Methylation of the RELA Gene is Associated with Expression of NF-κB1 in Response to TNF-α in Breast Cancer


Abstract
:1. Introduction
2. Results
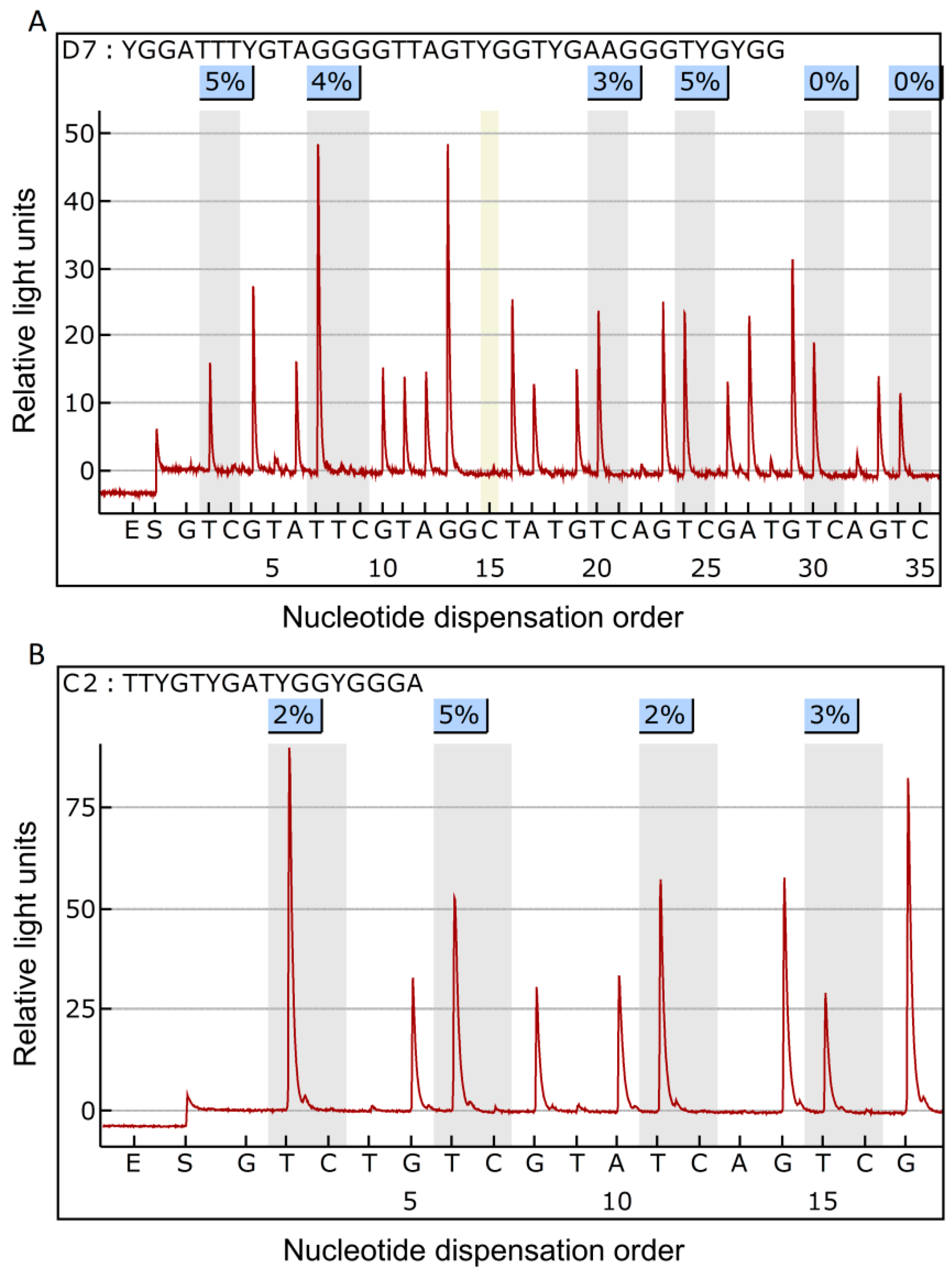
2.1. Methylation Status of the NF-κB-Related Genes in Breast Cancer
2.2. Association of Methylation Levels of the NF-κB-Related Genes and Gene Expression with Inflammatory Markers in Tumor Tissues
2.3. Association of Methylation Levels of the NF-κB-Related Genes with the Clinicopathologic Characteristics
3. Discussion
4. Materials and Methods
Author Contributions
Funding
Conflicts of Interest
References
- Gilmore, T.D. Introduction to NF-kappaB: Players, pathways, perspectives. Oncogene 2006, 25, 6680–6684. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, T.D.; Garbati, M.R. Inhibition of NF-κB Signaling as a Strategy in Disease Therapy. In NF-κB in Health and Disease. Curr Top Microbiol Immunol.; Karin, M., Ed.; Springer: Heidelberg, Germany, 2010; Volume 349, pp. 245–263. [Google Scholar]
- Arkan, M.C.; Greten, F.R. IKK- and NF-κB-Mediated Functions in Carcinogenesis. In NF-κB in Health and Disease. Curr Top Microbiol Immunol.; Karin, M., Ed.; Springer: Heidelberg, Germany, 2010; Volume 349, pp. 159–169. [Google Scholar]
- Prasad, S.; Ravindran, J.; Aqqarwal, B.B. NF-kappaB and cancer: How intimate is this relationship. Mol. Cell Biochem. 2010, 336, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Park, M.H.; Hong, J.T. Roles of NF-κB in cancer and inflammatory disease and their therapeutic approaches. Cells 2016, 5, 15. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Shen, S.; Verma, I.M. NF-κB, an active player in human cancers. Cancer Immunol. Res. 2014, 2, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Israel, A. The IKK complex, a central regulator of NF-kappaB activation. Cold Spring Harb. Perspect. Biol. 2010, 2, a000158. [Google Scholar] [CrossRef] [PubMed]
- Schmid, J.A.; Birbach, A. IkappaB kinase beta (IKKbeta/IKK2/IKBKB)—a key molecule in signaling to the transcription factor NF-kappaB. Cytokine Growth Factor Rev. 2008, 19, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Ea, C.K.; .Baltimore, D. Regulation of NF-kappaB activity through lysine monomethylation of p65. Proc. Natl. Acad. Sci. USA 2009, 106, 18972–19877. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.D.; Huang, B.; Li, M.; Lamb, A.; Kelleher, N.L.; Chen, L.F. Negative regulation of NF-kappaB action by Set9-mediated lysine methylation of the RELA subunit. Embo J. 2009, 28, 1055–1066. [Google Scholar] [CrossRef]
- Lu, T.; Yang, M.; Huang, D.B.; Wei, H.; Ozer, G.H.; Ghosh, G.; Stark, G.R. Role of lysine methylation of NF-κB in differential gene regulation. Proc. Natl. Acad. Sci. USA 2013, 110, 13510–13515. [Google Scholar] [CrossRef]
