Separation and Lipid Inhibition Effects of a Novel Decapeptide from Chlorella pyenoidose


Abstract
:1. Introduction
2. Results
2.1. Characterization of the Extracted Proteins and Enzymatic Hydrolysates
2.2. PL inhibitory Activities of Proteins and Enzymatic Hydrolysates
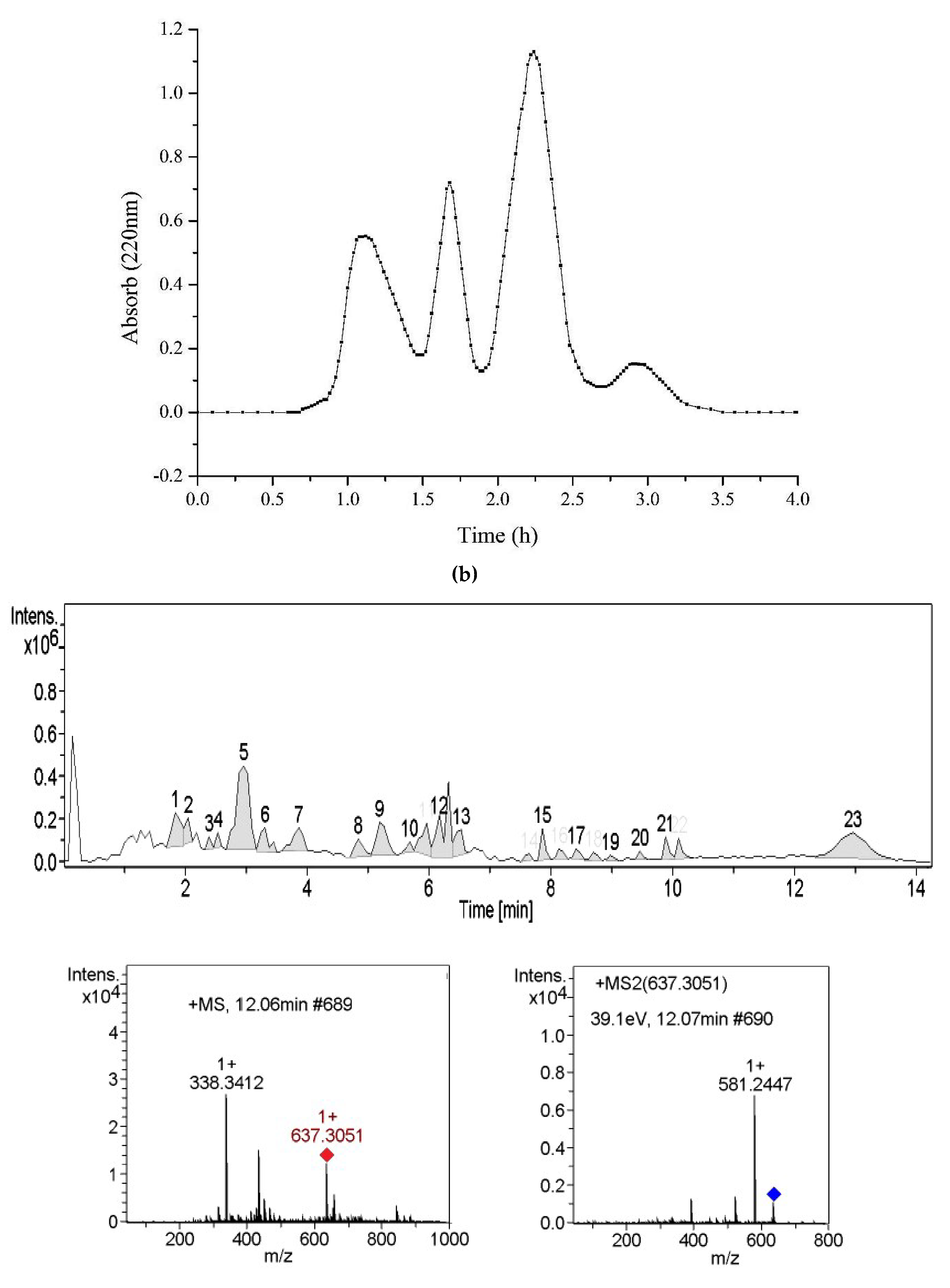
2.3. Separation of Alcalase Hydrolysates and PL Inhibition
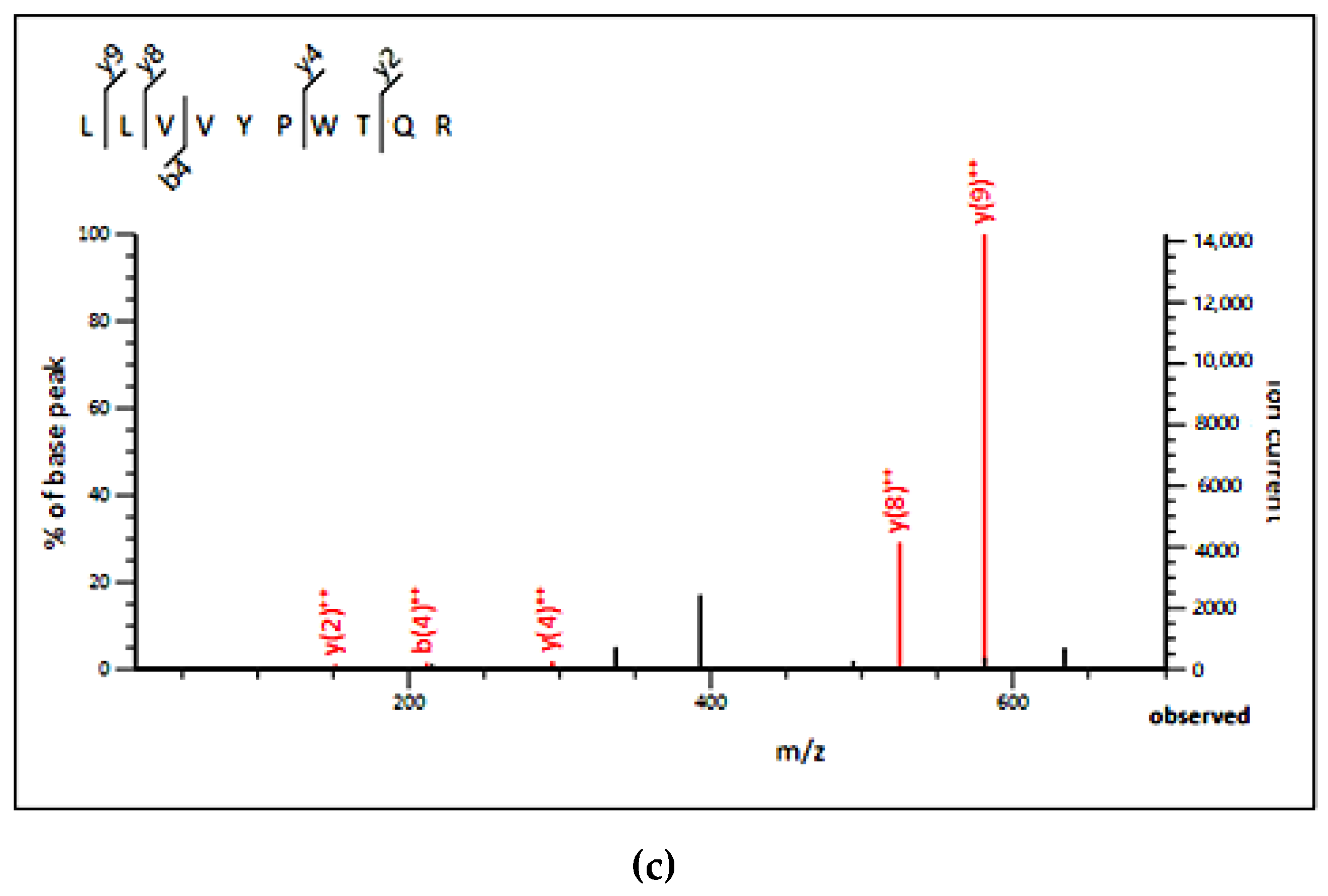
2.4. Identification of PL Inhibitory Peptides
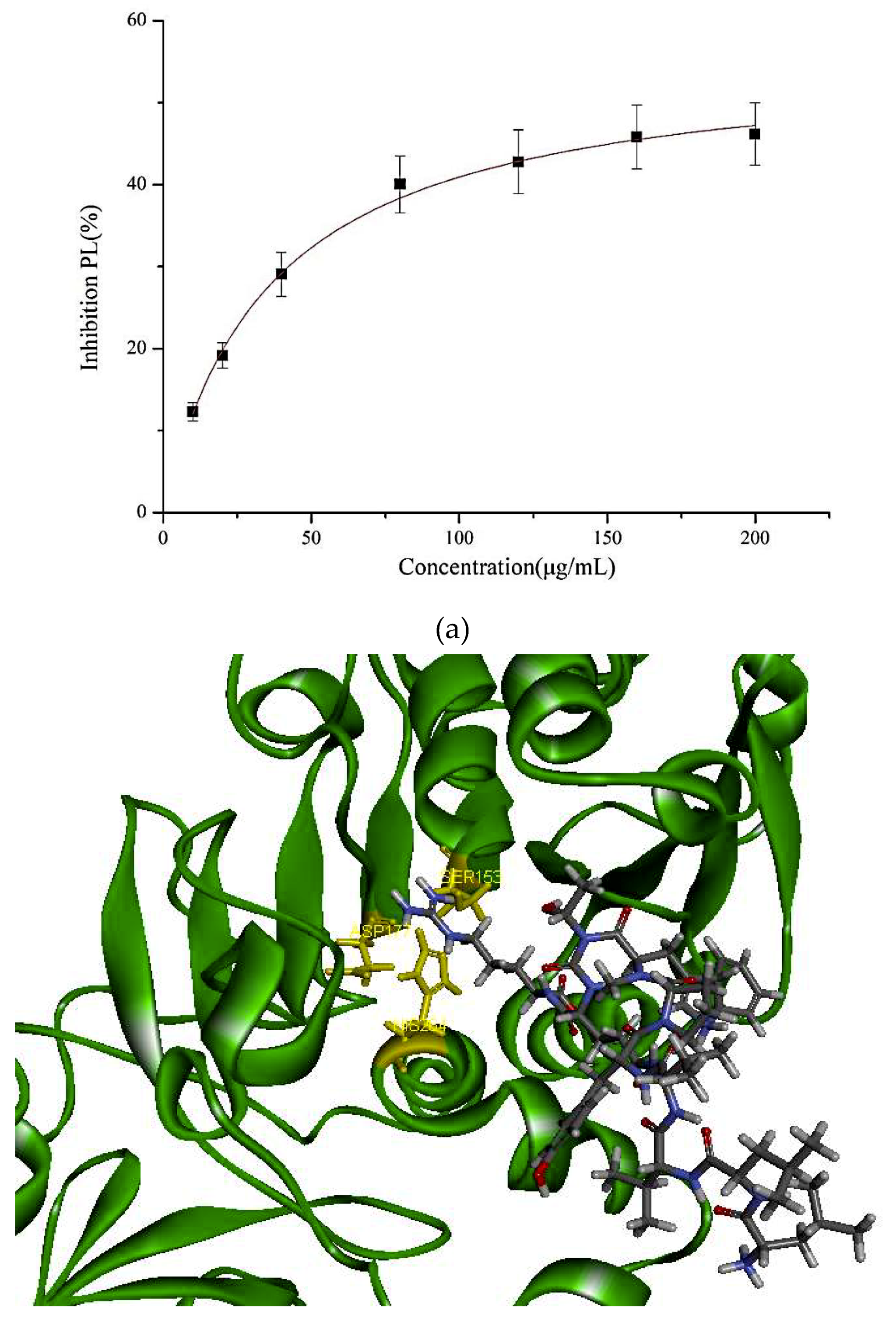
2.5. PL Inhibitory Activities of the Synthesized Peptide
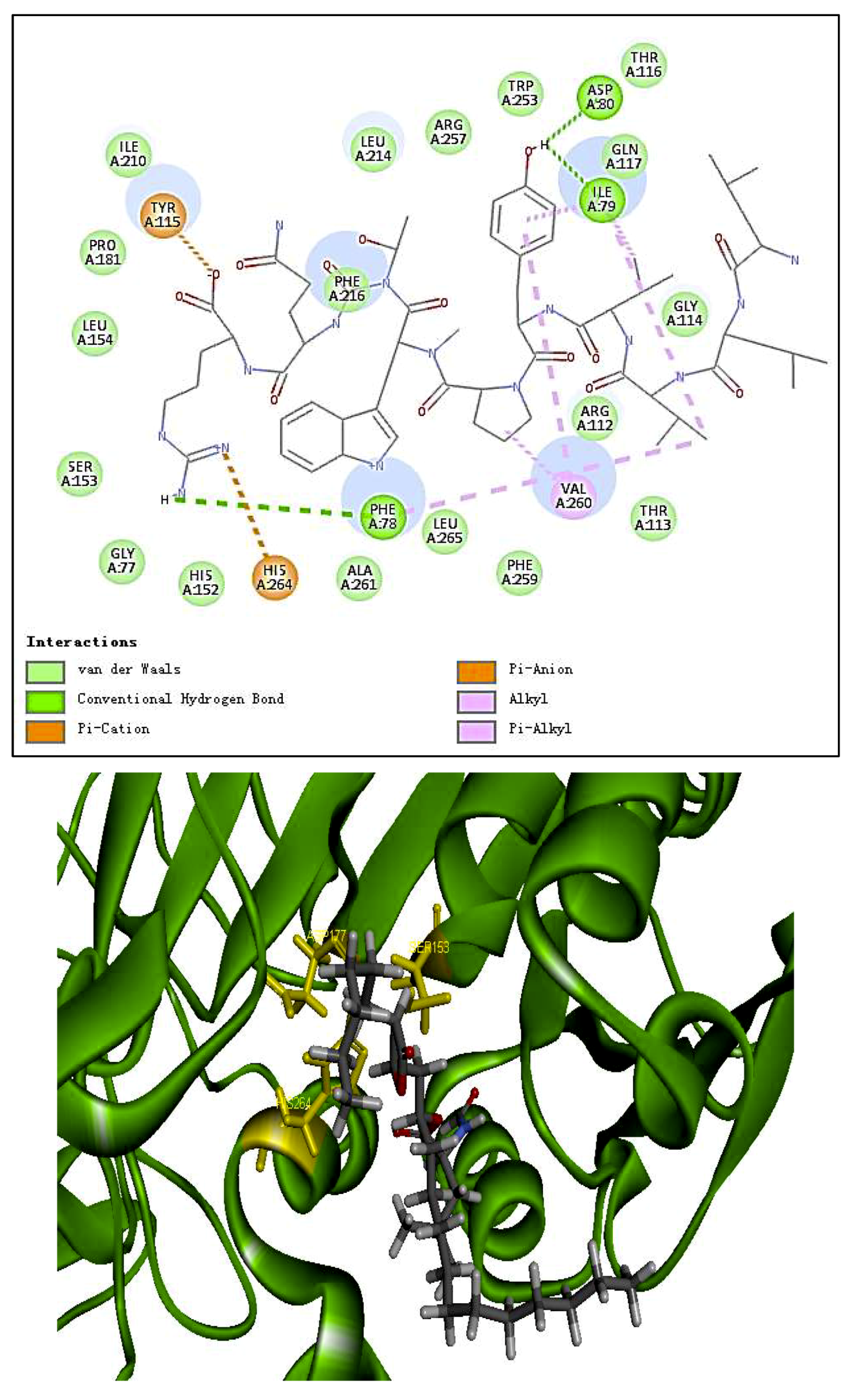
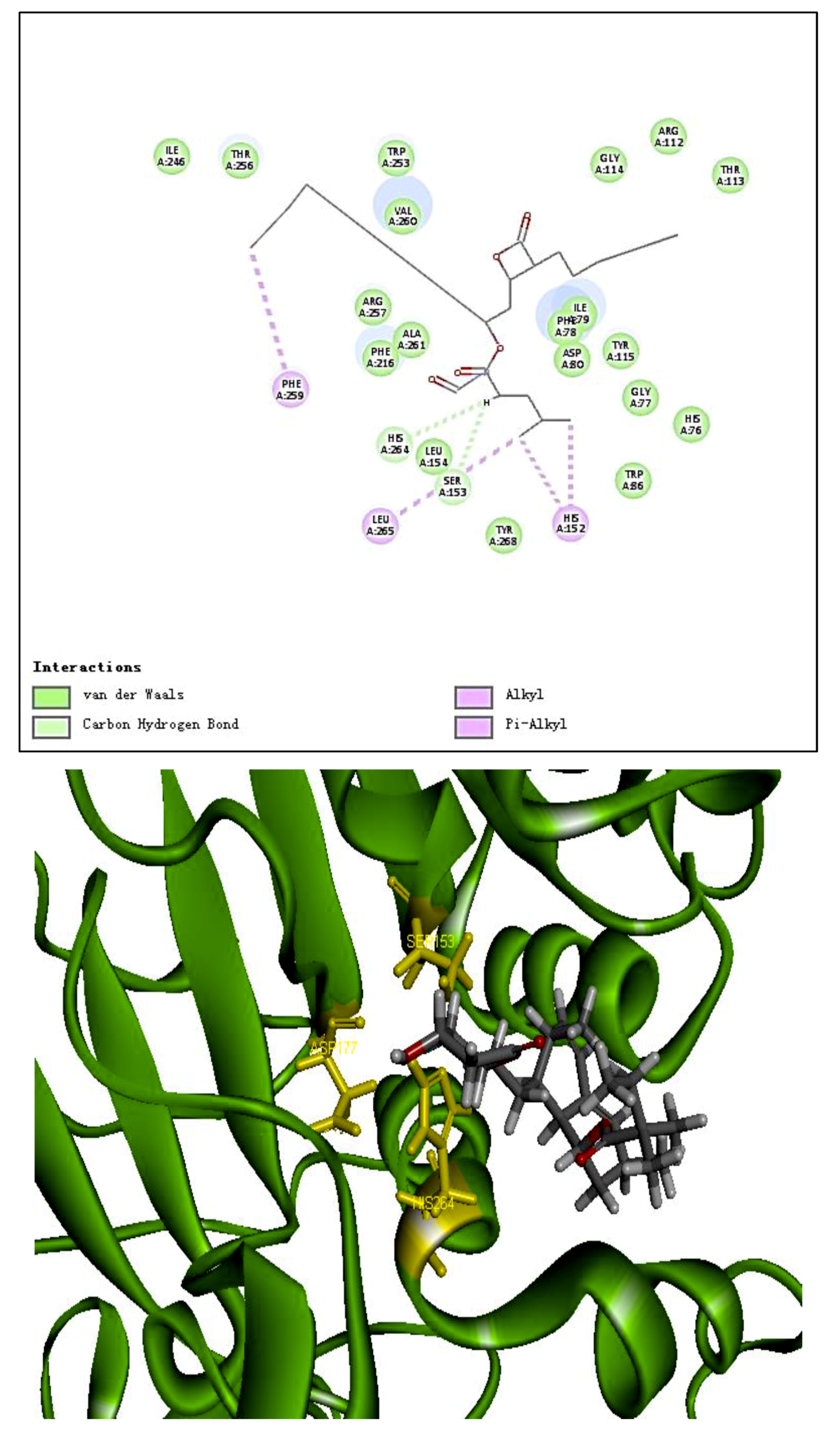
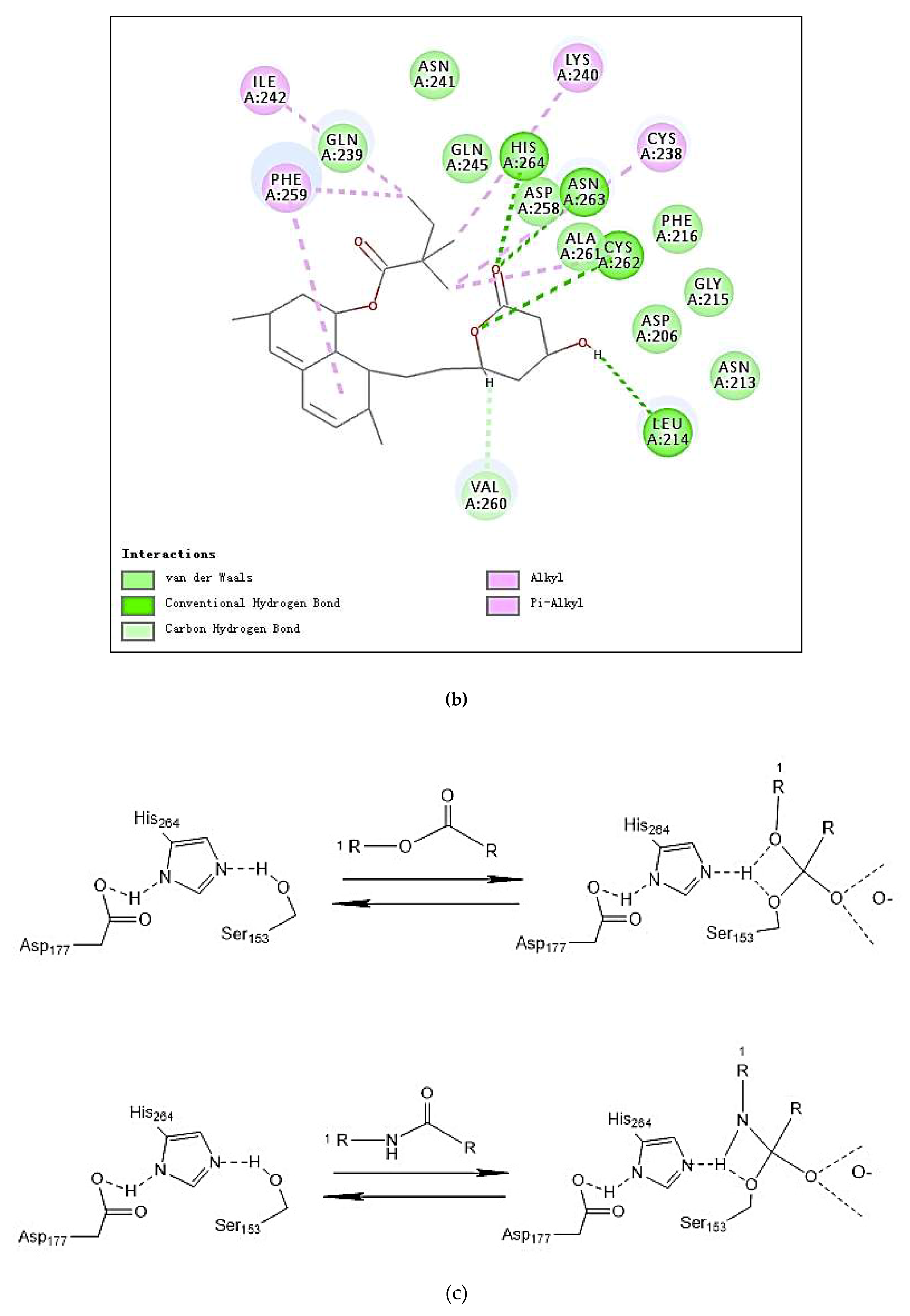
2.6. Docking Studies
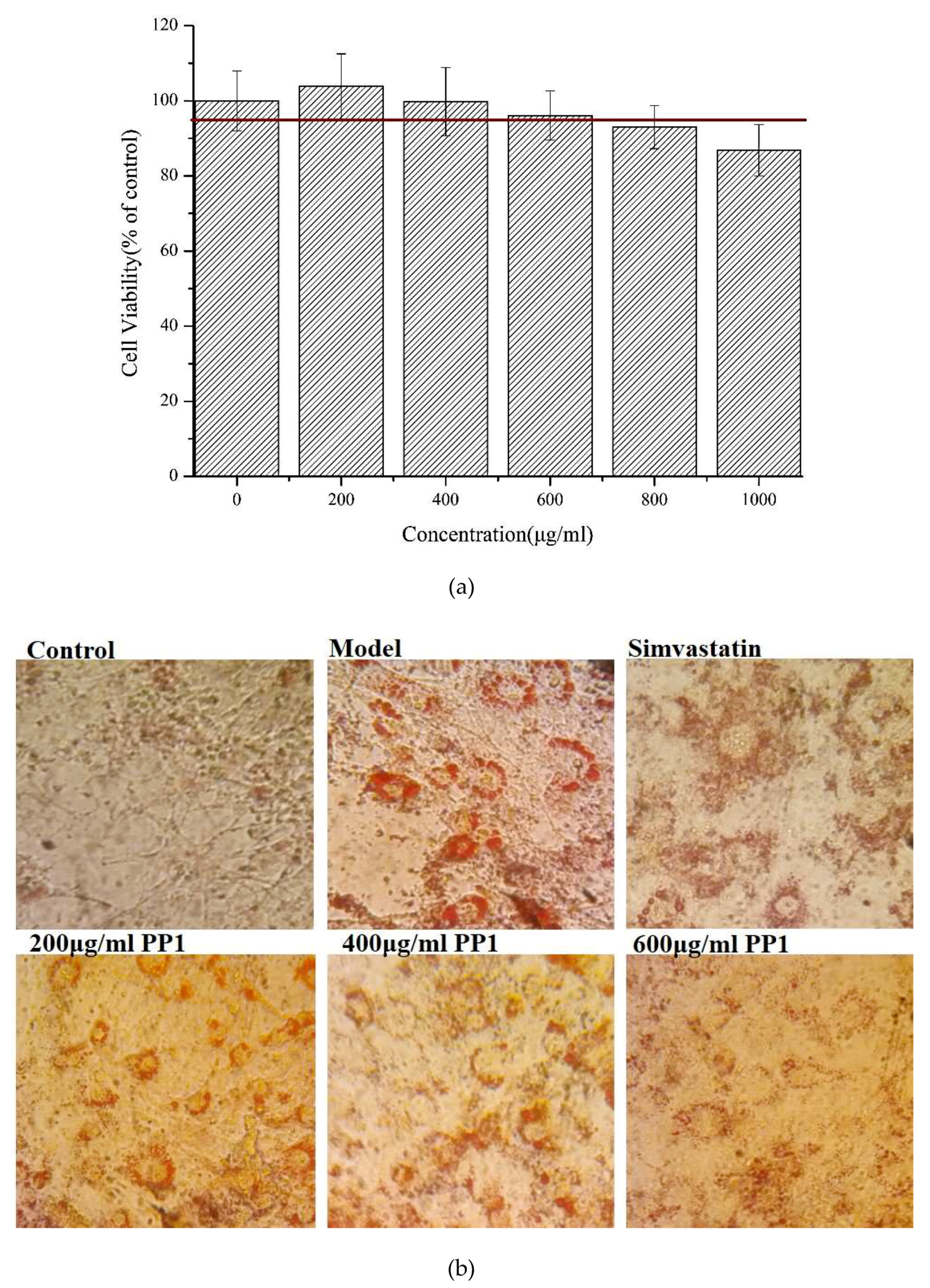
2.7. Effects of the Decapeptide PP1 on Lipid Accumulation in 3T3-L1 Cells
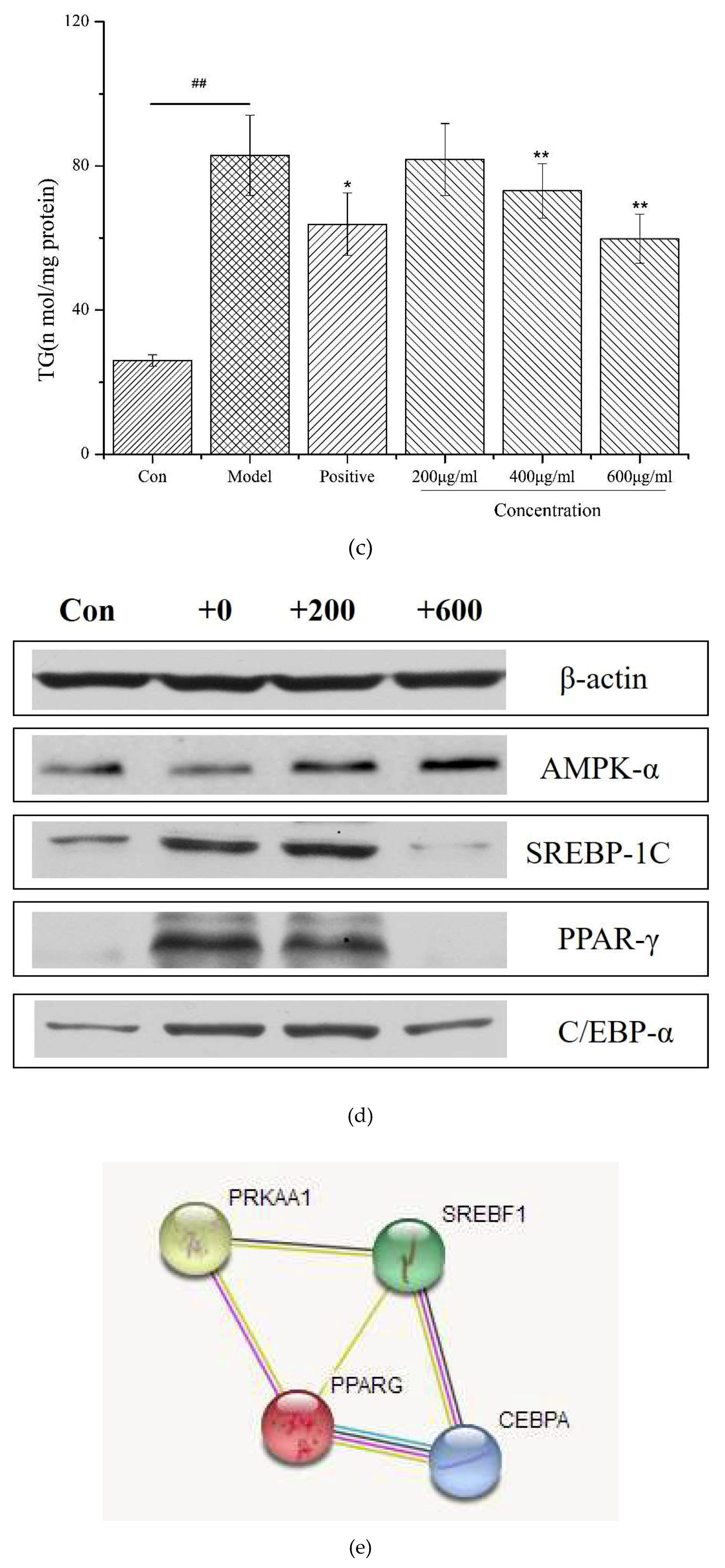
2.8. Effects of the Decapeptide PP1 on the Expression of Protein Levels in 3T3-L1 Cells
3. Discussion
4. Materials and Methods
4.1. Materials and Chemicals
4.2. Hydrolysis of Proteins
4.3. Molecular Weight Distributions
4.4. Separation by Ultrafiltration
4.5. Gel Filtration Chromatography
