Insight into the Modification of Phosphatidylcholine with n-3 Polyunsaturated Fatty Acids-Rich Ethyl Esters by Immobilized MAS1 Lipase



Abstract
:1. Introduction
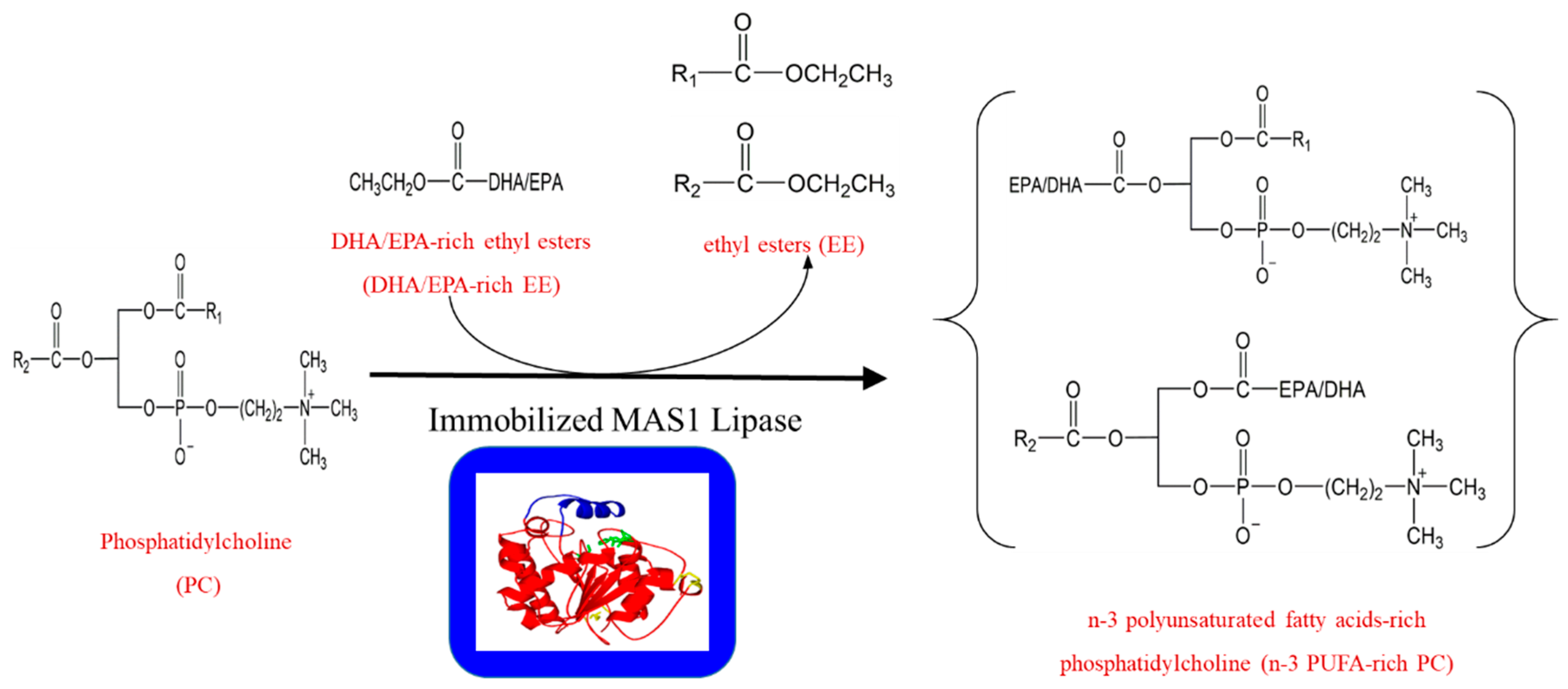
2. Results and Discussion
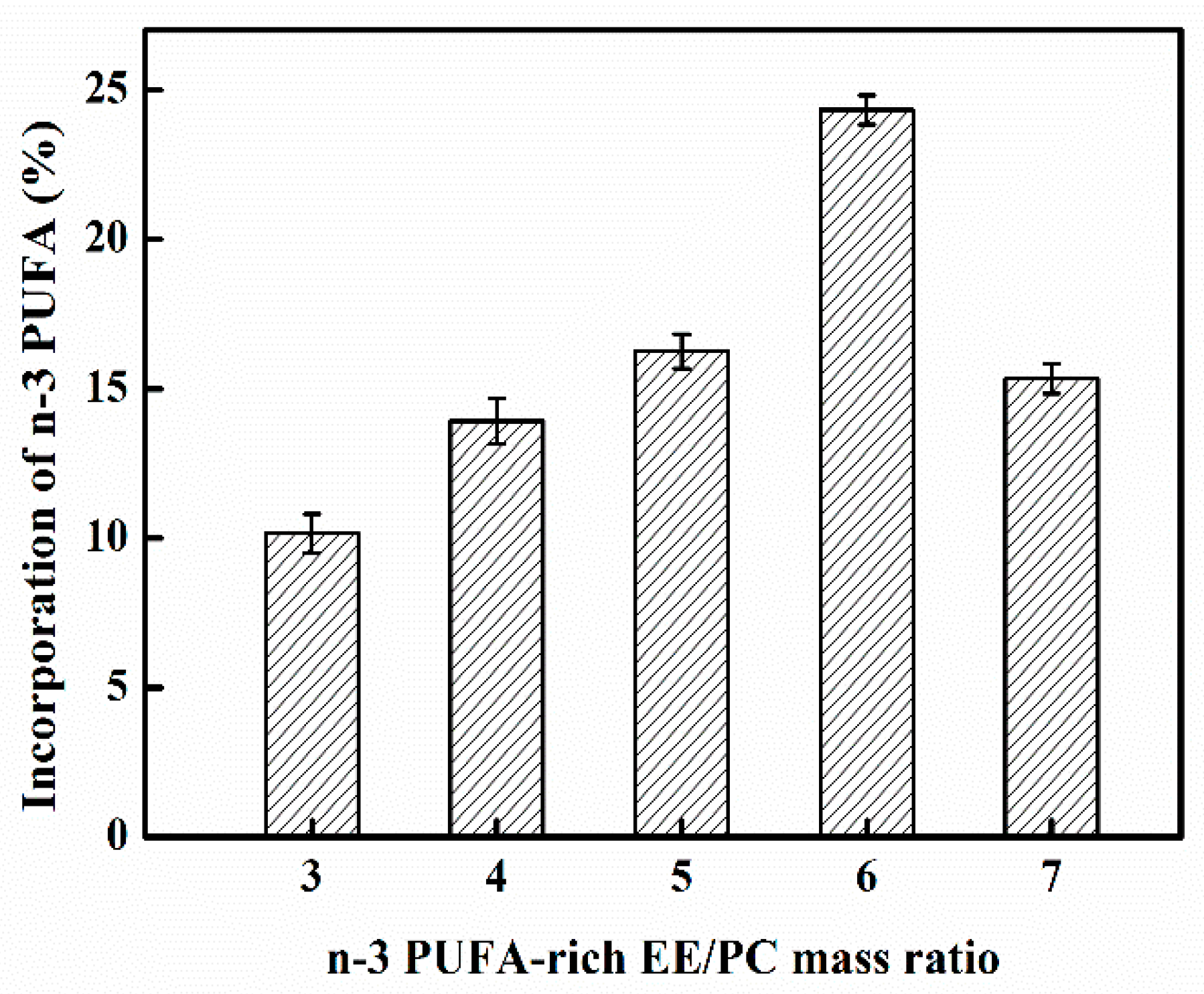
2.1. Effect of n-3 PUFA-Rich EE/PC Mass Ratio
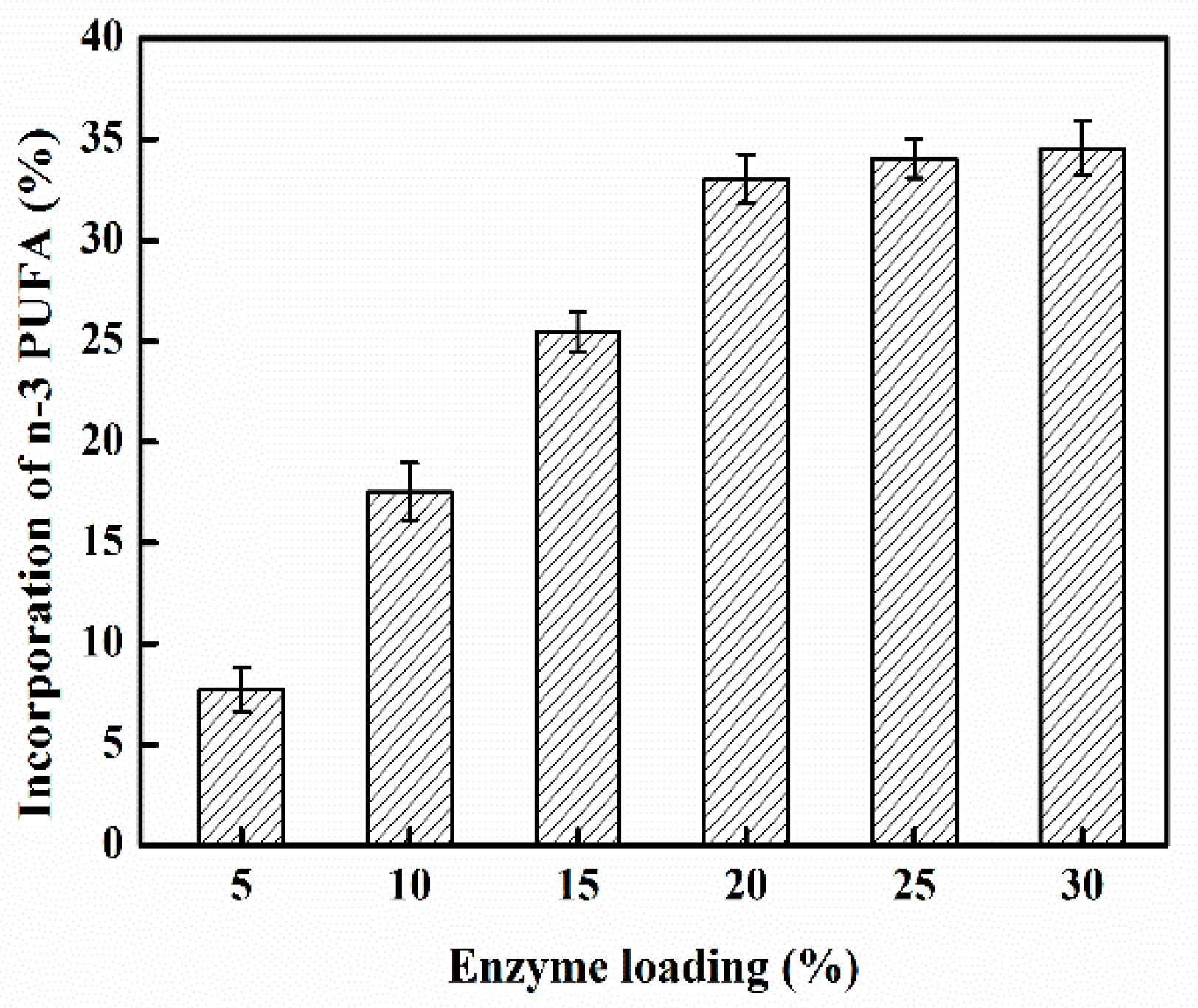
2.2. Effect of Enzyme Loading
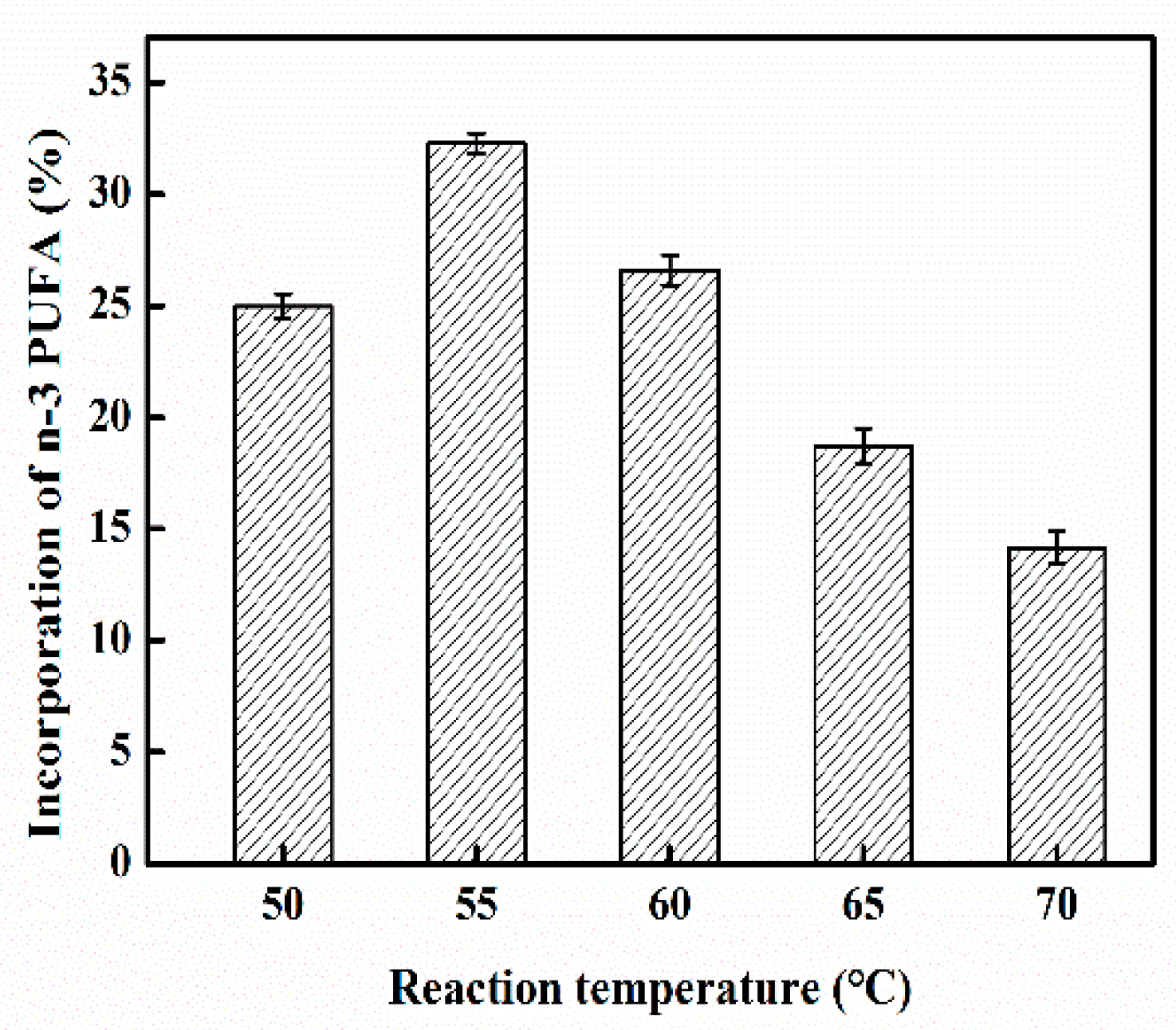
2.3. Effect of Reaction Temperature
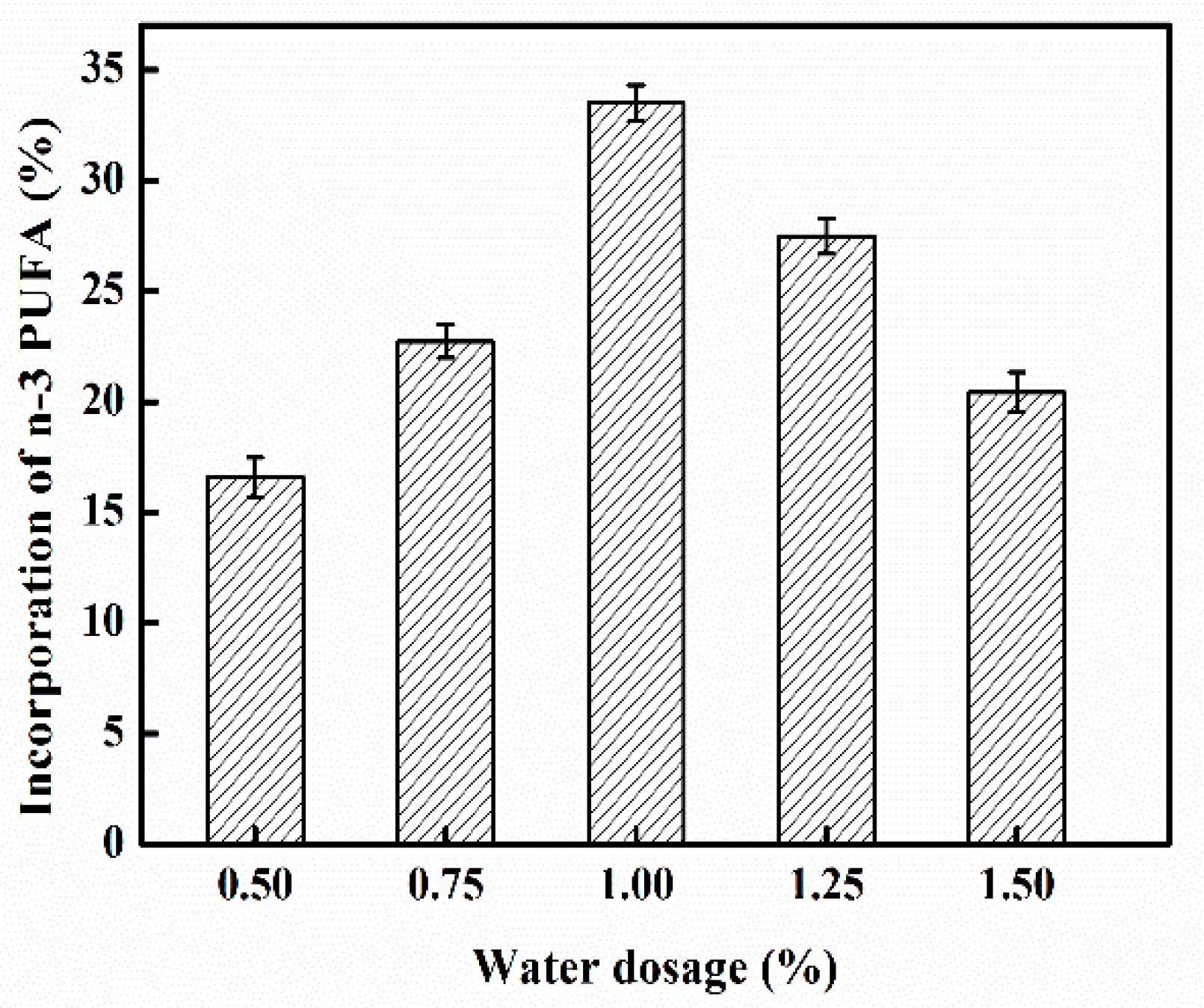
2.4. Effect of Water Dosage
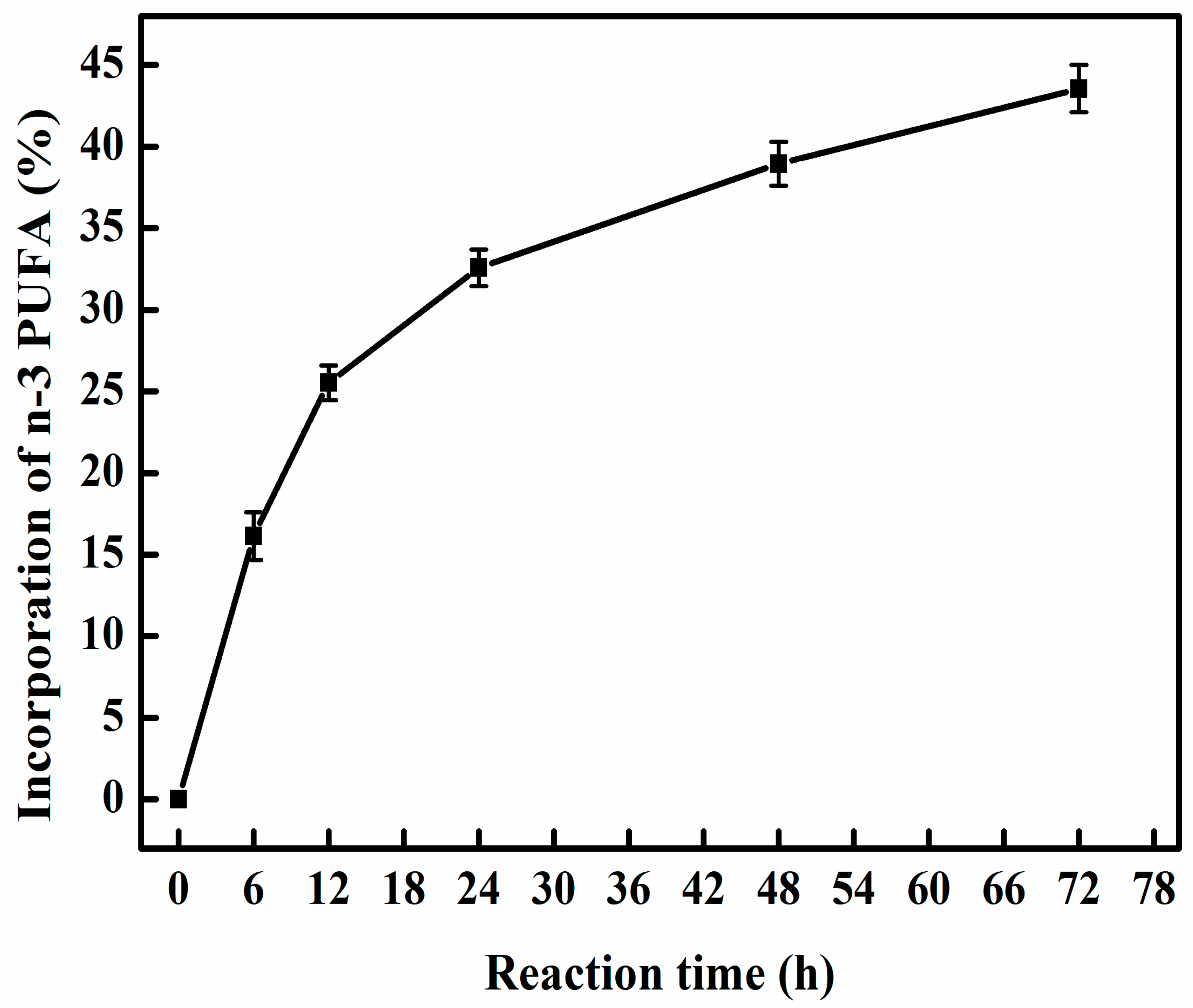
2.5. Time Course of Transesterification of PC with n-3 PUFA-Rich EE by Immobilized MAS1 Lipase
3. Materials and Methods
3.1. Materials
3.2. Immobilization of Lipase MAS1
3.3. Transesterification of PC with EPA/DHA-Rich EE
3.4. Analysis of FA Composition by GC
3.5. Analysis of PL Composition in the Final Products by 31P NMR
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PC | phosphatidylcholine |
| LPC | lysophosphatidylcholine |
| n-3 PUFA | n-3 polyunsaturated fatty acids |
| sn-1 LPC | 1-diacyl-sn-glycero-3-lysophosphatidylcholine |
| GPC | sn-glycero-3-phosphatidylcholine |
| sn-2 LPC | 2-diacyl-sn-glycero-3-lysophosphatidylcholine |
| NMR | nuclear magnetic resonance |
| EPA | eicosapentaenoic acid |
| DHA | docosahexaenoic acid |
| PL | phospholipids |
| EE | ethyl esters |
| FAME | fatty acid methyl esters |
| GC | gas chromatography |
References