- Fernandez-Jimenez, N.; Castellanos-Rubio, A.; Plaza-Izurieta, L.; Irastorza, I.; Elcoroaristizabal, X.; Jauregi-Miguel, A.; Lopez-Euba, T.; Tutau, C.; de Pancorbo, M.M.; Vitoria, J.C.; et al. Coregulation and modulation of NFκB-Related genes in celiac disease: Uncovered aspects of gut mucosal inflammation. Hum. Mol. Genet. 2014, 23, 1298–1310. [Google Scholar] [CrossRef]
- Cao, Y.; Karin, M. NF-κB in mammary gland development and breast cancer. J. Mammary Gland Biol. Neoplasia 2003, 8, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Biswas, D.K.; Shi, Q.; Baily, S.; Strickland, I.; Ghosh, S.; Pardee, A.B.; Iglehart, J.D. NF-κB activation in human breast cancer specimens and its role in cell proliferation and apoptosis. Proc. Natl. Acad. Sci. USA 2004, 101, 10137–10142. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, N.; Ito, T.; Azuma, S.; Ito, E.; Honma, R.; Yanagisawa, Y.; Nishikawa, A.; Kawamura, M.; Imai, J.I.; Watanabe, S.; et al. Constitutive activation of nuclear factor-κB is preferentially involved in the proliferation of basal-like subtype breast cancer cell lines. Cancer Sci. 2009, 100, 1668–1674. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Ito, T.; Shimizu, T.; Ishida, T.; Semba, K.; Watanabe, S.; Yamaguchi, N.; Inoue, J.I. Epigenetic alteration of the NF-κB-inducing kinase (NIK) gene is involved in enhanced NIK expression in basal-like breast cancer. Cancer Sci. 2010, 101, 2391–2397. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Lyu, Y.L.; Cai, L. NF-κB affects proliferation and invasiveness of breast cancer cells by regulating CD44 expression. PLoS ONE 2014, 9, e106966. [Google Scholar] [CrossRef] [PubMed]
- Fusella, F.; Seclì, L.; Busso, E.; Krepelova, A.; Moiso, E.; Rocca, S.; Conti, L.; Annaratone, L.; Rubinetto, C.; Mello-Grand, M.; et al. The IKK/NF-κB signaling pathway requires Morgana to drive breast cancer metastasis. Nat. Commun. 2017, 8, 1636. [Google Scholar] [CrossRef] [PubMed]
- Kulis, M.; Esteller. DNA methylation and cancer. Adv. Genet. 2010, 70, 27–56. [Google Scholar]
- Yao, C.; Li, H.; Shen, X.; He, Z.; He, L.; Guo, Z. Reproducibility and concordance of differential DNA methylation and gene expression in cancer. PLoS ONE 2012, 7, e29686. [Google Scholar] [CrossRef]
- Braustein, S.; Formenti, S.C.; Schneider, R.J. Acquisition of stable inducible up-regulation of nuclear factor-kappaB by tumor necrosis factor exposure confers increased radiation resistance without increased transformation in breast cancer cells. Mol. Cancer Res. 2008, 6, 78–88. [Google Scholar] [CrossRef]
- Zhang, T.; Park, K.A.; Li, Y.; Byun, H.S.; Jeon, J.; Lee, Y.; Hong, J.H.; Kim, J.M.; Huang, S.M.; Choi, S.W.; et al. PHF20 regulates NF-κB signaling by disrupting recruitment of PP2A to p65. Nat. Commun. 2013, 4, 2062. [Google Scholar] [CrossRef]