4.6. Analysis by LC–MS/MS
4.7. Peptide Synthesis
4.8. Lipase Activity Assay
4.9. Docking
4.10. Lipid Inhibition Assay in Cells
4.11. Western Blot Analysis
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Venuste, M.; Zhang, X.; Shoemaker, C.F.; Karangwa, E.; Abbas, S.; Kamdem, P.E. Influence of enzymatic hydrolysis and enzyme type on the nutritional and antioxidant properties of pumpkin meal hydrolysates. Food Funct. 2013, 4, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, X. Separation, antitumor activities, and encapsulation of polypeptide from Chlorella pyrenoidosa. Biotechnol. Prog. 2013, 29, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, X. Inhibitory effects of small molecular peptides from Spirulina (Arthrospira) platensis on cancer cell growth. Food Funct. 2016, 7, 781–788. [Google Scholar] [CrossRef] [PubMed]
- Moayedi, A.; Mora, L.; Aristoy, M.C.; Hashemi, M.; Safari, M.; Toldra, F. ACE-Inhibitory and Antioxidant Activities of Peptide Fragments Obtained from Tomato Processing By-Products Fermented Using Bacillus subtilis: Effect of Amino Acid Composition and Peptides Molecular Mass Distribution. Appl. Biochem. Biotechnol. 2017, 181, 48–64. [Google Scholar] [CrossRef] [PubMed]