- Cleland, L.G.; Caughey, G.E.; James, M.J.; Proudman, S.M. Reduction of cardiovascular risk factors with long term fish oil treatment in early rheumatoid arthritis. J. Rheumatol. 2006, 33, 1973–1979. [Google Scholar] [PubMed]
- Corsetto, P.A.; Montorfano, G.; Zava, S.; Jovenitti, I.E.; Cremona, A.; Berra, B.; Rizzo, A.M. Effects of n-3 PUFAs on breast cancer cells through their incorporation in plasma membrane. Lipids Health Dis. 2011, 10, 73. [Google Scholar] [CrossRef] [PubMed]
- Giudetti, A.M.; Cagnazzo, R. Beneficial effects of n-3 PUFA on Chronic airway inflammatory diseases. Prostag. Other Lipid Med. 2012, 99, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, T.; Khademi, H.; Moghadasian, M.H. The role of marine n-3 fatty acids in improving cardiovascular health: A review. Food Funct. 2013, 4, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Galli, C.; Sirtori, C.R.; Mosconi, C.; Medini, L.; Gianfranceschi, G.; Vaccarino, V.; Scolastico, C. Prolonged retention of doubly labeled phosphatidylcholine in human plasma and erythrocytes after oral administration. Lipids 1992, 27, 1005–1012. [Google Scholar] [CrossRef] [PubMed]
- Cansell, M.; Nacka, F.; Combe, N. Marine lipid-based liposomes increase in vivo FA bioavailability. Lipids 2003, 38, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Tecelão, C.; Silva, J.; Dubreucq, E.; Ribeiro, M.H.; Ferreira-Dias, S. Production of human milk fat substitutes enriched in omega-3 polyunsaturated fatty acids using immobilized commercial lipases and Candida parapsilosis lipase/acyltransferase. J. Mol. Catal. B Enzym. 2010, 65, 122–127. [Google Scholar]