- Schütze, S.; Wiegmann, K.; Machleidt, T.; Krönke, M. TNF-induced activation of NF-kappa B. Immnobiology 1995, 193, 193–203. [Google Scholar] [CrossRef]
- Acharyya, S.; Sharma, S.M.; Cheng, A.S.; Ladner, K.J.; He, W.; Klin, W.; Wang, H.; Ostrowski, M.C.; Huang, T.H.; Guttridge, D.C. TNF inhibits Notch-1 in skeletal muscle cells by Ezh2 and DNA methylation mediated repression: Implications in duchenne muscular dystrophy. PLoS ONE 2010, 5, e12479. [Google Scholar] [CrossRef] [PubMed]
- Morisawa, S.; Yasuda, H.; Kamiya, T.; Hara, H.; Adachi, T. Tumor necrosis factor-α decreases EC-SOD expression through DNA methylation. J. Clin. Biochem. Nutr. 2017, 60, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Slack, A.; Cervoni, N.; Pinard, M.; Szyf, M. Feedback regulation of DNA methyltransferase gene expression by methylation. Eur. J. Biochem. 1999, 264, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, C.; Kaneko, K. Epigenetic feedback regulation accelerates adaptation and evolution. PLoS ONE 2013, 8, e61251. [Google Scholar] [CrossRef] [PubMed]
- Akulenko, R.; Helms, V. DNA co-methylation analysis suggests novel functional associations between gene pairs in breast samples. Hum. Mol. Genet. 2013, 22, 3016–3022. [Google Scholar] [CrossRef]
- Jeong, Y.J.; Jeong, H.Y.; Bong, J.G.; Park, S.H.; Oh, H.K. Low methylation levels of the SFRP1 gene are associated with the basal-like subtype of breast cancer. Oncol. Rep. 2013, 29, 1946–1954. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |



{kind=link}
{kind=link}
| NFKB1 Methylation Level | RELA Methylation Level | ||||
|---|---|---|---|---|---|
| Mean (%) | p-Value | Mean (%) | p-Value | ||
| NF-κB p50 transcript | Negative | 1.31 ± 0.87 | 0.717 | 1.33 ± 0.64 | 0.182 |
| Positive | 1.41 ± 0.99 | 1.55 ± 0.48 | |||
| NFκB p50 transcript in TNF-α(+) tumor | Negative | 1.42 ± 1.27 | 0.691 | 1.17 ± 0.53 | 0.002 * |
| Positive | 1.27 ± 0.85 | 1.71 ± 0.39 | |||
| Inflammatory Markers | NFKB1 Methylation Level | RELA Methylation Level | |||
|---|---|---|---|---|---|
| Mean Level (%) | p-Value | Mean Level (%) | p-Value | ||
| TNF-α, n = 46 | Negative | 1.56 ± 0.92 | 0.055 | 1.18 ± 0.52 | 0.009 * |
| Positive | 1.06 ± 0.80 | 1.71 ± 0.39 | |||
| IL-4, n = 47 | Negative | 1.17 ± 1.02 | 0.082 | 1.47 ± 0.52 | 0.757 |
| Positive | 1.60 ± 0.73 | 1.51 ± 0.52 | |||
| Intratumoral inflammation, n = 47 | Negative | 1.43 ± 1.03 | 0.350 | 1.40 ± 0.56 | 0.107 |
| Positive | 1.19 ± 0.61 | 1.70 ± 0.44 | |||
| Peritumoral inflammation, n = 36 | Negative | 0.97 ± 0.87 | 0.139 | 1.32 ± 0.58 | 0.460 |
| Positive | 1.55 ± 0.97 | 1.49 ± 0.58 | |||
| Inflammatory Markers | NFκB p50 Transcript | |||
|---|---|---|---|---|
| Negative (%) | Positive (%) | p-Value | ||
| TNF-α, n = 47 | Negative | 55.2 | 44.8 | 0.001 * |
| Positive | 5.6 | 94.4 | ||
| IL-4, n =47 | Negative | 54.5 | 45.5 | 0.014 * |