- Chai, T.T.; Law, Y.C.; Wong, F.C.; Kim, S.K. Enzyme-Assisted Discovery of Antioxidant Peptides from Edible Marine Invertebrates: A Review. Mar. Drugs 2017, 15, 42. [Google Scholar] [CrossRef] [PubMed]
- Memarpoor-Yazdi, M.; Mahaki, H.; Zare-Zardini, H. Antioxidant activity of protein hydrolysates and purified peptides from Zizyphus jujuba fruits. J. Funct. Foods 2013, 5, 62–70. [Google Scholar] [CrossRef]
- Lee, S.H.; Kang, H.J.; Lee, H.J.; Kang, M.H.; Park, Y.K. Six-week supplementation with Chlorella has favorable impact on antioxidant status in Korean male smokers. Nutrition 2010, 26, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Ji, R.; Li, Y.; Zhang, J.; Chen, T. Study on immunoregulation of Chlorella pyrenoidos with disintegrated cell wall on mice. Chin J Public Health 2008, 7, 836–837. (In Chinese) [Google Scholar]
- Sheih, I.C.; Fang, T.J.; Wu, T.-K. Isolation and characterisation of a novel angiotensin I-converting enzyme (ACE) inhibitory peptide from the algae protein waste. Food Chem. 2009, 115, 279–284. [Google Scholar] [CrossRef]
