Evaluating the Bioactivity of a Novel Antimicrobial and Anticancer Peptide, Dermaseptin-PS4(Der-PS4), from the Skin Secretion of Phyllomedusa sauvagii

 , and
, and 
Abstract
:1. Introduction
2. Results
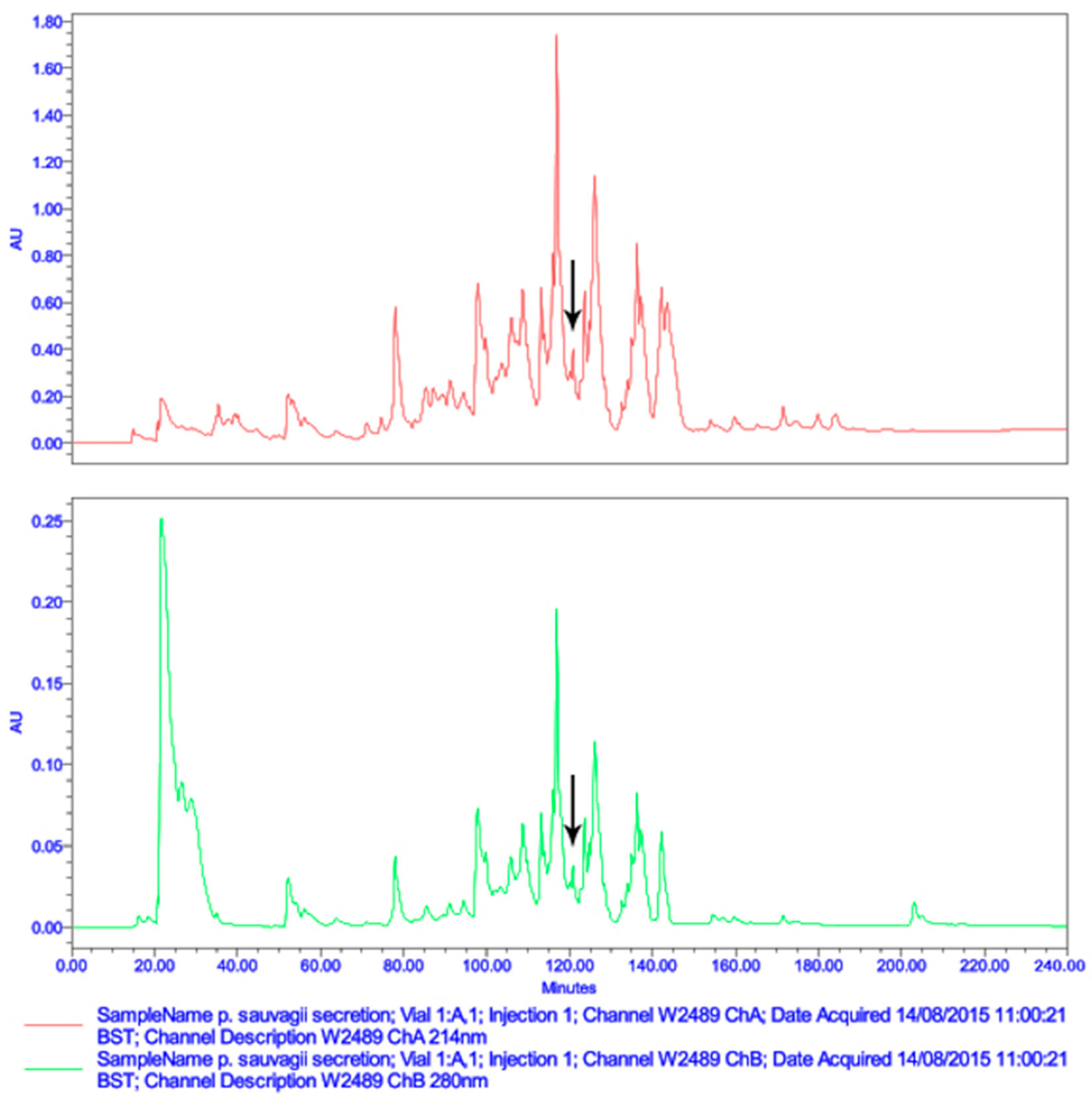
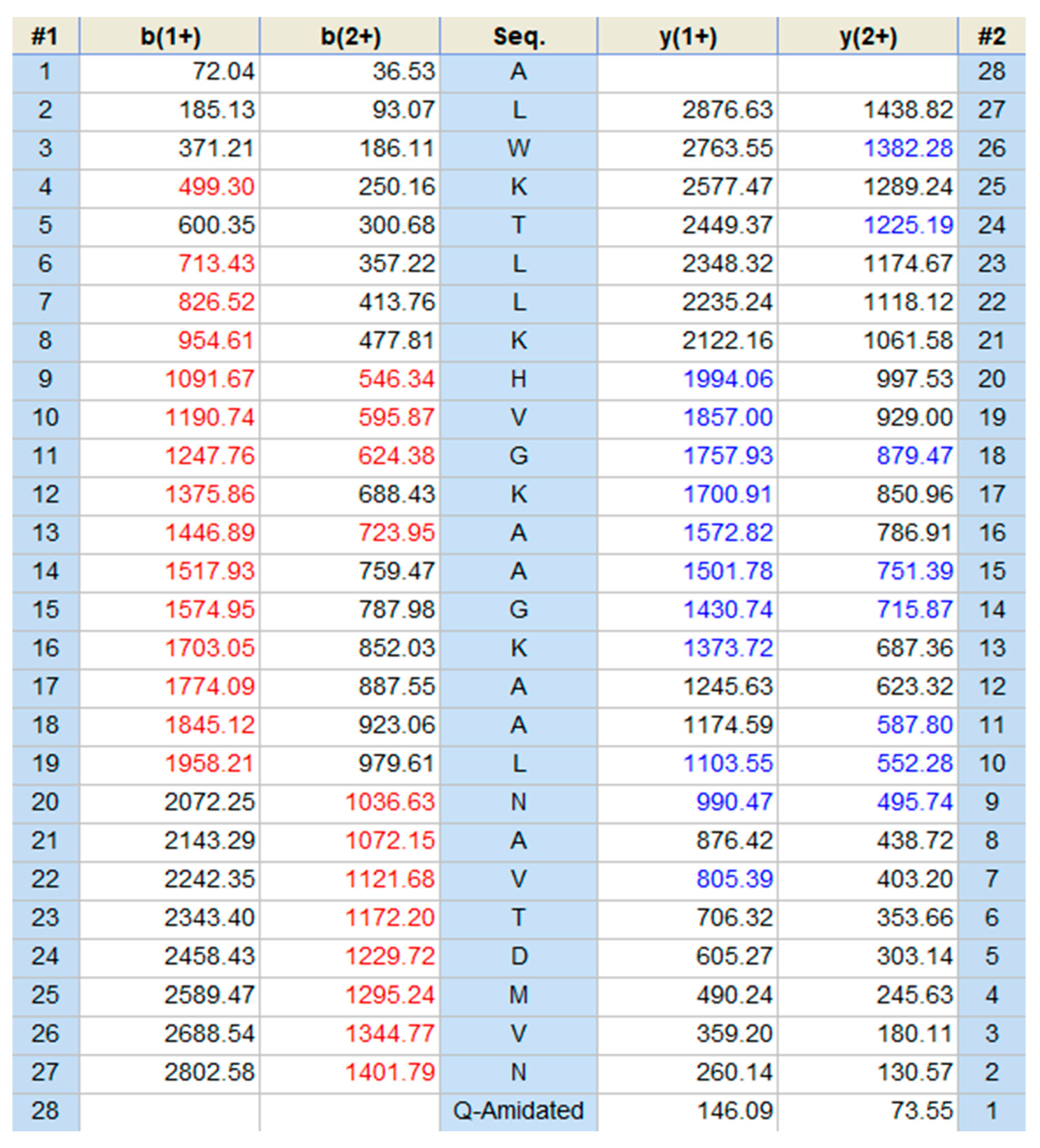
2.1. Molecular Cloning and Structural Characterisation of Der-PS4 from a Skin Secretion-Derived cDNA Library of Phyllomedusa sauvagii
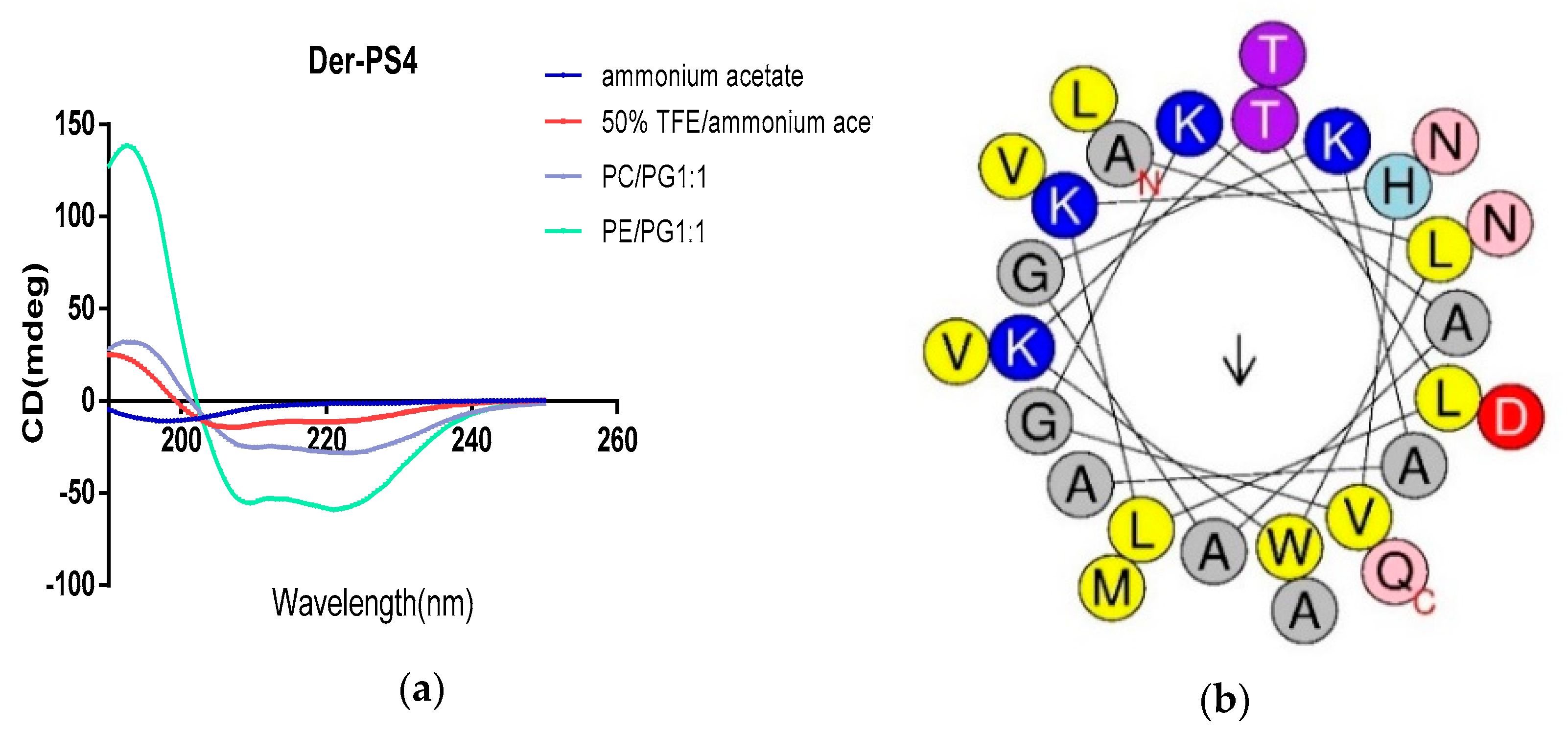
2.2. Secondary Structure Prediction and Structrual Parameters Analysis of Der-PS4
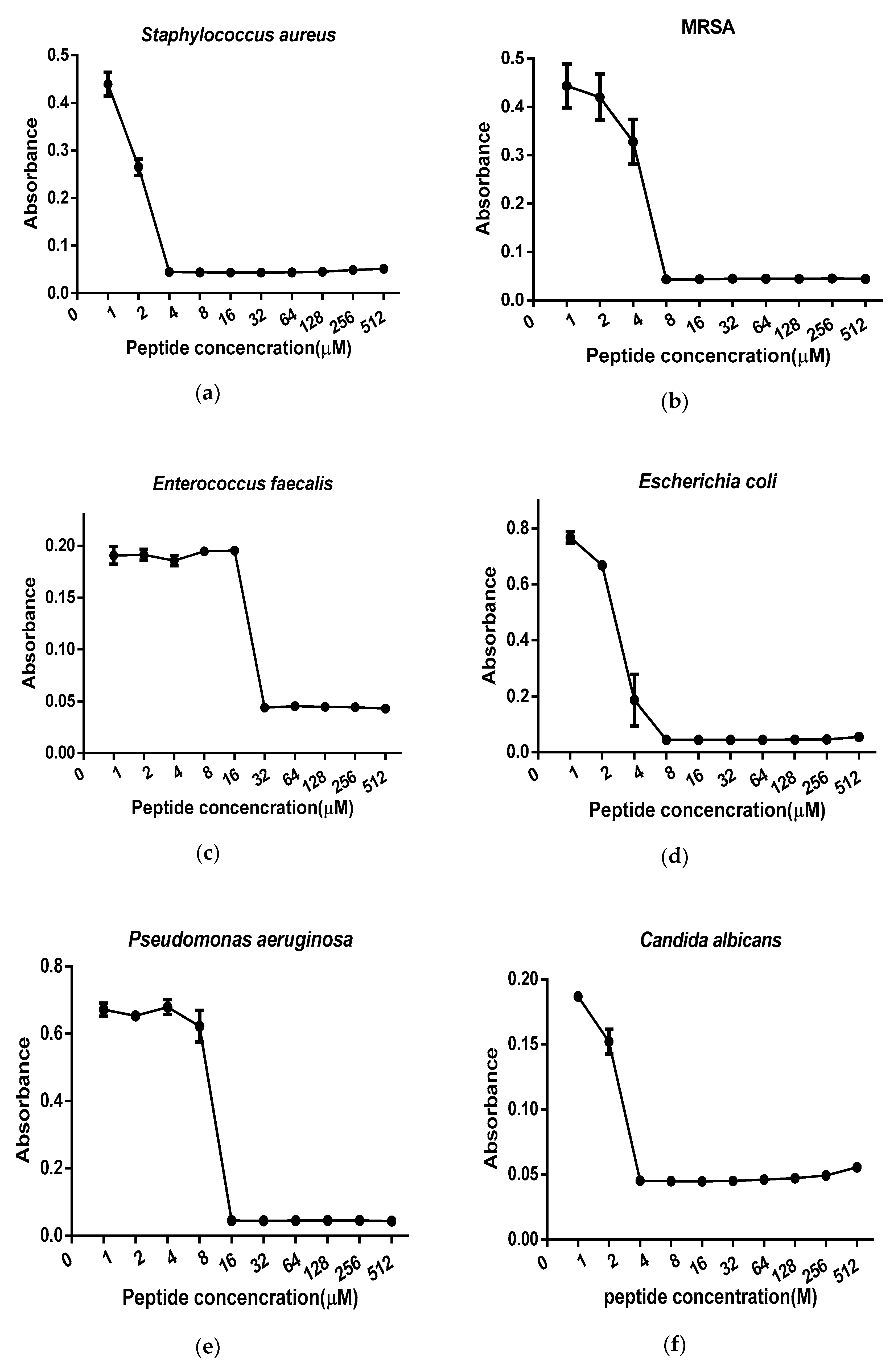
2.3. Antimicrobial Minimal Inhibitory Concentration Assay
2.4. Anti-Biofilm Activity of Der-PS4
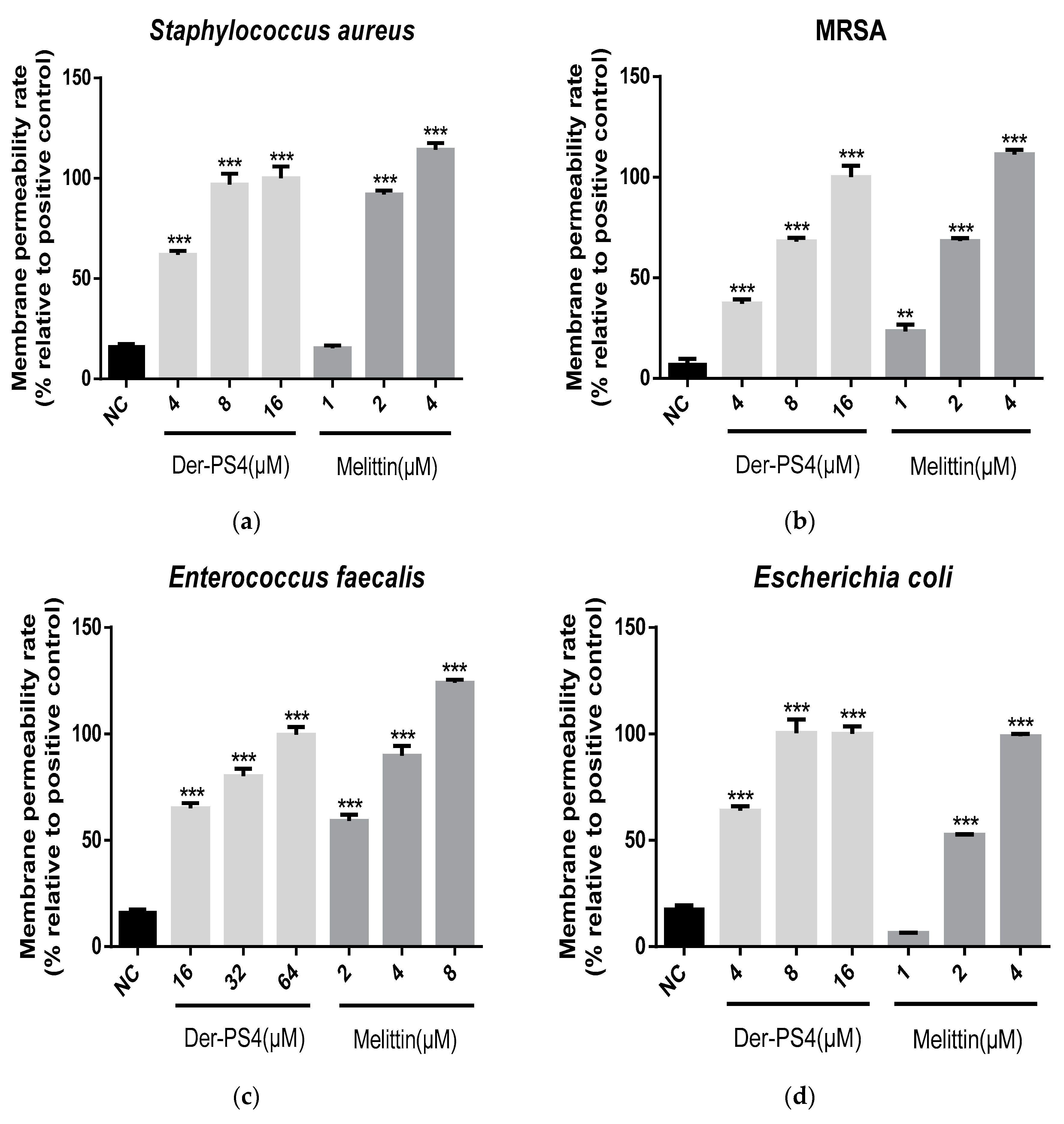
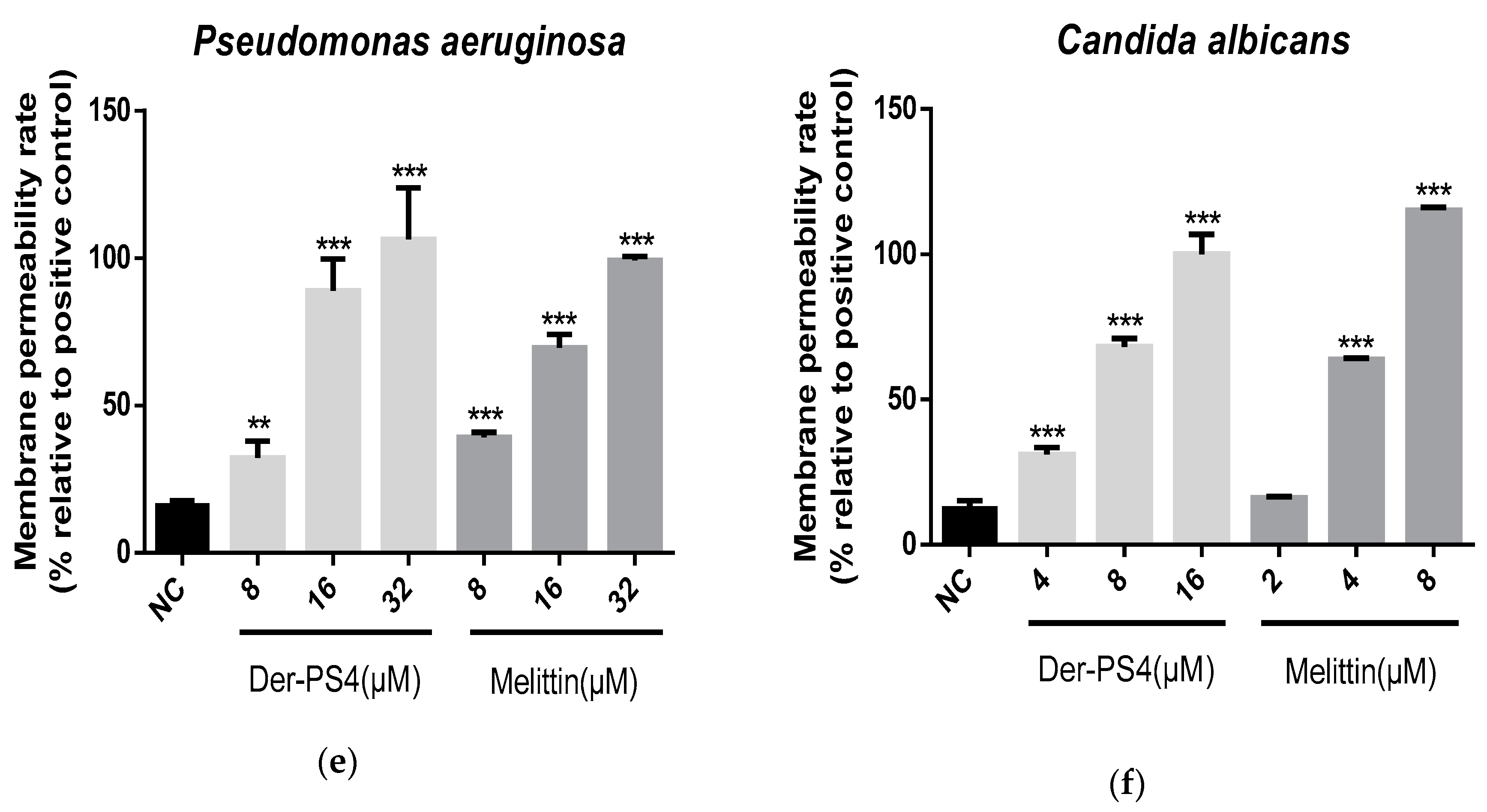
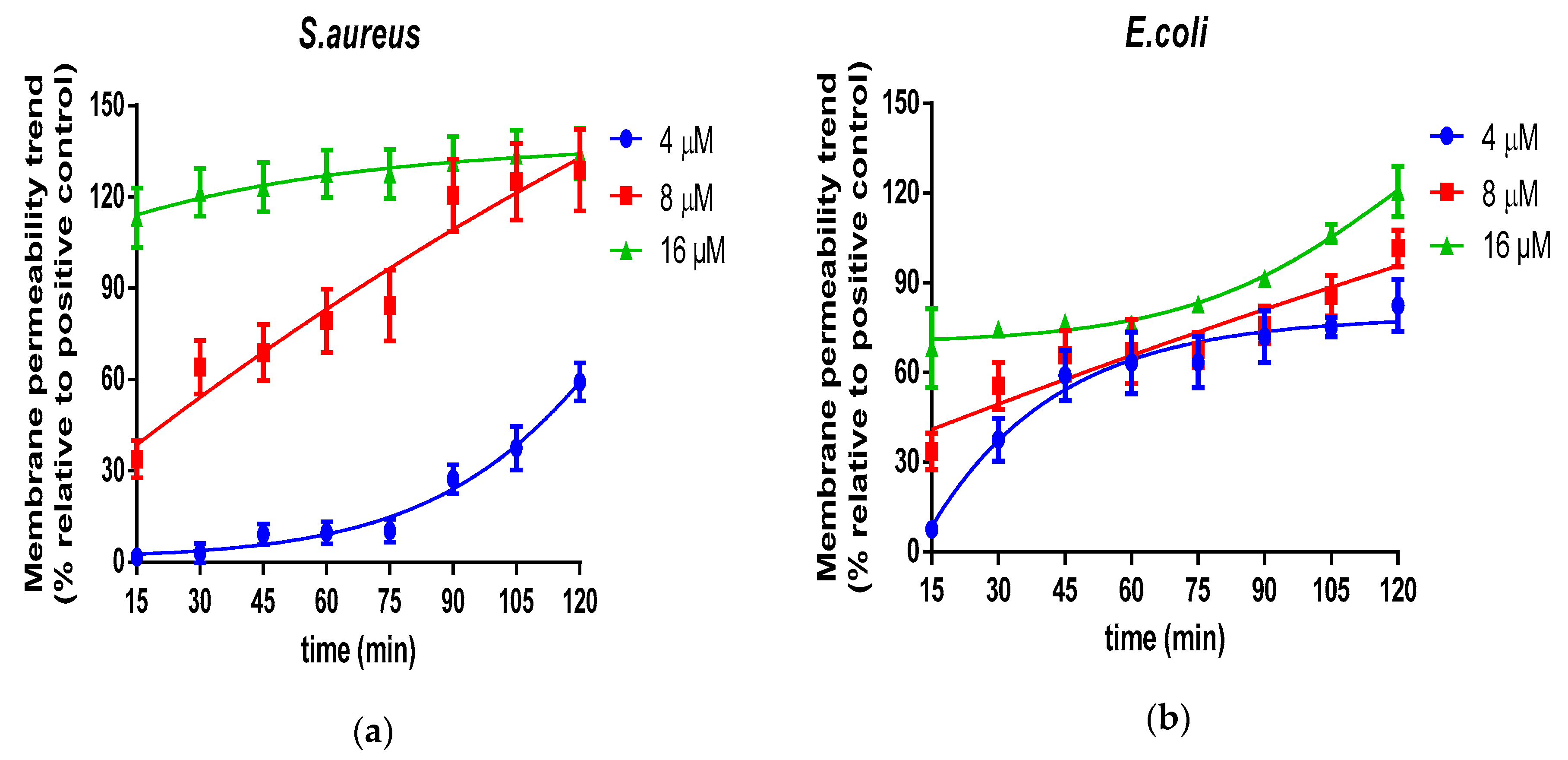
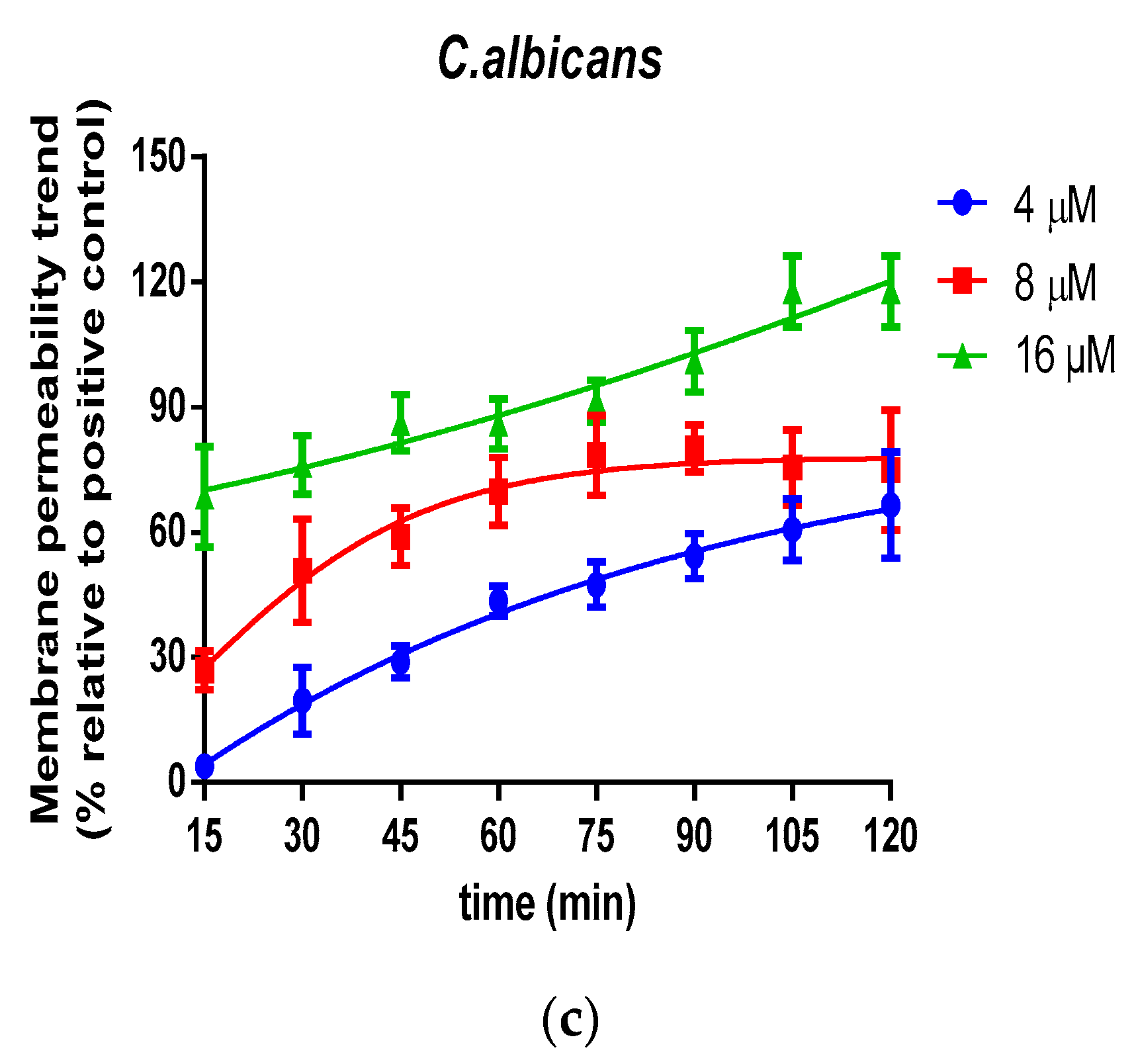
2.5. Bacteria Cell Membrane Permeabilized Activity of Der-PS4
2.6. Scanning Electron Microscopy (SEM) Characterisation
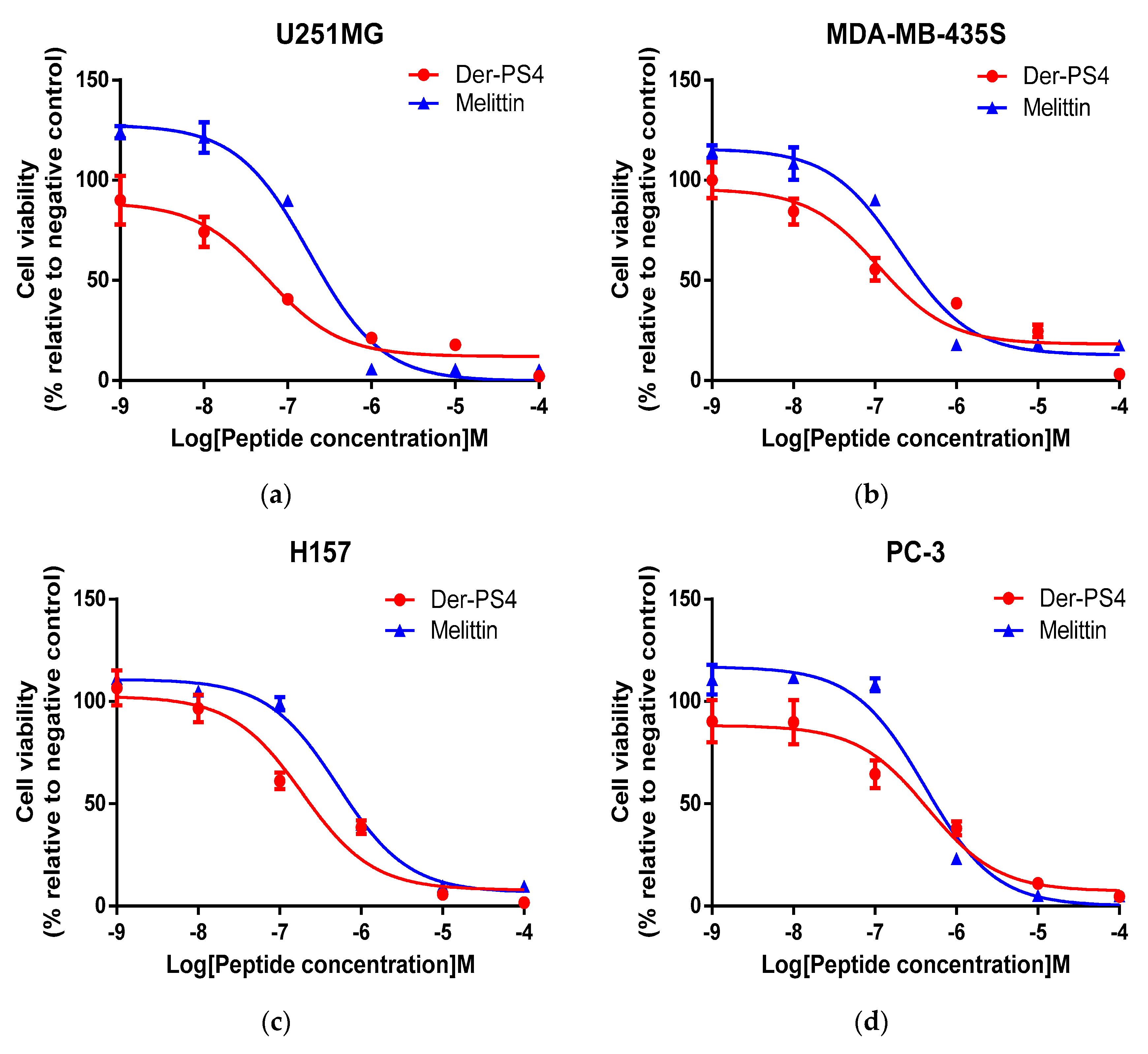
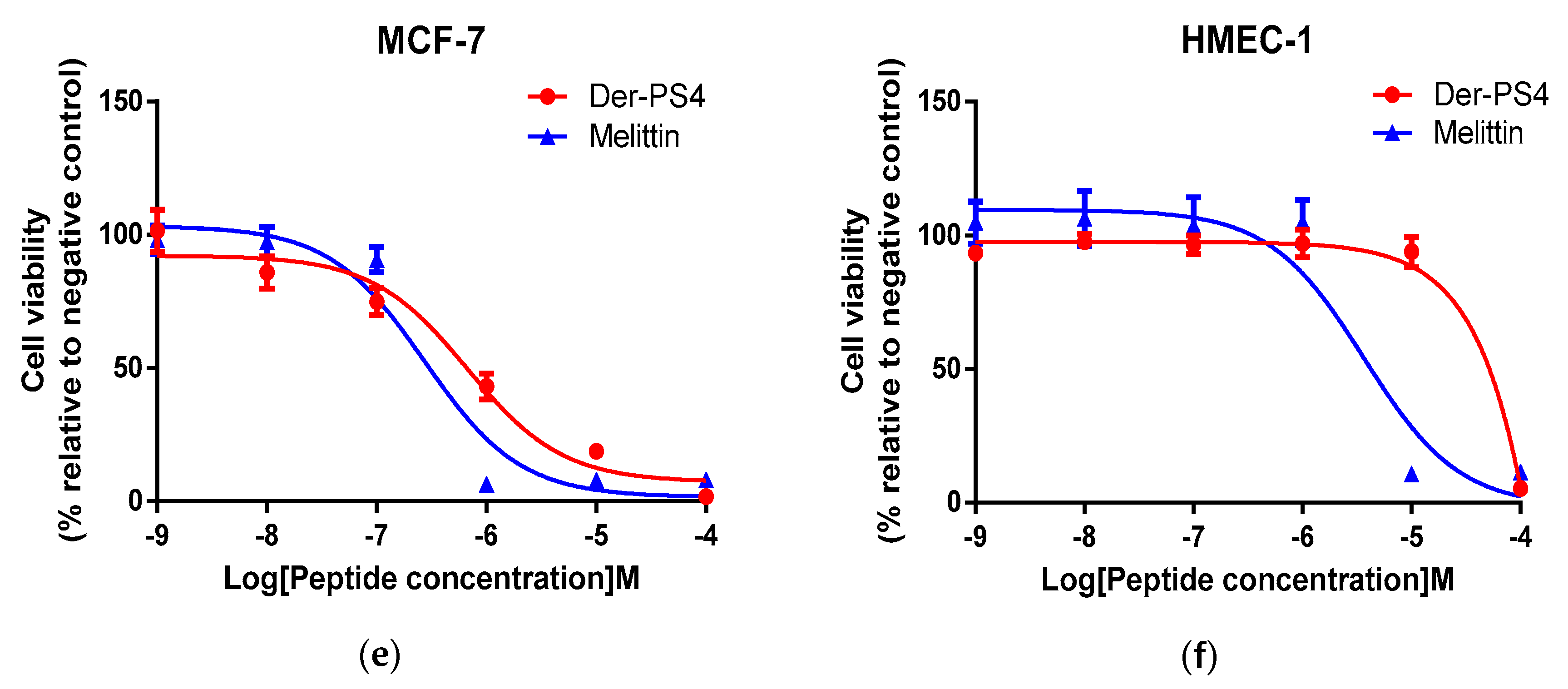
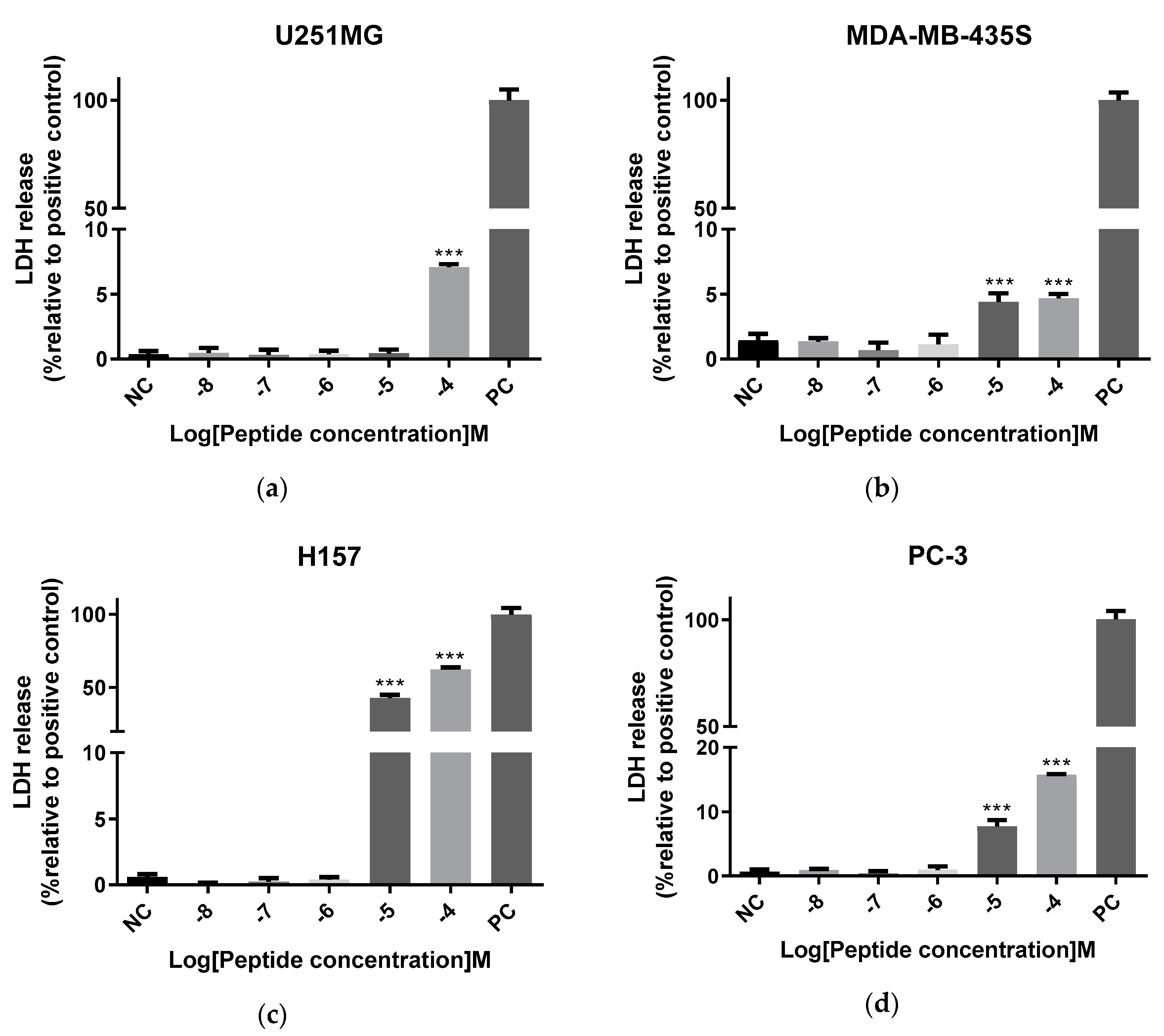
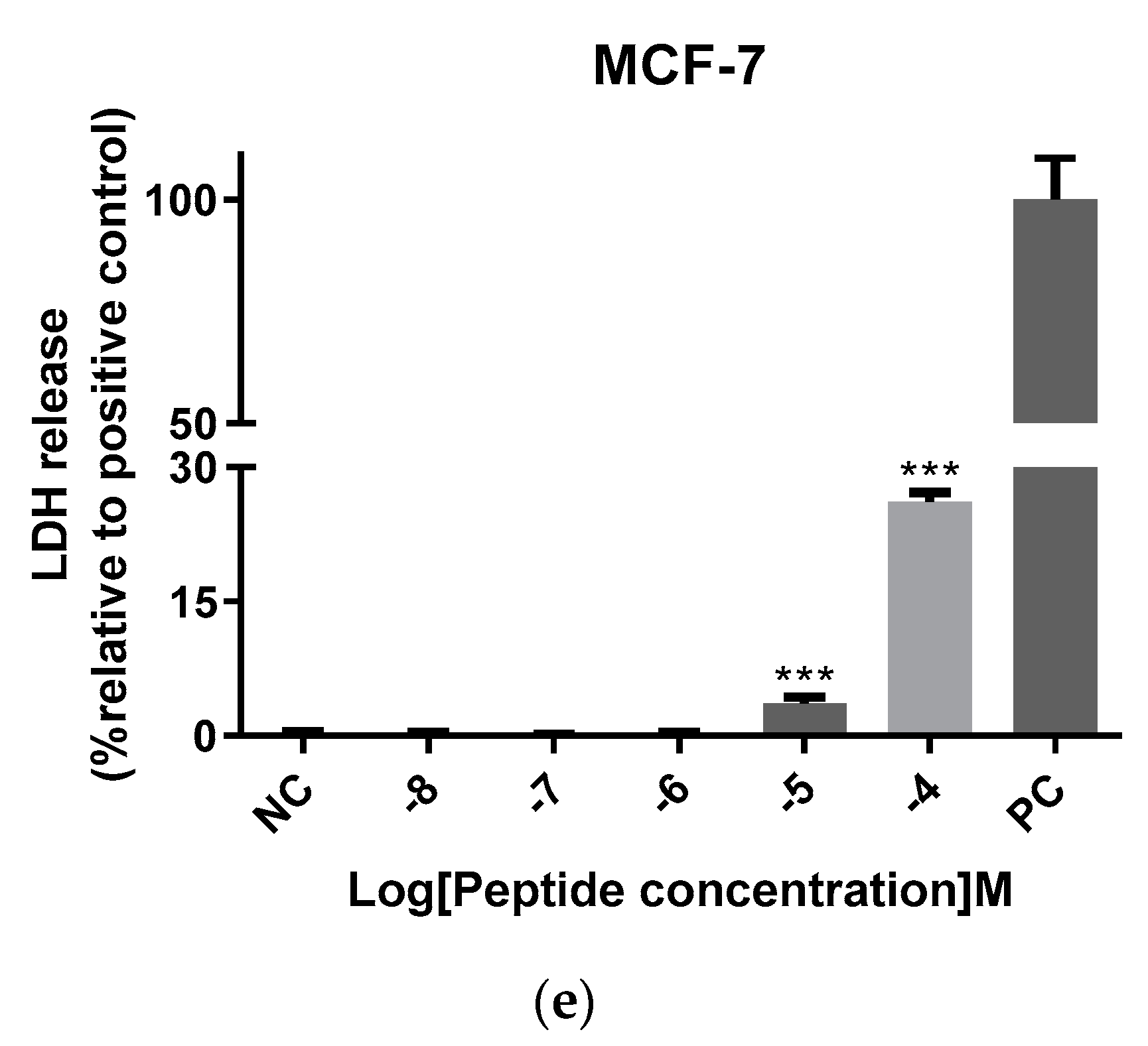
2.7. Antiproliferative Activity of Der-PS4
3. Discussion
4. Materials and Methods
4.1. Acquisition of Phyllomedusa sauvagii Skin Secretions
4.2. Molecular Cloning and Structural Characterisation of Der-PS4 from a Skin Secretion-Derived cDNA Library of Phyllomedusa sauvagii
4.3. Identification and Structural Chracterisation of the Predicted Der-PS4 from the Skin Secretion of Phyllomedusa sauvagii
4.4. Solid-Phase Synthesis of Der-PS4
4.5. Secondary Structural Analysis of Der-PS4 through Circular Dichroism (CD) Spectroscopy
4.6. Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) Assays
4.7. Minimum Biofilm Inhibitory Concentration (MBIC) and Minimum Biofilm Eradication Concentration (MBEC) Assays
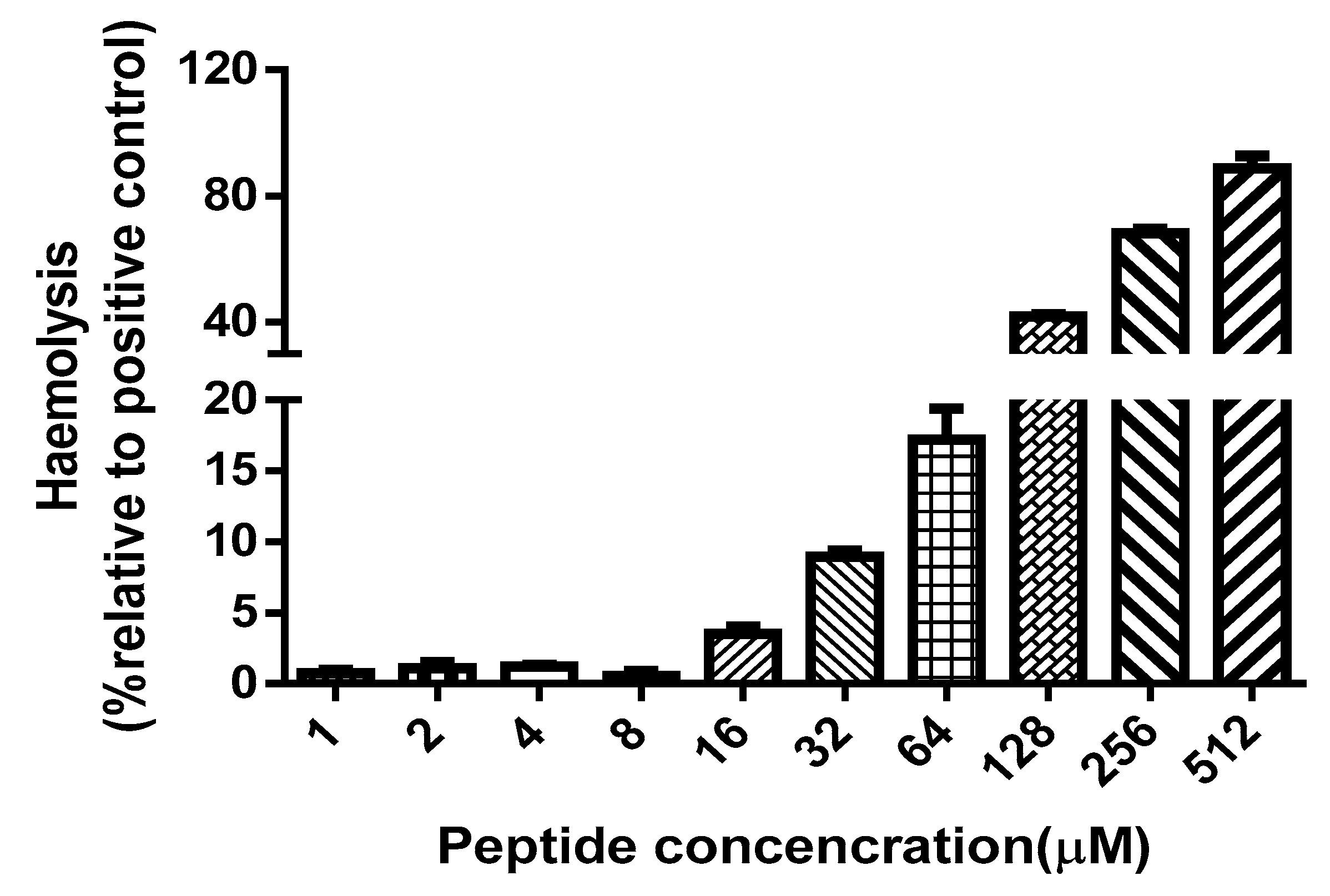
4.8. Haemollysis Assay
4.9. Bacteria Cell Membrane Permeability Assay
4.10. Scanning Electron Microscope (SEM) Characterisation