| Positive | 20.0 | 80.0 | ||
| Intratumoral inflammation, n = 47 | Negative | 33.3 | 66.7 | 0.493 |
| Positive | 45.5 | 54.5 | ||
| Peritumoral inflammation, n = 36 | Negative | 25.0 | 75.0 | 0.682 |
| Positive | 39.3 | 60.7 | ||
| Clinicopathologic Variables | NFKB1 Methylation Level | RELA Methylation Level | |||
|---|---|---|---|---|---|
| Mean Level (%) | p-Value | Mean Level (%) | p-Value | ||
| Age (years) | ≤50 | 1.70 ± 0.77 | 0.062 | 1.47 ± 0.50 | 0.960 |
| >50 | 1.17 ± 1.00 | 1.47 ± 0.58 | |||
| Menopausal status | Premenopause | 0.84 ± 0.98 | 0.208 | 1.63 ± 0.17 | 0.514 |
| Postmenopause | 1.49 ± 0.96 | 1.43 ± 0.60 | |||
| Tumor size (cm) | ≤2 | 1.30 ± 0.89 | 0.505 | 1.41 ± 0.53 | 0.405 |
| >2 | 1.49 ± 1.03 | 1.55 ± 0.57 | |||
| Node Metastasis | Negative | 1.30 ± 0.92 | 0.463 | 1.46 ± 0.58 | 0.830 |
| Positive | 1.51 ± 0.99 | 1.49 ± 0.51 | |||
| Histologic grade | I | 1.78 ± 0.91 | 0.386 | 1.28 ± 0.17 | 0.255 |
| II | 1.20 ± 1.33 | 1.67 ± 0.34 | |||
| III | 1.33 ± 0.72 | 1.47 ± 0.55 | |||
| Stage | I | 1.35 ± 0.98 | 0.982 | 1.42 ± 0.48 | 0.598 |
| II | 1.39 ± 0.95 | 1.57 ± 0.52 | |||
| III | 1.30 ± 1.24 | 1.34 ± 1.18 | |||
| Molecular subtype | Luminal A | 0.93 ± 0.79 | 0.392 | 1.47 ± 0.61 | 0.371 |
| Luminal B | 1.55 ± 1.04 | 1.48 ± 0.54 | |||
| HER2 | 1.376 ± 1.22 | 1.29 ± 0.58 | |||
| Basal-like | 1.80 ± 0.35 | 2.00 ± 0.69 | |||
| Positive | 1.33 ± 1.00 | 1.51 ± 0.56 | |||
| ER status | Negative | 1.45 ± 0.78 | 0.723 | 1.54 ± 0.53 | 0.723 |
| Positive | 1.34 ± 1.02 | 1.44 ± 0.56 | |||
| PR status | Negative | 1.53 ± 0.95 | 0.573 | 1.54 ± 0.64 | 0.650 |
| Positive | 1.33 ± 0.95 | 1.45 ± 0.53 | |||
| HER2 overexpression | Negative | 1.20 ± 0.88 | 0.155 | 1.58 ± 0.60 | 0.238 |
| Positive | 1.65 ± 1.08 | 1.67 ± 0.52 | |||
| Ki-67 | <14% | 1.27 ± 0.69 | 0.761 | 1.65 ± 0.27 | 0.351 |
| ≥14% | 1.39 ± 0.99 | 1.44 ± 0.58 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, Y.J.; Oh, H.K.; Choi, H.R. Methylation of the RELA Gene is Associated with Expression of NF-κB1 in Response to TNF-α in Breast Cancer. Molecules 2019, 24, 2834. https://doi.org/10.3390/molecules24152834
Jeong YJ, Oh HK, Choi HR. Methylation of the RELA Gene is Associated with Expression of NF-κB1 in Response to TNF-α in Breast Cancer. Molecules. 2019; 24(15):2834. https://doi.org/10.3390/molecules24152834
Chicago/Turabian StyleJeong, Young Ju, Hoon Kyu Oh, and Hye Ryeon Choi. 2019. "Methylation of the RELA Gene is Associated with Expression of NF-κB1 in Response to TNF-α in Breast Cancer" Molecules 24, no. 15: 2834. https://doi.org/10.3390/molecules24152834
APA StyleJeong, Y. J., Oh, H. K., & Choi, H. R. (2019). Methylation of the RELA Gene is Associated with Expression of NF-κB1 in Response to TNF-α in Breast Cancer. Molecules, 24(15), 2834. https://doi.org/10.3390/molecules24152834