- Mukherjee, M. Human digestive and metabolic lipases—A brief review. J. Mol. Catal. B: Enzym. 2003, 22, 369–376. [Google Scholar] [CrossRef]
- Birari, R.B.; Bhutani, K.K. Pancreatic lipase inhibitors from natural sources: Unexplored potential. Drug Discov. Today 2007, 12, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Grove, K.A.; Sae-tan, S.; Kennett, M.J.; Lambert, J.D. (-)-Epigallocatechin-3-gallate inhibits pancreatic lipase and reduces body weight gain in high fat-fed obese mice. Obesity 2012, 20, 2311–2313. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.W. Possible anti-obesity therapeutics from nature--a review. Phytochemistry 2010, 71, 1625–1641. [Google Scholar] [CrossRef]
- Shen, S.; Liao, Q.; Feng, Y.; Liu, J.; Pan, R.; Lee, S.M.; Lin, L. Myricanol mitigates lipid accumulation in 3T3-L1 adipocytes and high fat diet-fed zebrafish via activating AMP-activated protein kinase. Food Chem. 2019, 270, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.C.; Ding, Y.; Kim, H.S.; Jeon, Y.J.; Lee, S.H. Inhibition of Adipogenesis by Diphlorethohydroxycarmalol (DPHC) through AMPK Activation in Adipocytes. Mar. Drugs 2019, 17, 44. [Google Scholar] [CrossRef]