4.11. MTT Cell Viability Assay
4.12. Lactate Dehydrogenase (LDH) Cytotoxicity Assay
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| RP-HPLC | reverse-phase high-performance liquid chromatography |
| SPPS | solid-phase peptide synthesis |
| CD | circular dichroism |
| TFE | 9-fluorenylmethyloxycarbonyl |
| SEM | scanning electron microscopy |
| MTT | 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide |
| LDH | lactate dehydrogenase |
| RACE-PCR | rapid amplification of cDNA ends-polymerase chain reaction |
| NUP | nested universal primer |
| TFA | trifluoracetic acid |
| MALDI-TOF | matrix-assisted laser desorption/ionization time-of-flight |
| CHCA | α-cyano-4-hydroxycinnamic acid |
| Fmoc | 9-fluorenylmethyloxycarbonyl |
| HBTU | 2-(1H-benzotriazol-1-yl)-1,1,3,3-tetramethyluronium hexafluorophosphate |
| OD | optical density |
References
- Conlon, J.M.; Mechkarska, M.; Lukic, M.L.; Flatt, P.R. Potential therapeutic applications of multifunctional host-defense peptides from frog skin as anti-cancer, anti-viral, immunomodulatory, and anti-diabetic agents. Peptides 2014, 57, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389. [Google Scholar] [CrossRef] [PubMed]
- Simmaco, M.; Mignogna, G.; Barra, D. Antimicrobial peptides from amphibian skin: What do they tell us? Pept. Sci. 1998, 47, 435–450. [Google Scholar] [CrossRef]
- Conlon, J.M. The contribution of skin antimicrobial peptides to the system of innate immunity in anurans. Cell Tissue Res. 2011, 343, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Woodhams, D.C.; Vredenburg, V.T.; Simon, M.-A.; Billheimer, D.; Shakhtour, B.; Shyr, Y.; Briggs, C.J.; Rollins-Smith, L.A.; Harris, R.N. Symbiotic bacteria contribute to innate immune defenses of the threatened mountain yellow-legged frog, Rana muscosa. Biol. Conserv. 2007, 138, 390–398. [Google Scholar] [CrossRef]
