
Structural Insights into the Regulation Mechanism of Small GTPases by GEFs

Abstract
:1. Small GTPases and Their Regulators
1.1. An Overview
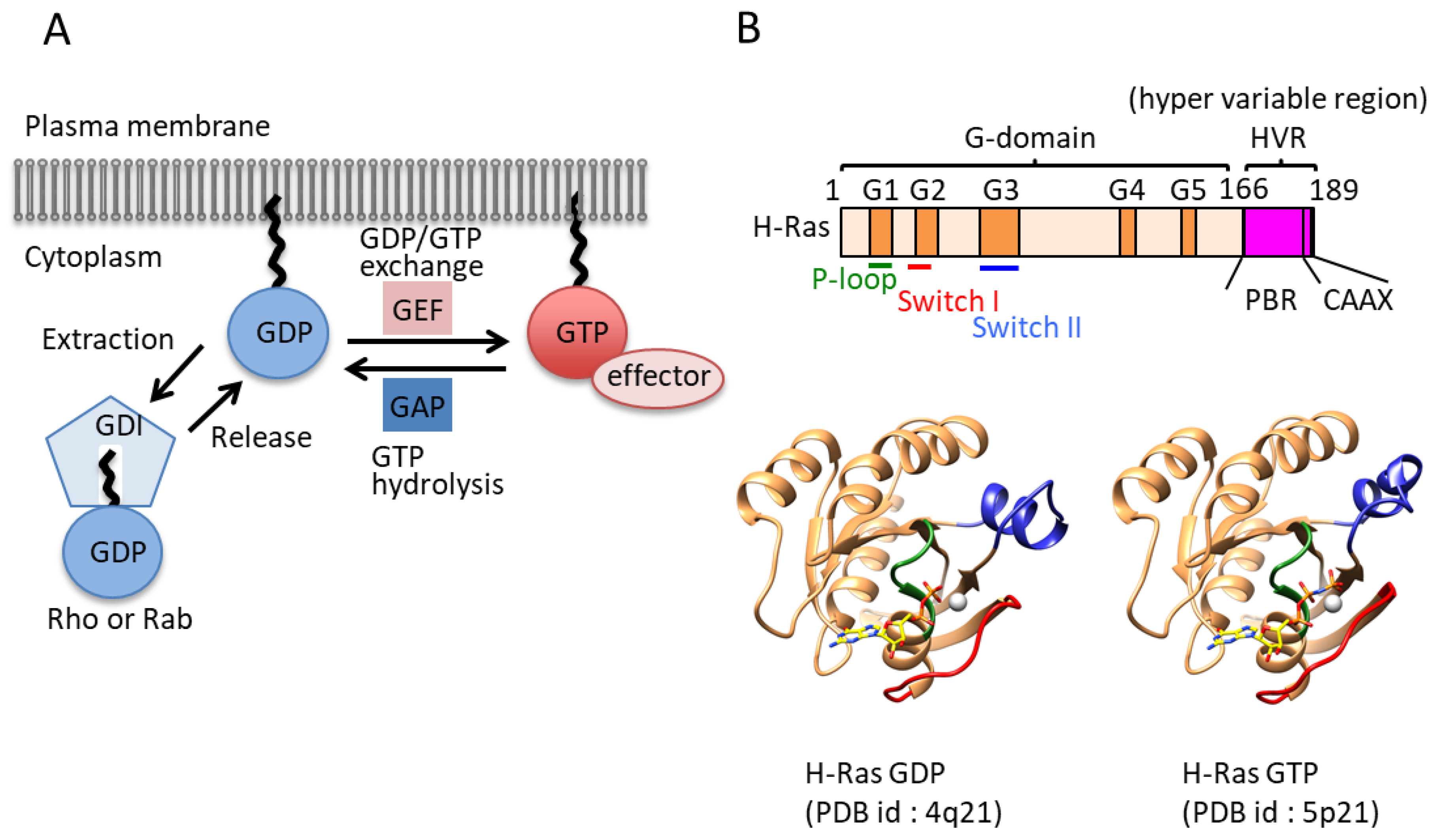
1.2. The GTP/GDP Cycle of GTPases by Their Regulators and PostTranslational Modifications
1.3. Small GTPase Families (Ras Superfamily)
1.4. The G Domain and the Molecular Switch Function
2. GEF Structures and Mechanisms of Small GTPase Activation
2.1. An Overview
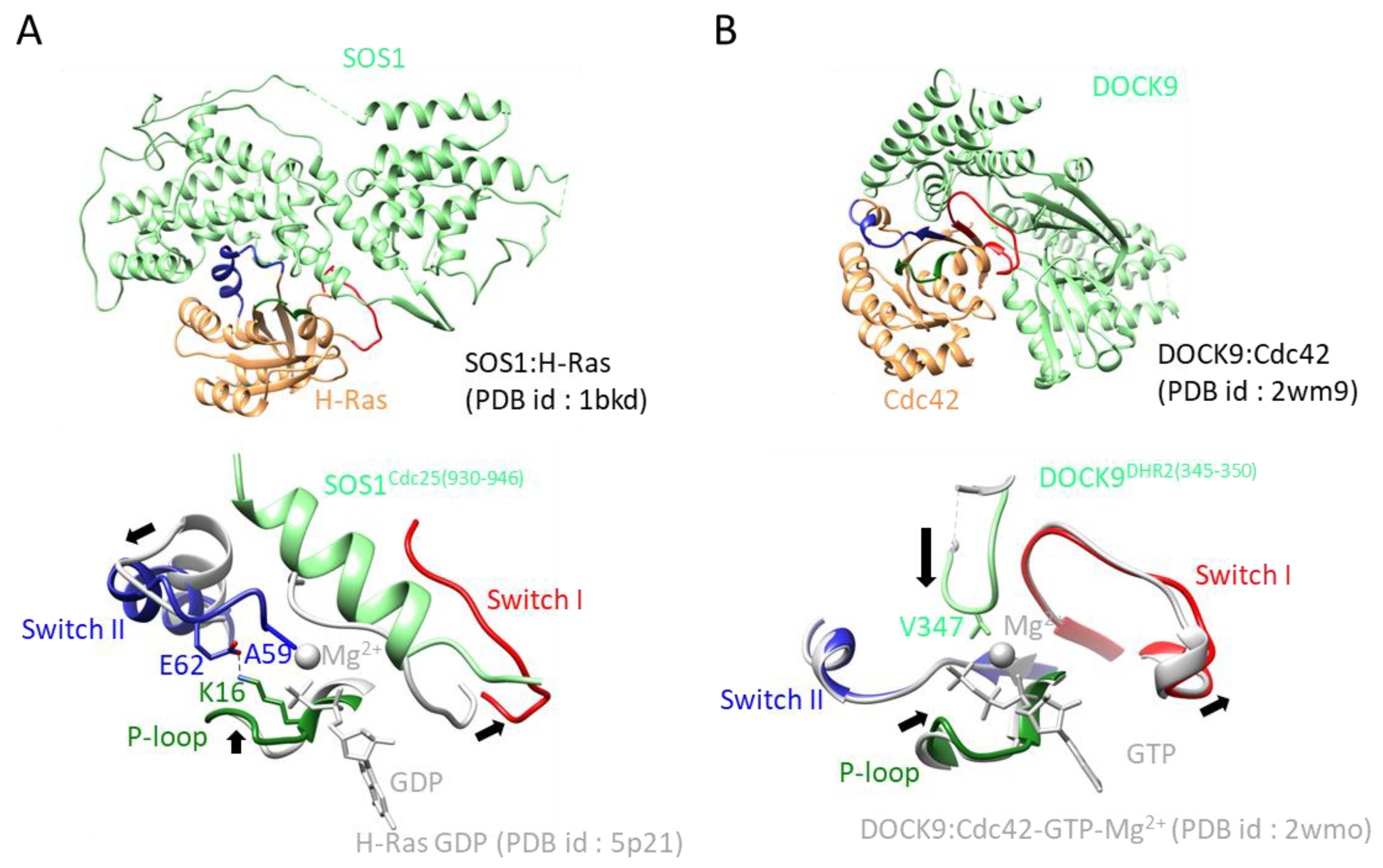
2.2. Common Mechanisms of GEF-Stimulated Exchange Reactions
2.3. Structures and Mechanisms of Family Specific GEFs
2.3.1. The Cdc 25 Family; GEFs for Ras, Ral, Rap, and Roc
2.3.2. DH, Dock, PRONE, and SmgGDS; GEFs for Rho Proteins
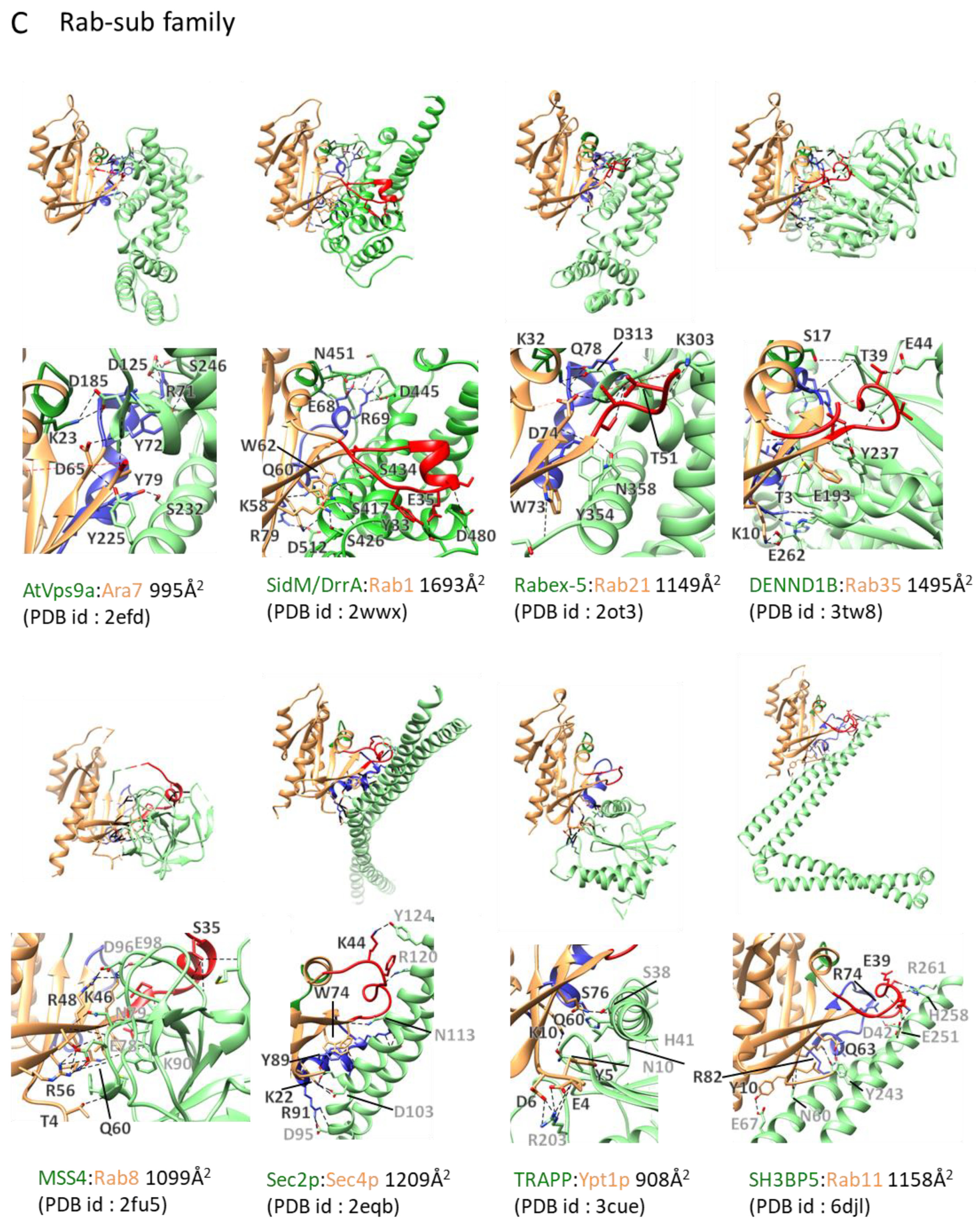
2.3.3. Rab GEFs
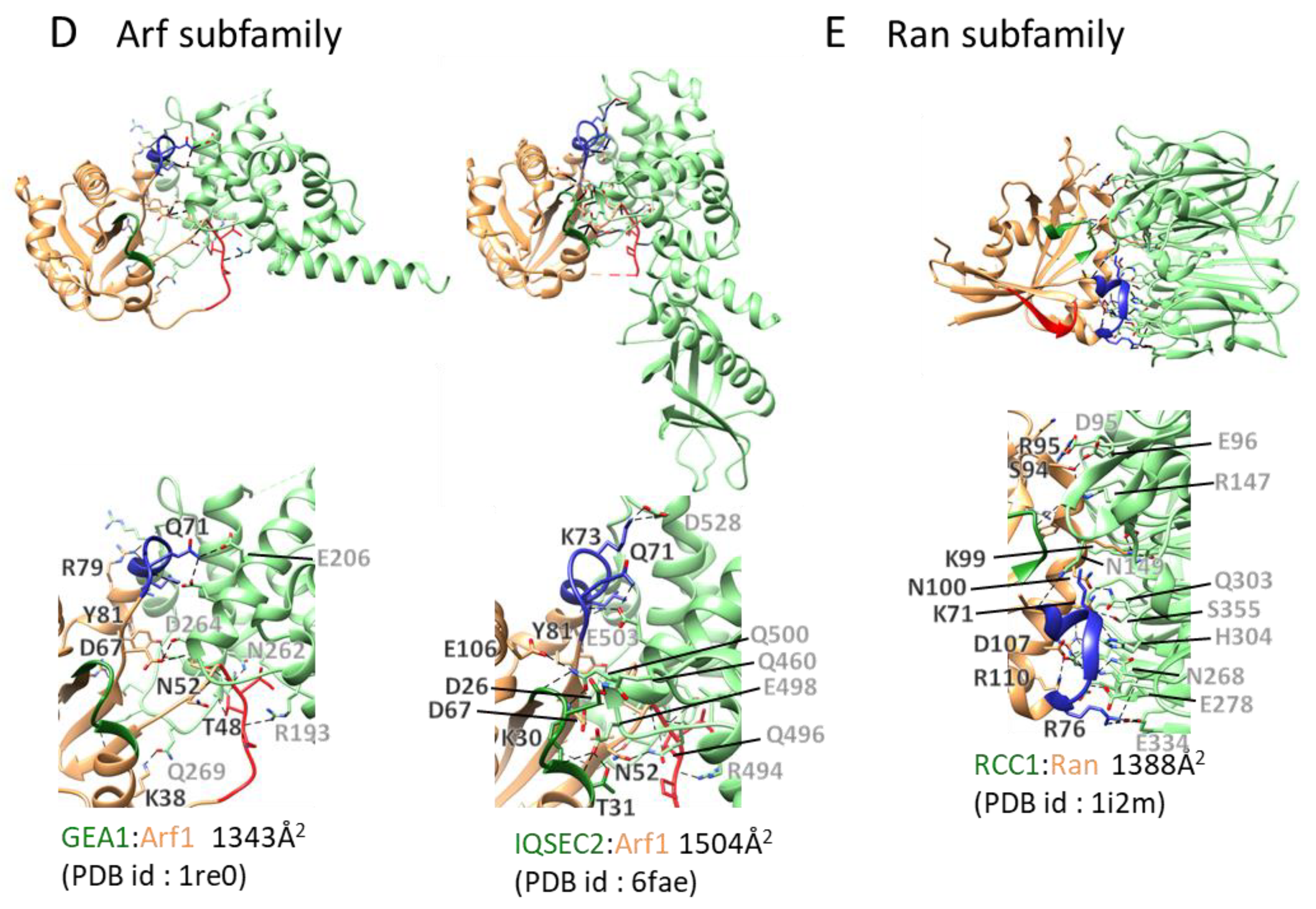
2.3.4. GEFs for Arf, Arf-like, and Sar GTPases
2.3.5. RanGEF
2.3.6. GEF for Mitochondrial Rho
2.4. Higher Level Regulatory Mechanisms of GEFs
2.4.1. The Feedback Loop of Rac/RasGEF SOS
2.4.2. The Feedback Loop of ArfGEF
2.4.3. Regulation by PH-DH Module
2.4.4. Regulation of GEF by Second Messenger
3. Local Protein Unfolding and Refolding of Small GTPase Induced by Binding
3.1. An Overview
3.2. MSS4
3.3. SmgGDS
4. Conclusions
Funding
Conflicts of Interest
References
- Wittinghofer, A.; Vetter, I.R. Structure-function relationships of the G domain, a canonical switch motif. Ann. Rev. Biochem. 2011, 80, 943–971. [Google Scholar] [CrossRef] [PubMed]
- Feinberg, A.P.; Vogelstein, B.; Droller, M.J.; Baylin, S.B.; Nelkin, B.D. Mutation affecting the 12th amino acid of the c-Ha-ras oncogene product occurs infrequently in human cancer. Science 1983, 220, 1175–1177. [Google Scholar] [CrossRef] [PubMed]
- Vetter, I.R.; Wittinghofer, A. The guanine nucleotide-binding switch in three dimensions. Science 2001, 294, 1299–1304. [Google Scholar] [CrossRef] [PubMed]
- Cherfils, J.; Zeghouf, M. Regulation of small GTPases by GEFs, GAPs, and GDIs. Physiol. Rev. 2013, 93, 269–309. [Google Scholar] [CrossRef] [PubMed]
- Bos, J.L.; Rehmann, H.; Wittinghofer, A. GEFs and GAPs: Critical elements in the control of small G proteins. Cell 2007, 129, 865–877. [Google Scholar] [CrossRef]
- Reiss, Y.; Goldstein, J.L.; Seabra, M.C.; Casey, P.J.; Brown, M.S. Inhibition of purified p21ras farnesyl: Protein transferase by Cys-AAX tetrapeptides. Cell 1990, 62, 81–88. [Google Scholar] [CrossRef]
- Seabra, M.C.; Reiss, Y.; Casey, P.J.; Brown, M.S.; Goldstein, J.L. Protein farnesyltransferase and geranylgeranyltransferase share a common alpha subunit. Cell 1991, 65, 429–434. [Google Scholar] [CrossRef]
- Seabra, M.C.; Goldstein, J.L.; Sudhof, T.C.; Brown, M.S. Rab geranylgeranyl transferase. A multisubunit enzyme that prenylates GTP-binding proteins terminating in Cys-X-Cys or Cys-Cys. J. Biol. Chem. 1992, 267, 14497–14503. [Google Scholar]
- Casey, P.J.; Seabra, M.C. Protein prenyltransferases. J. Biol. Chem. 1996, 271, 5289–5292. [Google Scholar] [CrossRef]
- Mitchell, A.L.; Attwood, T.K.; Babbitt, P.C.; Blum, M.; Bork, P.; Bridge, A.; Brown, S.D.; Chang, H.Y.; El-Gebali, S.; Fraser, M.I.; et al. InterPro in 2019: Improving coverage, classification and access to protein sequence annotations. Nucleic Acids Res. 2019, 47, D351–D360. [Google Scholar] [CrossRef]
- Valencia, A.; Chardin, P.; Wittinghofer, A.; Sander, C. The ras protein family: Evolutionary tree and role of conserved amino acids. Biochemistry 1991, 30, 4637–4648. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.E.; Saraste, M.; Runswick, M.J.; Gay, N.J. Distantly related sequences in the alpha- and beta-subunits of ATP synthase, myosin, kinases and other ATP-requiring enzymes and a common nucleotide binding fold. Embo J. 1982, 1, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Prive, G.G.; Milburn, M.V.; Tong, L.; de Vos, A.M.; Yamaizumi, Z.; Nishimura, S.; Kim, S.H. X-ray crystal structures of transforming p21 ras mutants suggest a transition-state stabilization mechanism for GTP hydrolysis. Proc. Natl. Acad. Sci. USA 1992, 89, 3649–3653. [Google Scholar] [CrossRef] [PubMed]
- Rensland, H.; John, J.; Linke, R.; Simon, I.; Schlichting, I.; Wittinghofer, A.; Goody, R.S. Substrate and product structural requirements for binding of nucleotides to H-ras p21: The mechanism of discrimination between guanosine and adenosine nucleotides. Biochemistry 1995, 34, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Milburn, M.V.; Tong, L.; deVos, A.M.; Brunger, A.; Yamaizumi, Z.; Nishimura, S.; Kim, S.H. Molecular switch for signal transduction: Structural differences between active and inactive forms of protooncogenic ras proteins. Science 1990, 247, 939–945. [Google Scholar] [CrossRef]
- Pai, E.F.; Kabsch, W.; Krengel, U.; Holmes, K.C.; John, J.; Wittinghofer, A. Structure of the guanine-nucleotide-binding domain of the Ha-ras oncogene product p21 in the triphosphate conformation. Nature 1989, 341, 209–214. [Google Scholar] [CrossRef]
- Huang, L.; Hofer, F.; Martin, G.S.; Kim, S.H. Structural basis for the interaction of Ras with RalGDS. Nat. Struct. Biol. 1998, 5, 422–426. [Google Scholar] [CrossRef]
- Boriack-Sjodin, P.A.; Margarit, S.M.; Bar-Sagi, D.; Kuriyan, J. The structural basis of the activation of Ras by Sos. Nature 1998, 394, 337–343. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Chardin, P.; Camonis, J.H.; Gale, N.W.; van Aelst, L.; Schlessinger, J.; Wigler, M.H.; Bar-Sagi, D. Human Sos1: A guanine nucleotide exchange factor for Ras that binds to GRB2. Science 1993, 260, 1338–1343. [Google Scholar] [CrossRef]