- Li, A.N.; Chen, J.J.; Li, Q.Q.; Zeng, G.Y.; Chen, Q.Y.; Chen, J.L.; Liao, Z.M.; Jin, P.; Wang, K.S.; Yang, Z.C. Alpha-glucosidase inhibitor 1-Deoxynojirimycin promotes beige remodeling of 3T3-L1 preadipocytes via activating AMPK. Biochem. Biophys. Res. Commun. 2019, 509, 1001–1007. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Chen, J.; Zhang, X. Extraction of intracellular protein from Chlorella pyrenoidosa using a combination of ethanol soaking, enzyme digest, ultrasonication and homogenization techniques. Bioresour. Technol. 2018, 247, 267–272. [Google Scholar] [CrossRef]
- Kim, Y.S.; Lee, Y.; Kim, J.; Sohn, E.; Kim, C.S.; Lee, Y.M.; Jo, K.; Shin, S.; Song, Y.; Kim, J.H.; et al. Inhibitory Activities of Cudrania tricuspidata Leaves on Pancreatic Lipase In Vitro and Lipolysis In Vivo. Evid. Based Complement Alternat. Med. 2012. [Google Scholar] [CrossRef]
- Lim, J.; Kale, M.; Kim, D.H.; Kim, H.S.; Chon, J.W.; Seo, K.H.; Lee, H.G.; Yokoyama, W.; Kim, H. Antiobesity Effect of Exopolysaccharides Isolated from Kefir Grains. J. Agric. Food Chem. 2017, 65, 10011–10019. [Google Scholar] [CrossRef]