- Shai, Y. Mechanism of the binding, insertion and destabilization of phospholipid bilayer membranes by α-helical antimicrobial and cell non-selective membrane-lytic peptides. Biochim. Biophys. Acta Biomembr. 1999, 1462, 55–70. [Google Scholar] [CrossRef]
- Tennessen, J.A. Molecular evolution of animal antimicrobial peptides: Widespread moderate positive selection. J. Evol. Biol. 2005, 18, 1387–1394. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.; Diamond, G. The role of cationic antimicrobial peptides in innate host defences. Trends Microbiol. 2000, 8, 402–410. [Google Scholar] [CrossRef]
- Nicolas, P.; El Amri, C. The dermaseptin superfamily: A gene-based combinatorial library of antimicrobial peptides. Biochim. Biophys. Acta. 2009, 1788, 1537–1550. [Google Scholar] [CrossRef] [Green Version]
- Mor, A.; Van Huong, N.; Delfour, A.; Migliore-Samour, D.; Nicolas, P. Isolation, amino acid sequence and synthesis of dermaseptin, a novel antimicrobial peptide of amphibian skin. Biochemistry 1991, 30, 8824–8830. [Google Scholar] [CrossRef]
- de Kroon, A.I.; Dolis, D.; Mayer, A.; Lill, R.; de Kruijff, B. Phospholipid composition of highly purified mitochondrial outer membranes of rat liver and Neurospora crassa. Is cardiolipin present in the mitochondrial outer membrane? Biochim. Biophys. Acta Biomembr. 1997, 1325, 108–116. [Google Scholar] [CrossRef] [Green Version]
- Daly, J.W.; Caceres, J.; Moni, R.W.; Gusovsky, F.; Moos, M.; Seamon, K.B.; Milton, K.; Myers, C.W. Frog secretions and hunting magic in the upper Amazon: Identification of a peptide that interacts with an adenosine receptor. Proc. Natl. Acad. Sci. USA 1992, 89, 10960–10963. [Google Scholar] [CrossRef] [PubMed]
- Mor, A.; Hani, K.; Nicolas, P. The vertebrate peptide antibiotics dermaseptins have overlapping structural features but target specific microorganisms. J. Biol. Chem. 1994, 269, 31635–31641. [Google Scholar] [PubMed]