- Crechet, J.B.; Poullet, P.; Mistou, M.Y.; Parmeggiani, A.; Camonis, J.; Boy-Marcotte, E.; Damak, F.; Jacquet, M. Enhancement of the GDP-GTP exchange of RAS proteins by the carboxyl-terminal domain of SCD25. Science 1990, 248, 866–868. [Google Scholar] [CrossRef] [PubMed]
- Popovic, M.; Schouten, A.; Rensen-de Leeuw, M.; Rehmann, H. The structure of the Guanine Nucleotide Exchange Factor Rlf in complex with the small G-protein Ral identifies conformational intermediates of the exchange reaction and the basis for the selectivity. J. Struct. Biol. 2016, 193, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Vercoulen, Y.; Kondo, Y.; Iwig, J.S.; Janssen, A.B.; White, K.A.; Amini, M.; Barber, D.L.; Kuriyan, J.; Roose, J.P. A Histidine pH sensor regulates activation of the Ras-specific guanine nucleotide exchange factor RasGRP1. Elife 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Popovic, M.; Rensen-de Leeuw, M.; Rehmann, H. Selectivity of CDC25 homology domain-containing guanine nucleotide exchange factors. J. Mol. Biol. 2013, 425, 2782–2794. [Google Scholar] [CrossRef] [PubMed]
- Abbott, J.R.; Hodges, T.R.; Daniels, R.N.; Patel, P.A.; Kennedy, J.P.; Howes, J.E.; Akan, D.T.; Burns, M.C.; Sai, J.; Sobolik, T.; et al. Discovery of Aminopiperidine Indoles That Activate the Guanine Nucleotide Exchange Factor SOS1 and Modulate RAS Signaling. J. Med. Chem. 2018, 61, 6002–6017. [Google Scholar] [CrossRef] [PubMed]
- Hodges, T.R.; Abbott, J.R.; Little, A.J.; Sarkar, D.; Salovich, J.M.; Howes, J.E.; Akan, D.T.; Sai, J.; Arnold, A.L.; Browning, C.; et al. Discovery and Structure-Based Optimization of Benzimidazole-Derived Activators of SOS1-Mediated Nucleotide Exchange on RAS. J. Med. Chem. 2018, 61, 8875–8894. [Google Scholar] [CrossRef] [PubMed]
- Burns, M.C.; Howes, J.E.; Sun, Q.; Little, A.J.; Camper, D.V.; Abbott, J.R.; Phan, J.; Lee, T.; Waterson, A.G.; Rossanese, O.W.; et al. High-throughput screening identifies small molecules that bind to the RAS:SOS:RAS complex and perturb RAS signaling. Anal. Biochem. 2018, 548, 44–52. [Google Scholar] [CrossRef]
- Hillig, R.C.; Sautier, B.; Schroeder, J.; Moosmayer, D.; Hilpmann, A.; Stegmann, C.M.; Werbeck, N.D.; Briem, H.; Boemer, U.; Weiske, J.; et al. Discovery of potent SOS1 inhibitors that block RAS activation via disruption of the RAS-SOS1 interaction. Proc. Natl. Acad. Sci. USA 2019, 116, 2551–2560. [Google Scholar] [CrossRef]
- Eva, A.; Vecchio, G.; Rao, C.D.; Tronick, S.R.; Aaronson, S.A. The predicted DBL oncogene product defines a distinct class of transforming proteins. Proc. Natl. Acad. Sci. USA 1988, 85, 2061–2065. [Google Scholar] [CrossRef]
- Hart, M.J.; Eva, A.; Evans, T.; Aaronson, S.A.; Cerione, R.A. Catalysis of guanine nucleotide exchange on the CDC42Hs protein by the dbl oncogene product. Nature 1991, 354, 311–314. [Google Scholar] [CrossRef]
- Worthylake, D.K.; Rossman, K.L.; Sondek, J. Crystal structure of Rac1 in complex with the guanine nucleotide exchange region of Tiam1. Nature 2000, 408, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, Z.; Roe, S.M.; Marshall, C.J.; Barford, D. Activation of Rho GTPases by DOCK exchange factors is mediated by a nucleotide sensor. Science 2009, 325, 1398–1402. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, K.; Yang, J.; Zhang, Z.; Barford, D. Multiple factors confer specific Cdc42 and Rac protein activation by dedicator of cytokinesis (DOCK) nucleotide exchange factors. J. Biol. Chem. 2011, 286, 25341–25351. [Google Scholar] [CrossRef] [PubMed]
- Harada, Y.; Tanaka, Y.; Terasawa, M.; Pieczyk, M.; Habiro, K.; Katakai, T.; Hanawa-Suetsugu, K.; Kukimoto-Niino, M.; Nishizaki, T.; Shirouzu, M.; et al. DOCK8 is a Cdc42 activator critical for interstitial dendritic cell migration during immune responses. Blood 2012, 119, 4451–4461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kukimoto-Niino, M.; Tsuda, K.; Ihara, K.; Mishima-Tsumagari, C.; Honda, K.; Ohsawa, N.; Shirouzu, M. Structural Basis for the Dual Substrate Specificity of DOCK7 Guanine Nucleotide Exchange Factor. Structure 2019, 27, 741–748.e3. [Google Scholar] [CrossRef] [PubMed]
- Mucha, E.; Fricke, I.; Schaefer, A.; Wittinghofer, A.; Berken, A. Rho proteins of plants--functional cycle and regulation of cytoskeletal dynamics. Eur. J. Cell Biol. 2011, 90, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; Fricke, I.; Scrima, A.; Berken, A.; Wittinghofer, A. Structural evidence for a common intermediate in small G protein-GEF reactions. Mol. Cell 2007, 25, 141–149. [Google Scholar] [CrossRef]
- Thomas, C.; Fricke, I.; Weyand, M.; Berken, A. 3D structure of a binary ROP-PRONE complex: The final intermediate for a complete set of molecular snapshots of the RopGEF reaction. Biol. Chem. 2009, 390, 427–435. [Google Scholar] [CrossRef]
- Yamamoto, T.; Kaibuchi, K.; Mizuno, T.; Hiroyoshi, M.; Shirataki, H.; Takai, Y. Purification and characterization from bovine brain cytosol of proteins that regulate the GDP/GTP exchange reaction of smg p21s, ras p21-like GTP-binding proteins. J. Biol. Chem. 1990, 265, 16626–16634. [Google Scholar]
- Mizuno, T.; Kaibuchi, K.; Yamamoto, T.; Kawamura, M.; Sakoda, T.; Fujioka, H.; Matsuura, Y.; Takai, Y. A stimulatory GDP/GTP exchange protein for smg p21 is active on the post-translationally processed form of c-Ki-ras p21 and rhoA p21. Proc. Natl. Acad. Sci. USA 1991, 88, 6442–6446. [Google Scholar] [CrossRef]
- Hamel, B.; Monaghan-Benson, E.; Rojas, R.J.; Temple, B.R.; Marston, D.J.; Burridge, K.; Sondek, J. SmgGDS is a guanine nucleotide exchange factor that specifically activates RhoA and RhoC. J. Biol. Chem. 2011, 286, 12141–12148. [Google Scholar] [CrossRef] [PubMed]