- Scott, J.W.; Galic, S.; Graham, K.L.; Foitzik, R.; Ling, N.X.; Dite, T.A.; Issa, S.M.; Langendorf, C.G.; Weng, Q.P.; Thomas, H.E.; et al. Inhibition of AMP-Activated Protein Kinase at the Allosteric Drug-Binding Site Promotes Islet Insulin Release. Chem. Biol. 2015, 22, 705–711. [Google Scholar] [CrossRef] [Green Version]
- Sinkovits, A.F.; Bryksa, B.C.; Tanaka, T.; Yada, R.Y. Understanding the structure–function role of specific catalytic residues in a model food related enzyme: Pepsin. Enzym. Microb. Technol. 2007, 40, 1175–1180. [Google Scholar] [CrossRef]
- Wu, X.; He, W.; Zhang, H.; Li, Y.; Liu, Z.; He, Z. Acteoside: A lipase inhibitor from the Chinese tea Ligustrum purpurascens kudingcha. Food Chem. 2014, 142, 306–310. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MW (Da) | Percentage of Molecular Weight (%) | ||||
|---|---|---|---|---|---|
| Original Protein | Pepsin Hydrolysate | Papain Hydrolysate | Trypsin Hydrolysate | Alcalase Hydrolysate | |
| >10000 | 13.95 | 7.14 | 5.98 | 5.26 | 4.83 |
| 10000~5000 | 18.95 | 16.50 | 12.95 | 11. 45 | 9.95 |
| 5000~3000 | 27.38 | 36.34 | 31.87 | 33.37 | 32.51 |
| 3000~1000 | 29.29 | 33.02 | 35.65 | 35.82 | 38.63 |
