rich Extract of Dendrobium loddigesii on 3T3-L1 Cells and db/db Mice by Up-Regulating AMPK–GLUT4–PPARα. Molecules. 2019; 24(14):2673. https://doi.org/10.3390/molecules24142673
Chicago/Turabian StyleLi, Xue-Wen, Meixiang Huang, Kakei Lo, Wei-Li Chen, Ying-Yan He, Yongli Xu, Huizhen Zheng, Haiyan Hu, and Jun Wang. 2019. "Anti-Diabetic Effect of a Shihunine-Rich Extract of Dendrobium loddigesii on 3T3-L1 Cells and db/db Mice by Up-Regulating AMPK–GLUT4–PPARα" Molecules 24, no. 14: 2673. https://doi.org/10.3390/molecules24142673
APA StyleLi, X. -W., Huang, M., Lo, K., Chen, W. -L., He, Y. -Y., Xu, Y., Zheng, H., Hu, H., & Wang, J. (2019). Anti-Diabetic Effect of a Shihunine-Rich Extract of Dendrobium loddigesii on 3T3-L1 Cells and db/db Mice by Up-Regulating AMPK–GLUT4–PPARα. Molecules, 24(14), 2673. https://doi.org/10.3390/molecules24142673