- Berg, T.J.; Gastonguay, A.J.; Lorimer, E.L.; Kuhnmuench, J.R.; Li, R.; Fields, A.P.; Williams, C.L. Splice variants of SmgGDS control small GTPase prenylation and membrane localization. J. Biol. Chem. 2010, 285, 35255–35266. [Google Scholar] [CrossRef]
- Schuld, N.J.; Vervacke, J.S.; Lorimer, E.L.; Simon, N.C.; Hauser, A.D.; Barbieri, J.T.; Distefano, M.D.; Williams, C.L. The chaperone protein SmgGDS interacts with small GTPases entering the prenylation pathway by recognizing the last amino acid in the CAAX motif. J. Biol. Chem. 2014, 289, 6862–6876. [Google Scholar] [CrossRef]
- Williams, C.L. A new signaling paradigm to control the prenylation and trafficking of small GTPases. Cell Cycle 2013, 12, 2933–2934. [Google Scholar] [CrossRef]
- Shimizu, H.; Toma-Fukai, S.; Kontani, K.; Katada, T.; Shimizu, T. GEF mechanism revealed by the structure of SmgGDS-558 and farnesylated RhoA complex and its implication for a chaperone mechanism. Proc. Natl. Acad. Sci. USA 2018, 115, 9563–9568. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, H.; Toma-Fukai, S.; Saijo, S.; Shimizu, N.; Kontani, K.; Katada, T.; Shimizu, T. Structure-based analysis of the guanine nucleotide exchange factor SmgGDS reveals armadillo-repeat motifs and key regions for activity and GTPase binding. J. Biol. Chem. 2017, 292, 13441–13448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaiswal, M.; Dvorsky, R.; Ahmadian, M.R. Deciphering the molecular and functional basis of Dbl family proteins: A novel systematic approach toward classification of selective activation of the Rho family proteins. J. Biol. Chem. 2013, 288, 4486–4500. [Google Scholar] [CrossRef]
- Barr, F.; Lambright, D.G. Rab GEFs and GAPs. Curr. Opin. Cell Biol. 2010, 22, 461–470. [Google Scholar] [CrossRef]
- Stenmark, H. Rab GTPases as coordinators of vesicle traffic. Nat. Rev. Mol. Cell Biol. 2009, 10, 513–525. [Google Scholar] [CrossRef]
- Muller, M.P.; Goody, R.S. Molecular control of Rab activity by GEFs, GAPs and GDI. Small GTPases 2018, 9, 5–21. [Google Scholar] [CrossRef]
- Pereira-Leal, J.B.; Seabra, M.C. The mammalian Rab family of small GTPases: Definition of family and subfamily sequence motifs suggests a mechanism for functional specificity in the Ras superfamily. J. Mol. Biol. 2000, 301, 1077–1087. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Leal, J.B.; Seabra, M.C. Evolution of the Rab family of small GTP-binding proteins. J. Mol. Biol. 2001, 313, 889–901. [Google Scholar] [CrossRef] [PubMed]
- Itzen, A.; Pylypenko, O.; Goody, R.S.; Alexandrov, K.; Rak, A. Nucleotide exchange via local protein unfolding--structure of Rab8 in complex with MSS4. Embo J. 2006, 25, 1445–1455. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Fukai, S.; Ishitani, R.; Nureki, O. Crystal structure of the Sec4p.Sec2p complex in the nucleotide exchanging intermediate state. Proc. Natl. Acad. Sci. USA 2007, 104, 8305–8310. [Google Scholar] [CrossRef] [PubMed]
- Dong, G.; Medkova, M.; Novick, P.; Reinisch, K.M. A catalytic coiled coil: Structural insights into the activation of the Rab GTPase Sec4p by Sec2p. Mol. Cell 2007, 25, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, F.C.; Packer, M.; Collins, R. New insights into the molecular mechanism of the Rab GTPase Sec4p activation. BMC Struct. Biol. 2015, 15, 14. [Google Scholar] [CrossRef]
- Delprato, A.; Lambright, D.G. Structural basis for Rab GTPase activation by VPS9 domain exchange factors. Nat. Struct. Mol. Biol. 2007, 14, 406–412. [Google Scholar] [CrossRef]
- Cai, Y.; Chin, H.F.; Lazarova, D.; Menon, S.; Fu, C.; Cai, H.; Sclafani, A.; Rodgers, D.W.; De La Cruz, E.M.; Ferro-Novick, S.; et al. The structural basis for activation of the Rab Ypt1p by the TRAPP membrane-tethering complexes. Cell 2008, 133, 1202–1213. [Google Scholar] [CrossRef]
- Gray, J.L.; von Delft, F.; Brennan, P. Targeting the Small GTPase Superfamily through their Regulatory Proteins. Angew. Chem. Int. Ed. Engl. 2019. [Google Scholar] [CrossRef]
- Guo, Z.; Hou, X.; Goody, R.S.; Itzen, A. Intermediates in the guanine nucleotide exchange reaction of Rab8 protein catalyzed by guanine nucleotide exchange factors Rabin8 and GRAB. J. Biol. Chem. 2013, 288, 32466–32474. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, T.; Wang, S.; Gong, Z.; Tang, C.; Chen, J.; Ding, J. Molecular mechanism for Rabex-5 GEF activation by Rabaptin-5. Elife 2014, 3. [Google Scholar] [CrossRef] [Green Version]
- Goto-Ito, S.; Morooka, N.; Yamagata, A.; Sato, Y.; Sato, K.; Fukai, S. Structural basis of guanine nucleotide exchange for Rab11 by SH3BP5. Life Sci. Alliance 2019, 2. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, M.L.; Margaria, J.P.; Stariha, J.T.B.; Hoffmann, R.M.; McPhail, J.A.; Hamelin, D.J.; Boulanger, M.J.; Hirsch, E.; Burke, J.E. Structural determinants of Rab11 activation by the guanine nucleotide exchange factor SH3BP5. Nat. Commun. 2018, 9, 3772. [Google Scholar] [CrossRef] [PubMed]
- Cherfils, J.; Menetrey, J.; Mathieu, M.; Le Bras, G.; Robineau, S.; Beraud-Dufour, S.; Antonny, B.; Chardin, P. Structure of the Sec7 domain of the Arf exchange factor ARNO. Nature 1998, 392, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Renault, L.; Guibert, B.; Cherfils, J. Structural snapshots of the mechanism and inhibition of a guanine nucleotide exchange factor. Nature 2003, 426, 525–530. [Google Scholar] [CrossRef]
- Weissman, J.T.; Plutner, H.; Balch, W.E. The mammalian guanine nucleotide exchange factor mSec12 is essential for activation of the Sar1 GTPase directing endoplasmic reticulum export. Traffic 2001, 2, 465–475. [Google Scholar] [CrossRef] [PubMed]