| <1000 | 11.03 | 9.03 | 13.27 | 14.06 | 13.81 |
| Groups | Inhibition of Pancreatic Lipase (%) |
|---|---|
| >5 kDa | 41.4 ± 2.75 |
| <5 kDa | 75.73 ± 2.88 |
| A1 | 54.51 ± 1.93 |
| A2 | 74.05 ± 3.7 |
| A3 | 60.5 ± 1.89 |
| A4 | 63.5 ± 2.81 |
| Positive drug: simvastatin | 35.2 ± 1.9 |
| Positive drug: Orlistat | 88.85 ± 2.42 |
| NO. Peptide | Identified Peptide SEQUENCE | Observed | Mr(expt) | Mr(calc) | Protein in C. pyenoidose | Bioactivity Prediction Score |
|---|---|---|---|---|---|---|
| 1 | SISISVAGGGR | 531.28 | 1060.54 | 1059.56 | Fructose-bisphosphate aldolase 1 | 0.45 |
| 2 | LLVVYPWTQR | 637.31 | 1272.59 | 1273.71 | Ribosomal protein S, Chloroplastic | 0.38 |
| 3 | SDDPHTFGQGTK | 645.80 | 1289.61 | 1288.56 | Protein brassinosteroid insensitive | 0.35 |
| 4 | SRQLTLYPGAER | 695.46 | 1388.89 | 1388.73 | Transmembrane protein | 0.17 |
| 5 | KNGAPAEK | 408.23 | 814.44 | 813.43 | ADP-ribosylation factor GTPase-activating protein | 0.15 |
| 6 | KQTALVELVK | 377.18 | 1128.52 | 1127.69 | Ribosomal protein S2, Chloroplastic | 0.09 |
| Descriptor | LLVVYPWTQR | Orlistat | Simvastatin |
|---|---|---|---|
| Docking sore | 147.67 | 152.887 | 124.76 |