- Amiche, M.; Ducancel, F.; Lajeunesse, E.; Boulain, J.; Menez, A.; Nicolas, P. Molecular cloning of a cDNA encoding the precursor of adenoregulin from frog skin: Relationships with the vertebrate defensive peptides, dermaseptins. Biochem. Biophys. Res. Commun. 1993, 191, 983–990. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Chen, D.; Wang, L.; Lin, C.; Ma, C.; Xi, X.; Chen, T.; Shaw, C.; Zhou, M. Dermaseptin-PH: A novel peptide with antimicrobial and anticancer activities from the skin secretion of the south American orange-legged leaf frog, Pithecopus (Phyllomedusa) hypochondrialis. Molecules 2017, 22, 1805. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Chen, X.; Ma, C.; Xi, X.; Wang, L.; Zhou, M.; Burrows, J.F.; Kwok, H.F.; Chen, T. Biological Activities of Cationicity-Enhanced and Hydrophobicity-Optimized Analogues of an Antimicrobial Peptide, Dermaseptin-PS3, from the Skin Secretion of Phyllomedusa sauvagii. Toxins 2018, 10, 320. [Google Scholar] [CrossRef]
- Uniprot. Available online: https://www.uniprot.org/ (accessed on 28 December 2018).
- Shi, D.; Hou, X.; Wang, L.; Gao, Y.; Wu, D.; Xi, X.; Zhou, M.; Kwok, H.F.; Duan, J.; Chen, T.; et al. Two novel dermaseptin-like antimicrobial peptides with anticancer activities from the skin secretion of Pachymedusa dacnicolor. Toxins 2016, 8, 144. [Google Scholar] [CrossRef]
- Gao, Y.; Wu, D.; Xi, X.; Wu, Y.; Ma, C.; Zhou, M.; Wang, L.; Yang, M.; Chen, T.; Shaw, C. Identification and characterisation of the antimicrobial peptide, phylloseptin-PT, from the skin secretion of Phyllomedusa tarsius, and comparison of activity with designed, cationicity-enhanced analogues and diastereomers. Molecules 2016, 21, 1667. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.; Carmeli, Y.; Falagas, M.; Giske, C.; Harbarth, S.; Hindler, J.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Falagas, M.E.; Bliziotis, I.A. Pandrug-resistant Gram-negative bacteria: The dawn of the post-antibiotic era? Int. J. Antimicrob. Agents 2007, 29, 630–636. [Google Scholar] [CrossRef]