- Mao, L.; Wang, M.; Li, Y.; Liu, Y.; Wang, J.; Xue, C. Eicosapentaenoic acid-containing phosphatidylcholine promotes osteogenesis: Mechanism of up-regulating Runx2 and ERK-mediated phosphorylation of PPARγ at serine 112. J. Funct. Foods 2019, 52, 73–80. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Harada, K.; Kasuga, S.; Hosokawa, M. Phospholipase A2-Mediated preparation of phosphatidylcholine containing ricinoleic acid and its anti-inflammatory effect on murine macrophage-like RAW264. 7 cells. Biocatal. Agric. Biotechnol. 2019, 19, 101141. [Google Scholar] [CrossRef]
- Ding, L.; Zhang, L.; Wen, M.; Che, H.; Du, L.; Wang, J.; Xue, C.; Xu, J.; Wang, Y. Eicosapentaenoic acid-enriched phospholipids improve atherosclerosis by mediating cholesterol metabolism. J. Funct. Foods 2017, 32, 90–97. [Google Scholar] [CrossRef]
- Liu, Y.; Shi, D.; Liu, Y.; Zhao, Y.; Tian, Y.; Xu, J.; Wang, J.; Xue, C. Eicosapentaenoic acid-containing phosphatidylcholine alleviated lipid accumulation in orotic acid-induced non-alcoholic fatty liver. J. Funct. Foods 2016, 23, 294–305. [Google Scholar] [CrossRef]
- Burri, L.; Hoem, N.; Banni, S.; Berge, K. Marine omega-3 phospholipids: Metabolism and biological activities. Int. J. Mol. Sci. 2012, 13, 15401–15419. [Google Scholar] [CrossRef] [PubMed]
- Carrie, I.; Smirnova, M.; Clement, M.; De Javel, D.; Frances, H.; Bourre, J.M. Docosahexaenoic acid-rich phospholipid supplementation: Effect on behavior, learning ability, and retinal function in control and n-3 polyunsaturated fatty acid deficient old mice. Nutr. Neurosci. 