- McMahon, C.; Studer, S.M.; Clendinen, C.; Dann, G.P.; Jeffrey, P.D.; Hughson, F.M. The structure of Sec12 implicates potassium ion coordination in Sar1 activation. J. Biol. Chem. 2012, 287, 43599–43606. [Google Scholar] [CrossRef]
- Renault, L.; Kuhlmann, J.; Henkel, A.; Wittinghofer, A. Structural basis for guanine nucleotide exchange on Ran by the regulator of chromosome condensation (RCC1). Cell 2001, 105, 245–255. [Google Scholar] [CrossRef]
- Nemergut, M.E.; Mizzen, C.A.; Stukenberg, T.; Allis, C.D.; Macara, I.G. Chromatin docking and exchange activity enhancement of RCC1 by histones H2A and H2B. Science 2001, 292, 1540–1543. [Google Scholar] [CrossRef] [PubMed]
- Makde, R.D.; England, J.R.; Yennawar, H.P.; Tan, S. Structure of RCC1 chromatin factor bound to the nucleosome core particle. Nature 2010, 467, 562–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, L.; Lei, Y.; Han, Y.; Li, Y.; Ji, X.; Liu, L. Vimar Is a Novel Regulator of Mitochondrial Fission through Miro. PLoS Genet. 2016, 12, e1006359. [Google Scholar] [CrossRef] [PubMed]
- Margarit, S.M.; Sondermann, H.; Hall, B.E.; Nagar, B.; Hoelz, A.; Pirruccello, M.; Bar-Sagi, D.; Kuriyan, J. Structural evidence for feedback activation by Ras.GTP of the Ras-specific nucleotide exchange factor SOS. Cell 2003, 112, 685–695. [Google Scholar] [CrossRef]
- Gureasko, J.; Kuchment, O.; Makino, D.L.; Sondermann, H.; Bar-Sagi, D.; Kuriyan, J. Role of the histone domain in the autoinhibition and activation of the Ras activator Son of Sevenless. Proc. Natl. Acad. Sci. USA 2010, 107, 3430–3435. [Google Scholar] [CrossRef] [Green Version]
- Sondermann, H.; Soisson, S.M.; Boykevisch, S.; Yang, S.S.; Bar-Sagi, D.; Kuriyan, J. Structural analysis of autoinhibition in the Ras activator Son of sevenless. Cell 2004, 119, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Pacheco-Rodriguez, G.; Moss, J.; Vaughan, M. Cytohesin-1: Structure, function, and ARF activation. Methods Enzymol. 2005, 404, 184–195. [Google Scholar] [CrossRef]
- DiNitto, J.P.; Delprato, A.; Gabe Lee, M.T.; Cronin, T.C.; Huang, S.; Guilherme, A.; Czech, M.P.; Lambright, D.G. Structural basis and mechanism of autoregulation in 3-phosphoinositide-dependent Grp1 family Arf GTPase exchange factors. Mol. Cell 2007, 28, 569–583. [Google Scholar] [CrossRef]
- Malaby, A.W.; van den Berg, B.; Lambright, D.G. Structural basis for membrane recruitment and allosteric activation of cytohesin family Arf GTPase exchange factors. Proc. Natl. Acad. Sci. USA 2013, 110, 14213–14218. [Google Scholar] [CrossRef] [Green Version]
- Rossman, K.L.; Der, C.J.; Sondek, J. GEF means go: Turning on RHO GTPases with guanine nucleotide-exchange factors. Nat. Rev. Mol. Cell Biol. 2005, 6, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Soisson, S.M.; Nimnual, A.S.; Uy, M.; Bar-Sagi, D.; Kuriyan, J. Crystal structure of the Dbl and pleckstrin homology domains from the human Son of sevenless protein. Cell 1998, 95, 259–268. [Google Scholar] [CrossRef]
- Nimnual, A.S.; Yatsula, B.A.; Bar-Sagi, D. Coupling of Ras and Rac guanosine triphosphatases through the Ras exchanger Sos. Science 1998, 279, 560–563. [Google Scholar] [CrossRef]
- Rossman, K.L.; Worthylake, D.K.; Snyder, J.T.; Siderovski, D.P.; Campbell, S.L.; Sondek, J. A crystallographic view of interactions between Dbs and Cdc42: PH domain-assisted guanine nucleotide exchange. Embo J. 2002, 21, 1315–1326. [Google Scholar] [CrossRef]
- Derewenda, U.; Oleksy, A.; Stevenson, A.S.; Korczynska, J.; Dauter, Z.; Somlyo, A.P.; Otlewski, J.; Somlyo, A.V.; Derewenda, Z.S. The crystal structure of RhoA in complex with the DH/PH fragment of PDZRhoGEF, an activator of the Ca(2+) sensitization pathway in smooth muscle. Structure 2004, 12, 1955–1965. [Google Scholar] [CrossRef] [PubMed]
- Chhatriwala, M.K.; Betts, L.; Worthylake, D.K.; Sondek, J. The DH and PH domains of Trio coordinately engage Rho GTPases for their efficient activation. J. Mol. Biol. 2007, 368, 1307–1320. [Google Scholar] [CrossRef] [PubMed]
- Baumeister, M.A.; Rossman, K.L.; Sondek, J.; Lemmon, M.A. The Dbs PH domain contributes independently to membrane targeting and regulation of guanine nucleotide-exchange activity. Biochem. J. 2006, 400, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Mitin, N.; Betts, L.; Yohe, M.E.; Der, C.J.; Sondek, J.; Rossman, K.L. Release of autoinhibition of ASEF by APC leads to CDC42 activation and tumor suppression. Nat. Struct. Mol. Biol. 2007, 14, 814–823. [Google Scholar] [CrossRef] [PubMed]
- Murayama, K.; Shirouzu, M.; Kawasaki, Y.; Kato-Murayama, M.; Hanawa-Suetsugu, K.; Sakamoto, A.; Katsura, Y.; Suenaga, A.; Toyama, M.; Terada, T.; et al. Crystal structure of the rac activator, Asef, reveals its autoinhibitory mechanism. J. Biol. Chem. 2007, 282, 4238–4242. [Google Scholar] [CrossRef] [PubMed]
- Vigil, D.; Cherfils, J.; Rossman, K.L.; Der, C.J. Ras superfamily GEFs and GAPs: Validated and tractable targets for cancer therapy? Nat. Rev. Cancer 2010, 10, 842–857. [Google Scholar] [CrossRef]
- Llorca, O.; Arias-Palomo, E.; Zugaza, J.L.; Bustelo, X.R. Global conformational rearrangements during the activation of the GDP/GTP exchange factor Vav3. Embo J. 2005, 24, 1330–1340. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.; Martins, I.R.; Li, P.; Amarasinghe, G.K.; Umetani, J.; Fernandez-Zapico, M.E.; Billadeau, D.D.; Machius, M.; Tomchick, D.R.; Rosen, M.K. Structural and energetic mechanisms of cooperative autoinhibition and activation of Vav1. Cell 2010, 140, 246–256. [Google Scholar] [CrossRef]