- Lara, H.H.; Ayala-Núñez, N.V.; Ixtepan Turrent, L.D.C.; Rodríguez Padilla, C. Bactericidal effect of silver nanoparticles against multidrug-resistant bacteria. World J. Microb. Biotechnol. 2009, 26, 615–621. [Google Scholar] [CrossRef]
- Tiwari, H.K.; Sapkota, D.; Sen, M.R. High prevalence of multidrug-resistant MRSA in a tertiary care hospital of northern India. Infect. Drug Resist. 2008, 1, 57. [Google Scholar] [CrossRef] [PubMed]
- Beveridge, T.J. Structures of gram-negative cell walls and their derived membrane vesicles. J. Bacteriol. 1999, 181, 4725–4733. [Google Scholar] [PubMed]
- Navarre, W.W.; Schneewind, O. Surface proteins of gram-positive bacteria and mechanisms of their targeting to the cell wall envelope. Microbiol. Mol. Biol. Rev. 1999, 63, 174–229. [Google Scholar] [PubMed]
- Lima, K.C.; Fava, L.R.; Siqueira, J.F., Jr. Susceptibilities of Enterococcus faecalis biofilms to some antimicrobial medications. J. Endod. 2001, 27, 616–619. [Google Scholar] [CrossRef] [PubMed]
- Strateva, T.; Yordanov, D. Pseudomonas aeruginosa—A phenomenon of bacterial resistance. J. Med. Microbiol. 2009, 58, 1133–1148. [Google Scholar] [CrossRef]
- Furiga, A.; Lajoie, B.; El Hage, S.; Baziard, G.; Roques, C. Impairment of Pseudomonas aeruginosa biofilm resistance to antibiotics by combining the drugs with a new quorum-sensing Inhibitor. Antimicrob. Agents Chemother. 2015, 60, 1676–1686. [Google Scholar] [CrossRef]
- Breidenstein, E.B.; de la Fuente-Núñez, C.; Hancock, R.E. Pseudomonas aeruginosa: All roads lead to resistance. Trends Microbiol. 2011, 19, 419–426. [Google Scholar] [CrossRef]
- O’Toole, G.; Kaplan, H.B.; Kolter, R. Biofilm formation as microbial development. Annu. Rev. Microbiol. 2000, 54, 49–79. [Google Scholar] [CrossRef]
- Rasamiravaka, T.; Labtani, Q.; Duez, P.; El Jaziri, M. The formation of biofilms by Pseudomonas aeruginosa: A review of the natural and synthetic compounds interfering with control mechanisms. Biomed. Res. Int. 2015, 2015, 759348. [Google Scholar] [CrossRef]