2002, 5, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.S.H.; Nielsen, N.S.; Baron, C.P.; Jacobsen, C. Marine phospholipids: The current understanding of their oxidation mechanisms and potential uses for food fortification. Crit. Rev. Food Sci. 2017, 57, 2057–2070. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Decker, E.A. Phospholipids in foods: Prooxidants or antioxidants? J. Sci. Food Agr. 2016, 96, 18–31. [Google Scholar] [CrossRef] [PubMed]
- van Hoogevest, P.; Wendel, A. The use of natural and synthetic phospholipids as pharmaceutical excipients. Eur. J. lipid Sci. Tech. 2014, 116, 1088–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Chen, R. Docosahexaenoic acid-containing phospholipids and triglycerides based nutritional supplements. Recent Pat. Food Nutr. Agric. 2010, 2, 213–220. [Google Scholar] [CrossRef]
- Iwasaki, Y.; Yamane, T. Phospholipases in enzyme engineering of phospholipids for food, cosmetics and medical applications. Lipid Biotechnol. 2002, 417–431. [Google Scholar]
- Akanbi, T.; Adcock, J.; Barrow, C. Selective concentration of EPA and DHA using Thermomyces lanuginosus lipase is due to fatty acid selectivity and not regioselectivity. Food Chem. 2013, 138, 615–620. [Google Scholar] [CrossRef]
- Vikbjerg, A.F.; Mu, H.; Xu, X. Parameters affecting incorporation and by-product formation during the production of structured phospholipids by lipase-catalyzed acidolysis in solvent-free system. J. Mol. Cataly. B Enzym. 2005, 36, 14–21. [Google Scholar] [CrossRef]