- Chrencik, J.E.; Brooun, A.; Zhang, H.; Mathews, I.I.; Hura, G.L.; Foster, S.A.; Perry, J.J.; Streiff, M.; Ramage, P.; Widmer, H.; et al. Structural basis of guanine nucleotide exchange mediated by the T-cell essential Vav1. J. Mol. Biol. 2008, 380, 828–843. [Google Scholar] [CrossRef]
- Raaijmakers, J.H.; Bos, J.L. Specificity in Ras and Rap signaling. J. Biol. Chem. 2009, 284, 10995–10999. [Google Scholar] [CrossRef]
- Li, P.; Martins, I.R.; Amarasinghe, G.K.; Rosen, M.K. Internal dynamics control activation and activity of the autoinhibited Vav DH domain. Nat. Struct. Mol. Biol. 2008, 15, 613–618. [Google Scholar] [CrossRef] [Green Version]
- Yohe, M.E.; Rossman, K.L.; Gardner, O.S.; Karnoub, A.E.; Snyder, J.T.; Gershburg, S.; Graves, L.M.; Der, C.J.; Sondek, J. Auto-inhibition of the Dbl family protein Tim by an N-terminal helical motif. J. Biol. Chem. 2007, 282, 13813–13823. [Google Scholar] [CrossRef]
- Yohe, M.E.; Rossman, K.; Sondek, J. Role of the C-terminal SH3 domain and N-terminal tyrosine phosphorylation in regulation of Tim and related Dbl-family proteins. Biochemistry 2008, 47, 6827–6839. [Google Scholar] [CrossRef]
- Jackson, L.P.; Kelly, B.T.; McCoy, A.J.; Gaffry, T.; James, L.C.; Collins, B.M.; Honing, S.; Evans, P.R.; Owen, D.J. A large-scale conformational change couples membrane recruitment to cargo binding in the AP2 clathrin adaptor complex. Cell 2010, 141, 1220–1229. [Google Scholar] [CrossRef]
- Yamaguchi, K.; Imai, K.; Akamatsu, A.; Mihashi, M.; Hayashi, N.; Shimamoto, K.; Kawasaki, T. SWAP70 functions as a Rac/Rop guanine nucleotide-exchange factor in rice. Plant J. 2012, 70, 389–397. [Google Scholar] [CrossRef]
- Gu, Y.; Li, S.; Lord, E.M.; Yang, Z. Members of a novel class of Arabidopsis Rho guanine nucleotide exchange factors control Rho GTPase-dependent polar growth. Plant. Cell 2006, 18, 366–381. [Google Scholar] [CrossRef]
- Hanawa-Suetsugu, K.; Kukimoto-Niino, M.; Mishima-Tsumagari, C.; Akasaka, R.; Ohsawa, N.; Sekine, S.; Ito, T.; Tochio, N.; Koshiba, S.; Kigawa, T.; et al. Structural basis for mutual relief of the Rac guanine nucleotide exchange factor DOCK2 and its partner ELMO1 from their autoinhibited forms. Proc. Natl. Acad. Sci. USA 2012, 109, 3305–3310. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.; Kinchen, J.M.; Rossman, K.L.; Grimsley, C.; deBakker, C.; Brugnera, E.; Tosello-Trampont, A.C.; Haney, L.B.; Klingele, D.; Sondek, J.; et al. PH domain of ELMO functions in trans to regulate Rac activation via Dock180. Nat. Struct. Mol. Biol. 2004, 11, 756–762. [Google Scholar] [CrossRef]
- Lu, M.; Kinchen, J.M.; Rossman, K.L.; Grimsley, C.; Hall, M.; Sondek, J.; Hengartner, M.O.; Yajnik, V.; Ravichandran, K.S. A Steric-inhibition model for regulation of nucleotide exchange via the Dock180 family of GEFs. Curr. Biol. 2005, 15, 371–377. [Google Scholar] [CrossRef]
- Medina, F.; Carter, A.M.; Dada, O.; Gutowski, S.; Hadas, J.; Chen, Z.; Sternweis, P.C. Activated RhoA is a positive feedback regulator of the Lbc family of Rho guanine nucleotide exchange factor proteins. J. Biol. Chem. 2013, 288, 11325–11333. [Google Scholar] [CrossRef]
- Chen, Z.; Medina, F.; Liu, M.Y.; Thomas, C.; Sprang, S.R.; Sternweis, P.C. Activated RhoA binds to the pleckstrin homology (PH) domain of PDZ-RhoGEF, a potential site for autoregulation. J. Biol. Chem. 2010, 285, 21070–21081. [Google Scholar] [CrossRef]
- Dada, O.; Gutowski, S.; Brautigam, C.A.; Chen, Z.; Sternweis, P.C. Direct regulation of p190RhoGEF by activated Rho and Rac GTPases. J. Struct. Biol. 2018, 202, 13–24. [Google Scholar] [CrossRef]
- Lin, Q.; Yang, W.; Baird, D.; Feng, Q.; Cerione, R.A. Identification of a DOCK180-related guanine nucleotide exchange factor that is capable of mediating a positive feedback activation of Cdc42. J. Biol. Chem. 2006, 281, 35253–35262. [Google Scholar] [CrossRef]
- De Rooij, J.; Zwartkruis, F.J.; Verheijen, M.H.; Cool, R.H.; Nijman, S.M.; Wittinghofer, A.; Bos, J.L. Epac is a Rap1 guanine-nucleotide-exchange factor directly activated by cyclic AMP. Nature 1998, 396, 474–477. [Google Scholar] [CrossRef]
- Kawasaki, H.; Springett, G.M.; Mochizuki, N.; Toki, S.; Nakaya, M.; Matsuda, M.; Housman, D.E.; Graybiel, A.M. A family of cAMP-binding proteins that directly activate Rap1. Science 1998, 282, 2275–2279. [Google Scholar] [CrossRef]
- Burton, J.L.; Burns, M.E.; Gatti, E.; Augustine, G.J.; De Camilli, P. Specific interactions of Mss4 with members of the Rab GTPase subfamily. Embo J. 1994, 13, 5547–5558. [Google Scholar] [CrossRef]
- Coppola, T.; Perret-Menoud, V.; Gattesco, S.; Magnin, S.; Pombo, I.; Blank, U.; Regazzi, R. The death domain of Rab3 guanine nucleotide exchange protein in GDP/GTP exchange activity in living cells. Biochem. J. 2002, 362, 273–279. [Google Scholar] [CrossRef]
- Muller-Pillasch, F.; Zimmerhackl, F.; Lacher, U.; Schultz, N.; Hameister, H.; Varga, G.; Friess, H.; Buchler, M.; Adler, G.; Gress, T.M. Cloning of novel transcripts of the human guanine-nucleotide-exchange factor Mss4: In situ chromosomal mapping and expression in pancreatic cancer. Genomics 1997, 46, 389–396. [Google Scholar] [CrossRef]
- Wada, M.; Nakanishi, H.; Satoh, A.; Hirano, H.; Obaishi, H.; Matsuura, Y.; Takai, Y. Isolation and characterization of a GDP/GTP exchange protein specific for the Rab3 subfamily small G proteins. J. Biol. Chem. 1997, 272, 3875–3878. [Google Scholar] [CrossRef]
- Walch-Solimena, C.; Collins, R.N.; Novick, P.J. Sec2p mediates nucleotide exchange on Sec4p and is involved in polarized delivery of post-Golgi vesicles. J. Cell Biol. 1997, 137, 1495–1509. [Google Scholar] [CrossRef]