- Blackledge, M.S.; Worthington, R.J.; Melander, C. Biologically inspired strategies for combating bacterial biofilms. Curr. Opin. Pharmacol. 2013, 13, 699–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Liu, Y.; Wu, H.; Song, Z.; Hoiby, N.; Molin, S.; Givskov, M. Combating biofilms. FEMS Immunol. Med. Microbiol. 2012, 65, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Masak, J.; Cejkova, A.; Schreiberova, O.; Rezanka, T. Pseudomonas biofilms: Possibilities of their control. FEMS Microbiol. Ecol. 2014, 89, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, N.W.; Wong, G.C. Antimicrobial peptides and induced membrane curvature: Geometry, coordination chemistry, and molecular engineering. Curr. Opin. Solid State Mater. Sci. 2013, 17, 151–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galanth, C.; Abbassi, F.; Lequin, O.; Ayala-Sanmartin, J.; Ladram, A.; Nicolas, P.; Amiche, M. Mechanism of antibacterial action of dermaseptin B2: Interplay between helix− hinge− helix structure and membrane curvature strain. Biochemistry 2008, 48, 313–327. [Google Scholar] [CrossRef] [PubMed]
- Hoskin, D.W.; Ramamoorthy, A. Studies on anticancer activities of antimicrobial peptides. Biochim. Biophys. Acta Biomembr. 2008, 1778, 357–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Utsugi, T.; Schroit, A.J.; Connor, J.; Bucana, C.D.; Fidler, I.J. Elevated expression of phosphatidylserine in the outer membrane leaflet of human tumor cells and recognition by activated human blood monocytes. Cancer Res. 1991, 51, 3062–3066. [Google Scholar] [PubMed]
- Dobrzyńska, I.; Szachowicz-Petelska, B.; Sulkowski, S.; Figaszewski, Z. Changes in electric charge and phospholipids composition in human colorectal cancer cells. Mol. Cell. Biochem. 2005, 276, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Yoon, W.-H.; Park, H.-D.; Lim, K.; Hwang, B.-D. Effect of O-glycosylated mucin on invasion and metastasis of HM7 human colon cancer cells. Biochem. Biophys. Res. Commun. 1996, 222, 694–699. [Google Scholar] [CrossRef] [PubMed]