- Xi, X.; Feng, X.M.; Shi, N.R.; Ma, X.X.; Lin, H.; Han, Y.Q. Immobilized phospholipase A1-catalyzed acidolysis of phosphatidylcholine from Antarctic krill (Euphausia superba) for docosahexaenoic acid enrichment under supercritical conditions. J. Mol. Catal. B Enzym. 2016, 126, 46–55. [Google Scholar] [CrossRef]
- Li, D.M.; Qin, X.L.; Wang, W.F.; Li, Z.G.; Yang, B.; Wang, Y.H. Synthesis of DHA/EPA-rich phosphatidylcholine by immobilized phospholipase A1-effect of water addition and vacuum condition. Bioprocess. Biosyst. Eng. 2016, 39, 1305–1314. [Google Scholar] [CrossRef] [PubMed]
- Kosugi, Y.; Azuma, N. Synthesis of triacylglycerol from polyunsaturated fatty acids by immobilized lipase. J. Am. Oil. Chem. Soc. 1994, 71, 1397–1403. [Google Scholar] [CrossRef]
- Kralovec, J.A.; Wang, W.J.; Barrow, C.J. Production of omega-3 triacylglycerol concentrates using a new food grade immobilized Candida Antarctica lipase B. Aust. J. Chem. 2010, 63, 922–928. [Google Scholar] [CrossRef]
- Hama, S.; Miura, K.; Yoshida, A.; Noda, H.; Fukuda, H.; Kondo, A. Transesterification of phosphatidylcholine in sn-1 position through direct use of lipase-producing Rhizopus oryzae cells as whole-cell biocatalyst. Appl. Microbial. Biot. 2011, 90, 1731–1738. [Google Scholar] [CrossRef] [PubMed]
- Park, C.W.; Kwon, S.J.; Han, J.J.; Rhee, J.S. Transesterification of phosphatidylcholine with eicosapentaenoic acid ethyl ester using phospholipase A2 in organic solvent. Biotechnol. Let. 2000, 22, 147–150. [Google Scholar] [CrossRef]