- Esters, H.; Alexandrov, K.; Iakovenko, A.; Ivanova, T.; Thoma, N.; Rybin, V.; Zerial, M.; Scheidig, A.J.; Goody, R.S. Vps9, Rabex-5 and DSS4: Proteins with weak but distinct nucleotide-exchange activities for Rab proteins. J. Mol. Biol. 2001, 310, 141–156. [Google Scholar] [CrossRef]
- Burton, J.; Roberts, D.; Montaldi, M.; Novick, P.; De Camilli, P. A mammalian guanine-nucleotide-releasing protein enhances function of yeast secretory protein Sec4. Nature 1993, 361, 464–467. [Google Scholar] [CrossRef]
- Moya, M.; Roberts, D.; Novick, P. DSS4-1 is a dominant suppressor of sec4-8 that encodes a nucleotide exchange protein that aids Sec4p function. Nature 1993, 361, 460–463. [Google Scholar] [CrossRef]
- Nuoffer, C.; Wu, S.K.; Dascher, C.; Balch, W.E. Mss4 does not function as an exchange factor for Rab in endoplasmic reticulum to Golgi transport. Mol. Biol. Cell 1997, 8, 1305–1316. [Google Scholar] [CrossRef]
- Gulbranson, D.R.; Davis, E.M.; Demmitt, B.A.; Ouyang, Y.; Ye, Y.; Yu, H.; Shen, J. RABIF/MSS4 is a Rab-stabilizing holdase chaperone required for GLUT4 exocytosis. Proc. Natl. Acad. Sci. USA 2017, 114, E8224–E8233. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Schreiber, S.L. Structure of guanine-nucleotide-exchange factor human Mss4 and identification of its Rab-interacting surface. Nature 1995, 376, 788–791. [Google Scholar] [CrossRef]
- Zhu, Z.; Dumas, J.J.; Lietzke, S.E.; Lambright, D.G. A helical turn motif in Mss4 is a critical determinant of Rab binding and nucleotide release. Biochemistry 2001, 40, 3027–3036. [Google Scholar] [CrossRef]
- Williams, C.L. The polybasic region of Ras and Rho family small GTPases: A regulator of protein interactions and membrane association and a site of nuclear localization signal sequences. Cell Signal. 2003, 15, 1071–1080. [Google Scholar] [CrossRef]
- Ogita, Y.; Egami, S.; Ebihara, A.; Ueda, N.; Katada, T.; Kontani, K. Di-Ras2 Protein Forms a Complex with SmgGDS Protein in Brain Cytosol in Order to Be in a Low Affinity State for Guanine Nucleotides. J. Biol. Chem. 2015, 290, 20245–20256. [Google Scholar] [CrossRef] [Green Version]
- Bergom, C.; Hauser, A.D.; Rymaszewski, A.; Gonyo, P.; Prokop, J.W.; Jennings, B.C.; Lawton, A.J.; Frei, A.; Lorimer, E.L.; Aguilera-Barrantes, I.; et al. The Tumor-suppressive Small GTPase DiRas1 Binds the Noncanonical Guanine Nucleotide Exchange Factor SmgGDS and Antagonizes SmgGDS Interactions with Oncogenic Small GTPases. J. Biol. Chem. 2016, 291, 6534–6545. [Google Scholar] [CrossRef] [Green Version]
- Tew, G.W.; Lorimer, E.L.; Berg, T.J.; Zhi, H.; Li, R.; Williams, C.L. SmgGDS regulates cell proliferation, migration, and NF-kappaB transcriptional activity in non-small cell lung carcinoma. J. Biol. Chem. 2008, 283, 963–976. [Google Scholar] [CrossRef]
- Schuld, N.J.; Hauser, A.D.; Gastonguay, A.J.; Wilson, J.M.; Lorimer, E.L.; Williams, C.L. SmgGDS-558 regulates the cell cycle in pancreatic, non-small cell lung, and breast cancers. Cell Cycle 2014, 13, 941–952. [Google Scholar] [CrossRef] [Green Version]
- Zhi, H.; Yang, X.J.; Kuhnmuench, J.; Berg, T.; Thill, R.; Yang, H.; See, W.A.; Becker, C.G.; Williams, C.L.; Li, R. SmgGDS is up-regulated in prostate carcinoma and promotes tumour phenotypes in prostate cancer cells. J. Pathol. 2009, 217, 389–397. [Google Scholar] [CrossRef]
- Hauser, A.D.; Bergom, C.; Schuld, N.J.; Chen, X.; Lorimer, E.L.; Huang, J.; Mackinnon, A.C.; Williams, C.L. The SmgGDS splice variant SmgGDS-558 is a key promoter of tumor growth and RhoA signaling in breast cancer. Mol. Cancer Res. 2014, 12, 130–142. [Google Scholar] [CrossRef]
- Peifer, M.; Berg, S.; Reynolds, A.B. A repeating amino acid motif shared by proteins with diverse cellular roles. Cell 1994, 76, 789–791. [Google Scholar] [CrossRef]
- Kuhlmann, N.; Wroblowski, S.; Knyphausen, P.; de Boor, S.; Brenig, J.; Zienert, A.Y.; Meyer-Teschendorf, K.; Praefcke, G.J.; Nolte, H.; Krüger, M. Structural and mechanistic insights into the regulation of the fundamental Rho regulator RhoGDIα by lysine acetylation. J. Biol. Chem. 2016, 291, 5484–5499. [Google Scholar] [CrossRef]
- Garcia-Mata, R.; Boulter, E.; Burridge, K. The ‘invisible hand’: Regulation of RHO GTPases by RHOGDIs. Nat. Rev. Mol. Cell Biol. 2011, 12, 493. [Google Scholar] [CrossRef]
- Dharmaiah, S.; Bindu, L.; Tran, T.H.; Gillette, W.K.; Frank, P.H.; Ghirlando, R.; Nissley, D.V.; Esposito, D.; McCormick, F.; Stephen, A.G. Structural basis of recognition of farnesylated and methylated KRAS4b by PDEδ. Proc. Natl. Acad. Sci. USA 2016, 113, E6766–E6775. [Google Scholar] [CrossRef]
- Wang, M.; Casey, P.J. Protein prenylation: Unique fats make their mark on biology. Nat. Rev. Mol. Cell Biol. 2016, 17, 110. [Google Scholar] [CrossRef]
- Ahearn, I.M.; Haigis, K.; Bar-Sagi, D.; Philips, M.R. Regulating the regulator: Post-translational modification of RAS. Nat. Rev. Mol. Cell Biol. 2012, 13, 39. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | PDB ID | Year | GEF | SmallG | GEF | Reso | Ligand *1 | Disorder *2 (p,sw1,sw2) | HVR *3 |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 1LFD | 1998 | RALGDS | H-Ras | RasGEF | 2.1 | GNP | no, no, no | no |
| 2 | 1BKD | 1998 | SOS1 | H-Ras | RasGEF | 2.8 | no, no, no | no | |
| 3 | 1NVU | 2000 | SOS1 | H-Ras | RasGEF | 2.2 | no, no, no | no | |
| 4 | 1NVV | 2003 | SOS1 | H-Ras | RasGEF | 2.2 | GNP | no, no, no | no |
| 5 | 1NVW | 2003 | SOS1 | H-Ras | RasGEF | 2.7 | GNP | no, no, no | no |