- Burdick, M.D.; Harris, A.; Reid, C.J.; Iwamura, T.; Hollingsworth, M.A. Oligosaccharides expressed on MUC1 produced by pancreatic and colon tumor cell lines. J. Biol. Chem. 1997, 272, 24198–24202. [Google Scholar] [CrossRef]
- Long, Q.; Li, L.; Wang, H.; Li, M.; Wang, L.; Zhou, M.; Su, Q.; Chen, T.; Wu, Y. Novel peptide dermaseptin-PS1 exhibits anticancer activity via induction of intrinsic apoptosis signalling. J. Cell. Mol. Med. 2018, 23, 1300–1312. [Google Scholar] [CrossRef] [PubMed]
- Tyler, M.J.; Stone, D.J.; Bowie, J.H. A novel method for the release and collection of dermal, glandular secretions from the skin of frogs. J. Pharmacol. Toxicol. Methods 1992, 28, 199–200. [Google Scholar] [CrossRef]
- Zhou, X.; Shi, D.; Zhong, R.; Ye, Z.; Ma, C.; Zhou, M.; Xi, X.; Wang, L.; Chen, T.; Kwok, H.F. Bioevaluation of Ranatuerin-2Pb from the Frog Skin Secretion of Rana pipiens and its Truncated Analogues. Biomolecules 2019, 9, 249. [Google Scholar] [CrossRef] [PubMed]
- Micsonai, A.; Wien, F.; Kernya, L.; Lee, Y.H.; Goto, Y.; Refregiers, M.; Kardos, J. Accurate secondary structure prediction and fold recognition for circular dichroism spectroscopy. Proc. Natl. Acad. Sci. USA 2015, 112, E3095–E3103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, G.; Balamurugan, P.; Uma Maheswari, C.; Anitha, A.; Princy, S.A. Combinatorial effects of aromatic 1,3-disubstituted ureas and fluoride on in vitro inhibition of Streptococcus mutans biofilm formation. Front. Microbiol. 2016, 7, 861. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Der-PS4: 1ALWKTLLKHVGKAAGKAALNAVTDMVNQ28 | |||
|---|---|---|---|
| Physico-Chemical Properties | Hydrophobicity (H) 0.368 | Hydrophobic Moment <μH> 0.226 | Net Charge 3 |
| Polar residues + GLY | Polar residues + GLY (n/%) 13/46.43 | Uncharged residues + GLY GLN 1, HIS 1, THR 2, ASN 2, GLY 2 | Charged residues LYS 4, ASP 1 |