- Li, X.; Chen, J.F.; Yang, B.; Li, D.M.; Wang, Y.H.; Wang, W.F. Production of structured Phosphatidylcholine with high content of DHA/EPA by immobilized phospholipase A1-catalyzed transesterification. Int. J. Mol. Sci. 2014, 15, 15244–15258. [Google Scholar] [CrossRef]
- Marsaoui, N.; Naghmouchi, K.; Baah, B.; Raies, A.; Laplante, S. Incorportation of ethyl esters of EPA and DHA in soybean lecithin using Rhizomucor miehei lipase: Effect of additives and solvent-free conditions. Appl. Biochem. Biotechnol. 2015, 176, 938–946. [Google Scholar] [CrossRef] [PubMed]
- Xing, X.; Jia, J.Q.; Zhang, J.F.; Zhou, Z.W.; Li, J.; Wang, N.; Yu, X.Q. CALB Immobilized onto Magnetic Nanoparticles for Efficient Kinetic Resolution of Racemic Secondary Alcohols: Long-Term Stability and Reusability. Molecules 2019, 24, 490. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Jiang, M.Y.; Shi, J.; Zheng, M.M.; Huang, F.H. Preparation of Immobilized Lipase Based on Hollow Mesoporous Silica Spheres and Its Application in Ester Synthesis. Molecules 2019, 24, 395. [Google Scholar] [CrossRef]
- Yuan, D.J.; Lan, D.M.; Xin, R.P.; Yang, B.; Wang, Y.H. Screening and characterization of a thermostable lipase from marine Streptomyces sp. Strain W007. Biotechnol. Appl. Bioc. 2015, 63, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Lan, D.M.; Qu, M.; Yang, B.; Wang, Y.H. Enhancing production of lipase MAS1 from marine Streptomyces sp. strain in Pichia pastoris by chaperones co-expression. Electron. J. Biotechn. 2016, 22, 62–67. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.M.; Li, D.M.; Wang, W.F.; Yang, B.; Wang, Y.H. A highly efficient immobilized MAS1 lipase for the glycerolysis reaction of n-3 PUFA-rich ethyl esters. J. Mol. Catal. B: Enzym. 2016, 134, 25–31. [Google Scholar] [CrossRef]
- Basso, A.; Froment, L.; Hesseler, M.; Serban, S. New highly robust divinyl benzene/acrylate polymer for immobilization of lipase CALB. Eur. J. Lipid Sci. Technol. 2013, 115, 468–472. [Google Scholar] [CrossRef]
- Wang, W.F.; Xu, Y.; Qin, X.L.; Lan, D.M.; Yang, B.; Wang, Y.H. Immobilization of lipase SMG1 and its application in synthesis partical glycerides. Eur. J. Lipid Sci. Technol. 2014, 116, 1063–1069. [Google Scholar] [CrossRef]
- Kim, I.; García, H.S.; Hill, C.G. Phospholipase A1-catalyzed synthesis of phospholipids enriched in n-3 polyunsaturated fatty acid residues. Enzym. Microb. Technol. 2007, 40, 1130–1135. [Google Scholar] [CrossRef]
- Wang, Y.H.; Mai, Q.Y.; Qin, X.L.; Yang, B.; Wang, Z.L.; Chen, H.T. Establishment of an evaluation model for human milk fat substitutes. J. Agric. Food Chem. 2010, 58, 642–649. [Google Scholar] [CrossRef]
- Qin, X.L.; Wang, Y.M.; Wang, Y.H.; Huang, H.H.; Yang, B. Preparation and characterization of 1,3-dioleoyl-2-palmitoylglycerol. J. Agric. Food Chem. 2011, 59, 5714–5719. [Google Scholar] [CrossRef]
- Marsaoui, N.; Laplante, S.; Raies, A.; Naghmouchi, K. Incorporation of omega-3 polyunsaturated fatty acids into soybean lecithin: Effect of amines and divalent cations on transesterification by lipases. World J. Microb. Biot. 2013, 29, 2233–2238. [Google Scholar] [CrossRef]