| Residues formed hydrogen bonds with the ligand | Ile79, Asp80, Tyr115, Ser153 | _ | Leu214, Cys262, Asn263, His264 |
| Pi interaction | 0 | 4 | 1 |
| Number of amino acids | 20 | 24 | 15 |
| Number of hydrogen bonds | 6 | 0 | 4 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, R.; Chen, J.; Mao, X.; Qi, P.; Zhang, X. Separation and Lipid Inhibition Effects of a Novel Decapeptide from Chlorella pyenoidose. Molecules 2019, 24, 3527. https://doi.org/10.3390/molecules24193527
Zhang R, Chen J, Mao X, Qi P, Zhang X. Separation and Lipid Inhibition Effects of a Novel Decapeptide from Chlorella pyenoidose. Molecules. 2019; 24(19):3527. https://doi.org/10.3390/molecules24193527
Chicago/Turabian StyleZhang, Ruilin, Jian Chen, Xinwu Mao, Ping Qi, and Xuewu Zhang. 2019. "Separation and Lipid Inhibition Effects of a Novel Decapeptide from Chlorella pyenoidose" Molecules 24, no. 19: 3527. https://doi.org/10.3390/molecules24193527
APA StyleZhang, R., Chen, J., Mao, X., Qi, P., & Zhang, X. (2019). Separation and Lipid Inhibition Effects of a Novel Decapeptide from Chlorella pyenoidose. Molecules, 24(19), 3527. https://doi.org/10.3390/molecules24193527