| Nonpolar residues | Nonpolar residues (n/%) 15/53.57 | Aromatic residues TRP 1 | Special residues CYS 0, PRO 0 |
| Hydrophobic face: W A A L M A | |||
| Drugs | MIC/MBC (µM) | |||||
|---|---|---|---|---|---|---|
| S. aureus | MRSA | E. faecalis | E. coli | P. aeruginosa | C. albicans | |
| Der-PS4 | 4/8 | 8/16 | 32/32 | 8/16 | 16/32 | 4/16 |
| Melittin | 2/2 | 2/4 | 2/2 | 2/4 | 32/32 | 4/4 |
| Ampicillin | 0.3/0.3 | - | 4.8/4.8 | 36.6/36.6 | - | 146/- |
| Norfloxacin | 1.3/2.5 | 2.5/5.2 | 5.2/5.2 | 0.6/0.6 | 2.5/5.2 | 1.3/2.5 |
| S. aureus | MRSA | E. faecalis | E. coli | P. aeruginosa | |
|---|---|---|---|---|---|
| MBIC (μM) | 4 | 8 | 64 | 8 | 32 |
| MBEC (μM) | 64 | 64 | 128 | 32 | 256 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, D.; Zhou, X.; Chen, X.; Huang, L.; Xi, X.; Ma, C.; Zhou, M.; Wang, L.; Chen, T. Evaluating the Bioactivity of a Novel Antimicrobial and Anticancer Peptide, Dermaseptin-PS4(Der-PS4), from the Skin Secretion of Phyllomedusa sauvagii. Molecules 2019, 24, 2974. https://doi.org/10.3390/molecules24162974
Chen D, Zhou X, Chen X, Huang L, Xi X, Ma C, Zhou M, Wang L, Chen T. Evaluating the Bioactivity of a Novel Antimicrobial and Anticancer Peptide, Dermaseptin-PS4(Der-PS4), from the Skin Secretion of Phyllomedusa sauvagii. Molecules. 2019; 24(16):2974. https://doi.org/10.3390/molecules24162974
Chicago/Turabian StyleChen, Dong, Xiaowei Zhou, Xi Chen, Linyuan Huang, Xinping Xi, Chengbang Ma, Mei Zhou, Lei Wang, and Tianbao Chen. 2019. "Evaluating the Bioactivity of a Novel Antimicrobial and Anticancer Peptide, Dermaseptin-PS4(Der-PS4), from the Skin Secretion of Phyllomedusa sauvagii" Molecules 24, no. 16: 2974. https://doi.org/10.3390/molecules24162974
APA StyleChen, D., Zhou, X., Chen, X., Huang, L., Xi, X., Ma, C., Zhou, M., Wang, L., & Chen, T. (2019). Evaluating the Bioactivity of a Novel Antimicrobial and Anticancer Peptide, Dermaseptin-PS4(Der-PS4), from the Skin Secretion of Phyllomedusa sauvagii. Molecules, 24(16), 2974. https://doi.org/10.3390/molecules24162974