Sample Availability: Not available. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phospholipids | Content (%) | |
|---|---|---|
| Original PC | Structured PC | |
| PC | 97.96 ± 0.45 | 32.68 ± 0.32 |
| sn-1 LPC | 1.76 ± 0.23 | 28.76 ± 0.58 |
| sn-2 LPC | 0.28 ± 0.22 | 4.90 ± 0.21 |
| GPC | - | 33.66 ± 0.67 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Qin, X.; Li, X.; Zhao, Z.; Yang, B.; Wang, Y. Insight into the Modification of Phosphatidylcholine with n-3 Polyunsaturated Fatty Acids-Rich Ethyl Esters by Immobilized MAS1 Lipase. Molecules 2019, 24, 3528. https://doi.org/10.3390/molecules24193528
Wang X, Qin X, Li X, Zhao Z, Yang B, Wang Y. Insight into the Modification of Phosphatidylcholine with n-3 Polyunsaturated Fatty Acids-Rich Ethyl Esters by Immobilized MAS1 Lipase. Molecules. 2019; 24(19):3528. https://doi.org/10.3390/molecules24193528
Chicago/Turabian StyleWang, Xiumei, Xiaoli Qin, Xiuting Li, Zexin Zhao, Bo Yang, and Yonghua Wang. 2019. "Insight into the Modification of Phosphatidylcholine with n-3 Polyunsaturated Fatty Acids-Rich Ethyl Esters by Immobilized MAS1 Lipase" Molecules 24, no. 19: 3528. https://doi.org/10.3390/molecules24193528
APA StyleWang, X., Qin, X., Li, X., Zhao, Z., Yang, B., & Wang, Y. (2019). Insight into the Modification of Phosphatidylcholine with n-3 Polyunsaturated Fatty Acids-Rich Ethyl Esters by Immobilized MAS1 Lipase. Molecules, 24(19), 3528. https://doi.org/10.3390/molecules24193528