| 6 | 1NVX | 2003 | SOS1 | H-Ras | RasGEF | 3.2 | GTP | no, no, no | no |
| 7 | 1XD2 | 2004 | SOS1 | H-Ras | RasGEF | 2.7 | GDP | no, no, no | no |
| 8 | 3CF6 | 2008 | Epac2 | Rap1B | RapGEF | 2.2 | SO4 | no, no, no | no |
| 9 | 4NYJ | 2014 | SOS1 | H-Ras | RasGEF | 2.9 | GNP | no, no, no | no |
| 10 | 4NYM | 2014 | SOS1 | H-Ras | RasGEF | 3.6 | no, no, no | no | |
| 11 | 4MGI | 2014 | Epac | Rap1b | RapGEF | 2.8 | SO4 | no, no, no | no |
| 12 | 4MGK | 2014 | Epac | Rap1b | RapGEF | 2.7 | SO4 | no, no, no | no |
| 13 | 4MGY | 2014 | Epac | Rap1b | RapGEF | 2.6 | SO4 | no, no, no | no |
| 14 | 4MGZ | 2014 | Epac | Rap1b | RapGEF | 3.0 | SO4 | no, no, no | no |
| 15 | 4MH0 | 2014 | Epac | Rap1b | RapGEF | 2.4 | SO4 | no, no, no | no |
| 16 | 4URU | 2015 | SOS1 | H-Ras | RasGEF | 2.8 | no, no, no | no | |
| 17 | 4URV | 2015 | SOS1 | H-Ras | RasGEF | 2.6 | no, no, no | no | |
| 18 | 4URW | 2015 | SOS1 | H-Ras | RasGEF | 2.8 | no, no, no | no | |
| 19 | 4URX | 2015 | SOS1 | H-Ras | RasGEF | 2.5 | no, no, no | no | |
| 20 | 4URY | 2015 | SOS1 | H-Ras | RasGEF | 2.5 | no, no, no | no | |
| 21 | 4URZ | 2015 | SOS1 | H-Ras | RasGEF | 2.2 | no, no, no | no | |
| 22 | 4US0 | 2015 | SOS1 | H-Ras | RasGEF | 2.2 | no, no, no | no | |
| 23 | 4US1 | 2015 | SOS1 | H-Ras | RasGEF | 2.7 | L71 | no, no, no | no |
| 24 | 4US2 | 2015 | SOS1 | H-Ras | RasGEF | 2.5 | L71 | no, no, no | no |
| 25 | 5CM8 | 2015 | Rlf (Rgl2) | RalA | RalGEF | 2.6 | no, no, no | no | |
| 26 | 6AXG | 2017 | RasGRP4 | H-Ras | RasGEF | 3.3 | no, yes, no | no | |
| 27 | 6AXF | 2017 | RasGRP | Rap1b | RapGEF | 3.1 | no, no, no | no | |
| 28 | 6D55 | 2018 | SOS1 | H-Ras | RasGEF | 1.7 | Na, FMT, GOL, Mg, GNP, FWA | no, no, no | no |
| 29 | 6D5W | 2018 | SOS1 | H-Ras | RasGEF | 2.5 | FVV, Mg, GNP | no, no, no | no |
| 30 | 6D56 | 2018 | SOS1 | H-Ras | RasGEF | 1.7 | Na, FMT, GOL, Mg, GNP, FVN | no, no, no | no |
| 31 | 6D59 | 2018 | SOS1 | H-Ras | RasGEF | 1.7 | Na, FMT, GOL, Mg, GNP, FVJ | no, no, no | no |
| 32 | 6D5E | 2018 | SOS1 | H-Ras | RasGEF | 1.8 | Na, FMT, GOL, Mg, GNP, FVG, CL | no, no, no | no |
| 33 | 6D5G | 2018 | SOS1 | H-Ras | RasGEF | 1.9 | FMT, GOL, Mg, GNP, FVD, Cl, BME | no, no, no | no |
| 34 | 6D5H | 2018 | SOS1 | H-Ras | RasGEF | 1.8 | FMT, GOL, MG, GNP, FV7, Cl | no, no, no | no |
| 35 | 6D5J | 2018 | SOS1 | H-Ras | RasGEF | 1.8 | Na, FMT, GOL, MG, GNP, FV4 | no, no, no | no |
| 36 | 6D5L | 2018 | SOS1 | H-Ras | RasGEF | 1.7 | Na, FMT, GOL, MG, GNP, FW7 | no, no, no | no |
| 37 | 6D5V | 2018 | SOS1 | H-Ras | RasGEF | 2.0 | Mg, GNP, FVY | no, no, no | no |
| 38 | 6D5M | 2018 | SOS1 | H-Ras | RasGEF | 2.1 | Mg, GNP, FW4 | no, no, no | no |
| 39 | 5WFO | 2018 | SOS1 | H-Ras | RasGEF | 2.0 | Mg, GNP, 5UU | no, no, no | no |
| 40 | 5WFP | 2018 | SOS1 | H-Ras | RasGEF | 2.1 | Mg, GNP, 5UX | no, no, no | no |
| 41 | 5WFQ | 2018 | SOS1 | H-Ras | RasGEF | 2.3 | Mg, GNP, 5UV | no, no, no | no |
| 42 | 5WFR | 2018 | SOS1 | H-Ras | RasGEF | 2.5 | Mg, GNP, 5UW | no, no, no | no |
| 43 | 6BVI | 2018 | SOS1 | H-Ras | RasGEF | 1.8 | NA, FMT, EC4, GOL, Mg, GNP | no, no, no | no |
| 44 | 6BVJ | 2018 | SOS1 | H-Ras | RasGEF | 1.7 | NA, FMT, EAS, GOL, Mg, GNP | no, no, no | no |
| 45 | 6BVK | 2018 | SOS1 | H-Ras | RasGEF | 1.8 | NA, FMT, EAV, GOL, Mg, GNP | no, no, no | no |
| 46 | 6BVL | 2018 | SOS1 | H-Ras | RasGEF | 1.7 | NA, FBY, EAV, GOL, Mg, GNP | no, no, no | no |
| 47 | 6BVM | 2018 | SOS1 | H-Ras | RasGEF | 1.8 | NA, FBY, EBV, GOL, Mg, GNP | no, no, no | no |
| 48 | 6EPL | 2019 | SOS1 | K-Ras | RasGEF | 2.6 | GOL | no, no, no | no |
| 49 | 6EPM | 2019 | SOS1 | K-Ras | RasGEF | 2.5 | BQ5, GOL | no, no, no | no |
| 50 | 6EPN | 2019 | SOS1 | K-Ras | RasGEF | 2.5 | BQ2, DMS, GOL | no, no, no | no |
| 51 | 6EPO | 2019 | SOS1 | K-Ras | RasGEF | 2.4 | GOL, BPW | no, no, no | no |
| 52 | 6EPP | 2019 | SOS1 | K-Ras | RasGEF | 2.4 | GOL, BOQ | no, no, no | no |
| 53 | 1LB1 | 2002 | Dbs (Mcf21) | RhoA | RhoGEF | 2.8 | no, no, no | no | |
| 54 | 1 × 86 | 2004 | ARHGEF12 | RhoA | RhoGEF | 3.2 | PO4 | no, no, no | no |
| 55 | 1XCG | 2004 | ARHGEF11 (PDZ-rhoGEF) | RhoA | RhoGEF | 2.5 | no, no, no | no | |
| 56 | 2RGN | 2007 | ARHGEF25 (p63RhoGEF) | RhoA | RhoGEF | 3.5 | no, no, no | no | |
| 57 | 3KZ1 | 2010 | ARHGEF11 (PDZ-rhoGEF) | RhoA | RhoGEF | 2.7 | GSP | no, no, no | no |
| 58 | 3LW8 | 2010 | IpgB2 | RhoA | RhoGEF | 1.9 | GDP | no, no, no | no |
| 59 | 3LWN | 2010 | IpgB2 | RhoA | RhoGEF | 2.3 | GDP | no, no, no | no |
| 60 | 3LXR | 2010 | IpgB2 | RhoA | RhoGEF | 1.7 | GDP | no, no, no | no |
| 61 | 3T06 | 2011 | ARHGEF11 (PDZ-rhoGEF) | RhoA | RhoGEF | 2.8 | no, no, no | no | |
| 62 | 4D0N | 2014 | AKAP13 | RhoA | RhoGEF | 2.1 | GDP | no, no, no | no |
| 63 | 4XH9 | 2015 | NET1 (ARHGEF8) | RhoA | RhoGEF | 2.0 | no, yes, no | no | |
| 64 | 6BC0 | 2017 | ARHGEF28 (p190RhoGEF) | RhoA | RhoGEF | 2.2 | GSP | no, no, no | no |
| 65 | 5JHG | 2017 | ARHGEF11 | RhoA | RhoGEF | 2.5 | GOL | no, no, no | no |
| 66 | 6BCA | 2017 | ARHGEF18 (LbcRhoGEF) | RhoA | RhoGEF | 2.0 | GSP | no, no, no | no |
| 67 | 6BCB | 2017 | ARHGEF18 (p114RhoGEF) | RhoA | RhoGEF | 1.4 | GSP | no, no, no | no |
| 68 | 5JHH | 2017 | ARHGEF11 | RhoA | RhoGEF | 2.3 | RAO | no, no, no | no |
| 69 | 5ZHX | 2018 | SmgGDS | RhoA | RhoGEF | 3.5 | yes, yes, yes | yes | |
| 70 | 1FOE | 2000 | Tiam | Rac1 | RhoGEF | 2.8 | SO4 | no, no, no | no |
| 71 | 2NZ8 | 2007 | Trio | Rac1 | RhoGEF | 2.0 | no, no, no | no | |
| 72 | 2VRW | 2008 | Vav1 | Rac1 | RhoGEF | 1.9 | no, no, no | no | |
| 73 | 2YIN | 2011 | DOCK2 | Rac1 | RhoGEF | 2.7 | no, no, no | no | |
| 74 | 3B13 | 2012 | DOCK2 | Rac1 | RhoGEF | 3.0 | no, no, no | no | |
| 75 | 3BJI | 2007 | Vav1 | Rac1 | RhoGEF | 2.6 | no, no, no | no | |
| 76 | 4YON | 2015 | P-Rex1 | Rac1 | RhoGEF | 2.0 | no, no, no | no | |
| 77 | 5FI0 | 2016 | P-Rex1 | Rac1 | RhoGEF | 3.3 | no, no, no | yes | |
| 78 | 5O33 | 2017 | Kalirin | Rac1 | RhoGEF | 1.6 | GDP | no, no, no | no |
| 79 | 6BC1 | 2018 | ARHGEF28 (p190RhoGEF) | Rac1 | RhoGEF | 2.9 | GSP | no, no, no | no |
| 80 | 1KZ7 | 2002 | Dbs (Mcf2l) | Cdc42 | RhoGEF | 2.4 | no, no, no | no | |
| 81 | 1GZS | 2002 | SOPE | Cdc42 | RhoGEF | 2.3 | PO4 | no, no, no | no |
| 82 | 1KI1 | 2002 | ITSN1 | Cdc42 | RhoGEF | 2.3 | PO4 | no, no, no | no |
| 83 | 1KZG | 2002 | Mcf2l (Dbs) | Cdc42 | RhoGEF | 2.6 | no, no, no | no | |
| 84 | 2DFK | 2006 | ARHGEF9 (Collybistin II) | Cdc42 | RhoGEF | 2.2 | PO4 | no, no, no | yes |
| 85 | 2WM9 | 2009 | DOCK9 | Cdc42 | RhoGEF | 2.2 | GOL | no, no, no | no |
| 86 | 2WMN | 2009 | DOCK9 | Cdc42 | RhoGEF | 2.4 | GDP | no, no, no | no |
| 87 | 2WMO | 2009 | DOCK9 | Cdc42 | RhoGEF | 2.2 | GDP | no, no, no | no |
| 88 | 3GCG | 2009 | map (L0028) | Cdc42 | RhoGEF | 2.3 | no, yes, no | no | |
| 89 | 3QBV | 2012 | ITSN1 | Cdc42 | RhoGEF | 2.7 | GDP | no, yes, no | no |
| 90 | 3VHL | 2012 | DOCK8 | Cdc42 | RhoGEF | 2.1 | PO4 | no, no, no | no |
| 91 | 6AJ4 | 2019 | DOCK7 | Cdc42 | RhoGEF | 3.2 | no, no, no | yes | |
| 92 | 6AJL | 2019 | DOCK7 | Cdc42 | RhoGEF | 3.2 | no, no, no | yes | |
| 93 | 3CX6 | 2008 | ARHGEF11 (PDZRhoGEF) | Galpha-13 | RhoGEF | 2.5 | GDP | no, no, no | no |
| 94 | 3CX7 | 2008 | ARHGEF11 (PDZRhoGEF) | Galpha-13 | RhoGEF | 2.3 | GSP | no, no, no | no |
| 95 | 3CX8 | 2008 | ARHGEF11 (PDZRhoGEF) | Galpha-13 | RhoGEF | 2.5 | GDP | no, no, no | no |
| 96 | 1SHZ | 2005 | ARHGEF1 (p115RhoGEF) | Gnai1,13 | RhoGEF | 2.9 | GDP+ALF | no, no, no | no |
| 97 | 2NTY | 2007 | ROPGEF8 | ROP4 | RopGEF | 3.1 | GDP | no, yes, no | no |
| 98 | 2WBL | 2009 | ROPGEF8 | ROP7 | RopGEF | 2.9 | no, yes, no | no | |
| 99 | 2WWX | 2009 | SidM/DrrA | Rab1 | RabGEF | 1.5 | no, no, no | no | |
| 100 | 3L0I | 2010 | SidM/DrrA | Rab1 | RabGEF | 2.9 | SO4 | no, no, no | no |
| 101 | 3JZA | 2010 | DrrA/SidM | Rab1b | RabGEF | 1.8 | PO4 | no, no, no | no |
| 102 | 5O74 | 2017 | DrrA/SidM | Rab1b | RabGEF | 2.5 | GDP | no, yes, yes | no |
| 103 | 2OT3 | 2007 | Rabex-5 (RabGEF1) | Rab21 | RabGEF | 2.1 | no, yes, no | no | |
| 104 | 3TW8 | 2011 | DENND1B | Rab35 | RabGEF | 2.1 | no, yes, no | no | |
| 105 | 4Q9U | 2014 | Rabex-5 (RabGEF1) | Rab5A | RabGEF | 4.6 | yes, yes, no | no | |
| 106 | 2FU5 | 2006 | MSS4 | Rab8A | RabGEF | 2.0 | no, yes, yes | no | |
| 107 | 4LHX | 2013 | RAB3IP (Rabin8) | Rab8A | RabGEF | 3.1 | SO4 | no, no, no | no |
| 108 | 4LHY | 2013 | RAB3IP (Rabin8) | Rab8A | RabGEF | 3.1 | GDP | no, no, no | no |
| 109 | 4LHZ | 2013 | RAB3IP (Rabin8) | Rab8A | RabGEF | 3.2 | GTP | no, no, no | no |
| 110 | 4LI0 | 2013 | RAB3IP (Rabin8) | Rab8A | RabGEF | 3.3 | GDP | no, no, no | no |
| 111 | 2EFD | 2010 | AtVps9a | Ara7 (RabF2B) | RabGEF | 3.0 | no, yes, no | no | |
| 112 | 2EFE | 2010 | AtVps9a | Ara7 (RabF2B) | RabGEF | 2.1 | GDP | no, no, no | no |
| 113 | 2EFH | 2010 | AtVps9a | Ara7 (RabF2B) | RabGEF | 2.1 | GDP | no, yes, no | no |
| 114 | 2EFC | 2010 | AtVps9a | Ara7 (RabF2B) | RabGEF | 2.1 | GDP | no, no, no | no |
| 115 | 4G01 | 2013 | Vps9a | Ara7 (RabF2B) | RabGEF | 2.2 | GDP | no, no, no | no |
| 116 | 2OCY | 2007 | Sec2p | Sec4p | RabGEF | 3.3 | GDP | no, no, no | no |
| 117 | 2EQB | 2007 | Sec2p | Sec4p | RabGEF | 2.7 | no, no, no | no | |
| 118 | 4ZDW | 2015 | Sec2p | Sec4p | RabGEF | 2.9 | no, no, no | no | |
| 119 | 3CUE | 2008 | TRAPPI assembly | Ypt1p | RabGEF | 3.7 | no, yes, no | no | |
| 120 | 5LDD | 2017 | Mon1-Ccz1 | Ypt7 | RabGEF | 2.5 | SO4 | no, no, no | no |
| 121 | 6IXV | 2019 | SH3BP5 | Rab11 | RabGEF | 3.8 | PO4 | no, no, no | no |
| 122 | 6EKK | 2019 | DENND 1A | Rab35 | RabGEF | 1.8 | GDP, SO4, EDO | no, no, no | no |
| 123 | 6DJL | 2019 | SH3BP5 | Rab11 | RabGEF | 3.1 | no, no, no | no | |
| 124 | 1RE0 | 2003 | GEA1 | ARF1 | ArfGEF | 2.4 | GDP | no, no, no | no |
| 125 | 1R8Q | 2003 | CYTH2 (Arno) | ARF1 | ArfGEF | 1.9 | G3D | no, no, no | no |
| 126 | 1R8S | 2003 | CYTH2 (Arno) | ARF1 | ArfGEF | 1.5 | GDP | no, no, no | no |
| 127 | 1S9D | 2003 | CYTH2 (Arno) | ARF1 | ArfGEF | 1.8 | GDP | no, yes, no | no |
| 128 | 4C0A | 2013 | IQSEC1 | ARF1 | ArfGEF | 3.3 | G3D | no, no, no | no |
| 129 | 6FAE | 2018 | IQSEC2 | ARF1 | ArfGEF | 2.3 | no, yes, no | no | |
| 130 | 4KAX | 2013 | CYTH3 | ARF6 | ArfGEF | 1.9 | GTP | no, no, no | no |
| 131 | 5EE5 | 2016 | ARFGEF1 | ARL1 | ArfGEF | 2.3 | GTP | no, no, no | no |
| 132 | 5J5C | 2016 | ARFGEF1 | ARL1 | ArfGEF | 3.4 | GTP | no, no, no | no |
| 133 | 1I2M | 2001 | RCC1 | RAN | RanGEF | 1.8 | SO4 | no, yes, no | no |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toma-Fukai, S.; Shimizu, T. Structural Insights into the Regulation Mechanism of Small GTPases by GEFs. Molecules 2019, 24, 3308. https://doi.org/10.3390/molecules24183308
Toma-Fukai S, Shimizu T. Structural Insights into the Regulation Mechanism of Small GTPases by GEFs. Molecules. 2019; 24(18):3308. https://doi.org/10.3390/molecules24183308
Chicago/Turabian StyleToma-Fukai, Sachiko, and Toshiyuki Shimizu. 2019. "Structural Insights into the Regulation Mechanism of Small GTPases by GEFs" Molecules 24, no. 18: 3308. https://doi.org/10.3390/molecules24183308
APA StyleToma-Fukai, S., & Shimizu, T. (2019). Structural Insights into the Regulation Mechanism of Small GTPases by GEFs. Molecules, 24(18), 3308. https://doi.org/10.3390/molecules24183308