A New Approach of Extraction of α-Amylase/trypsin Inhibitors from Wheat (Triticum aestivum L.), Based on Optimization Using Plackett–Burman and Box–Behnken Designs

 ,
, 

Abstract
:1. Introduction
2. Results and Discussion
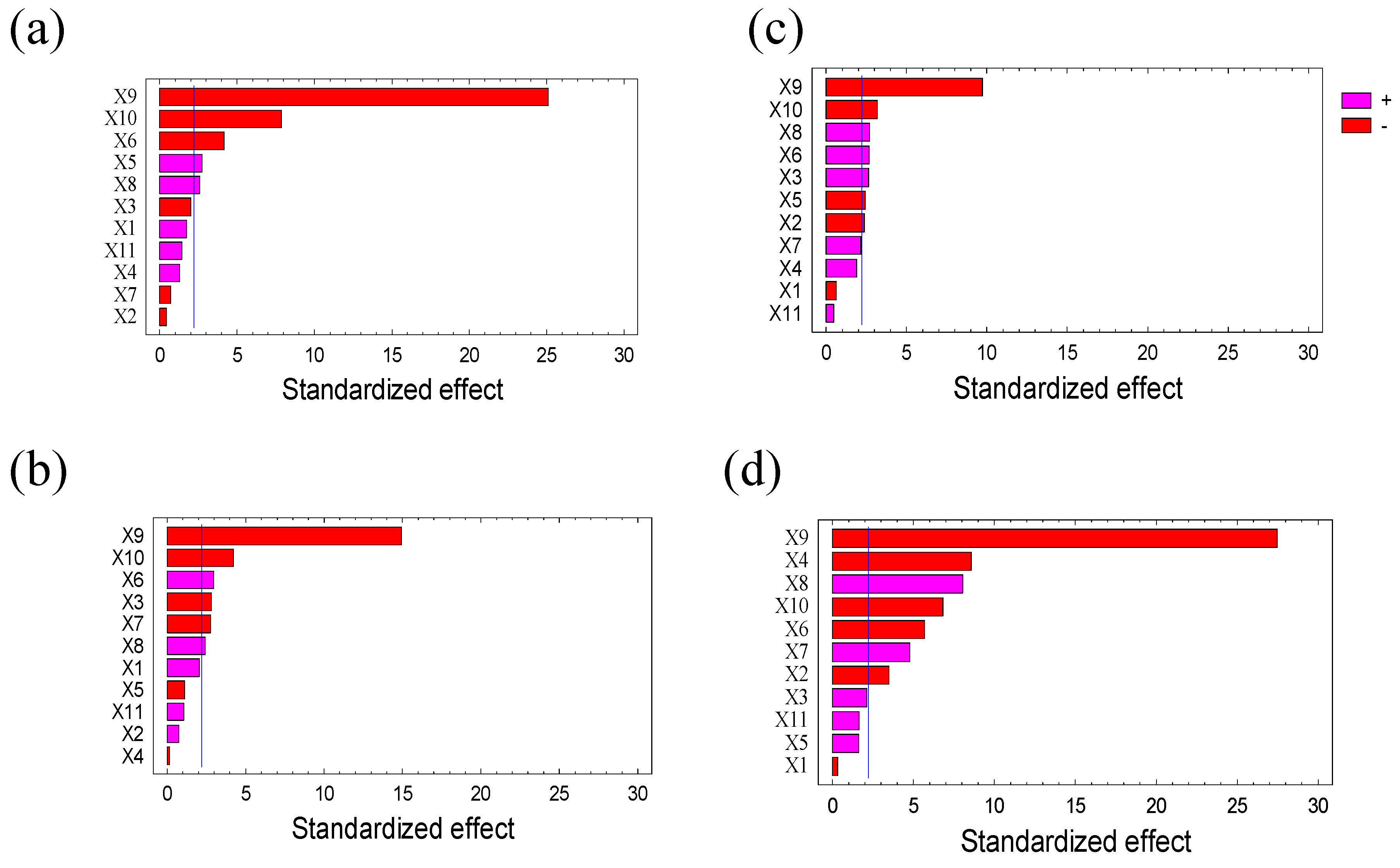
2.1. Screening with Plackett Burman Design
2.1.1. Evaluation of Parameters Effects on the Extraction of ATIs
2.1.2. Selection of Parameters for Optimization
2.2. Optimization of Extraction Process
2.2.1. Evaluation of the Effect of Parameters on Protein Concentration in IAA
2.2.2. Numerical Optimization
3. Materials and Methods
3.1. Materials
3.2. Extraction of ATIs
3.3. Experimental Designs
3.3.1. Plackett–Burman Design to Screen Parameters
3.3.2. Doehlert Design
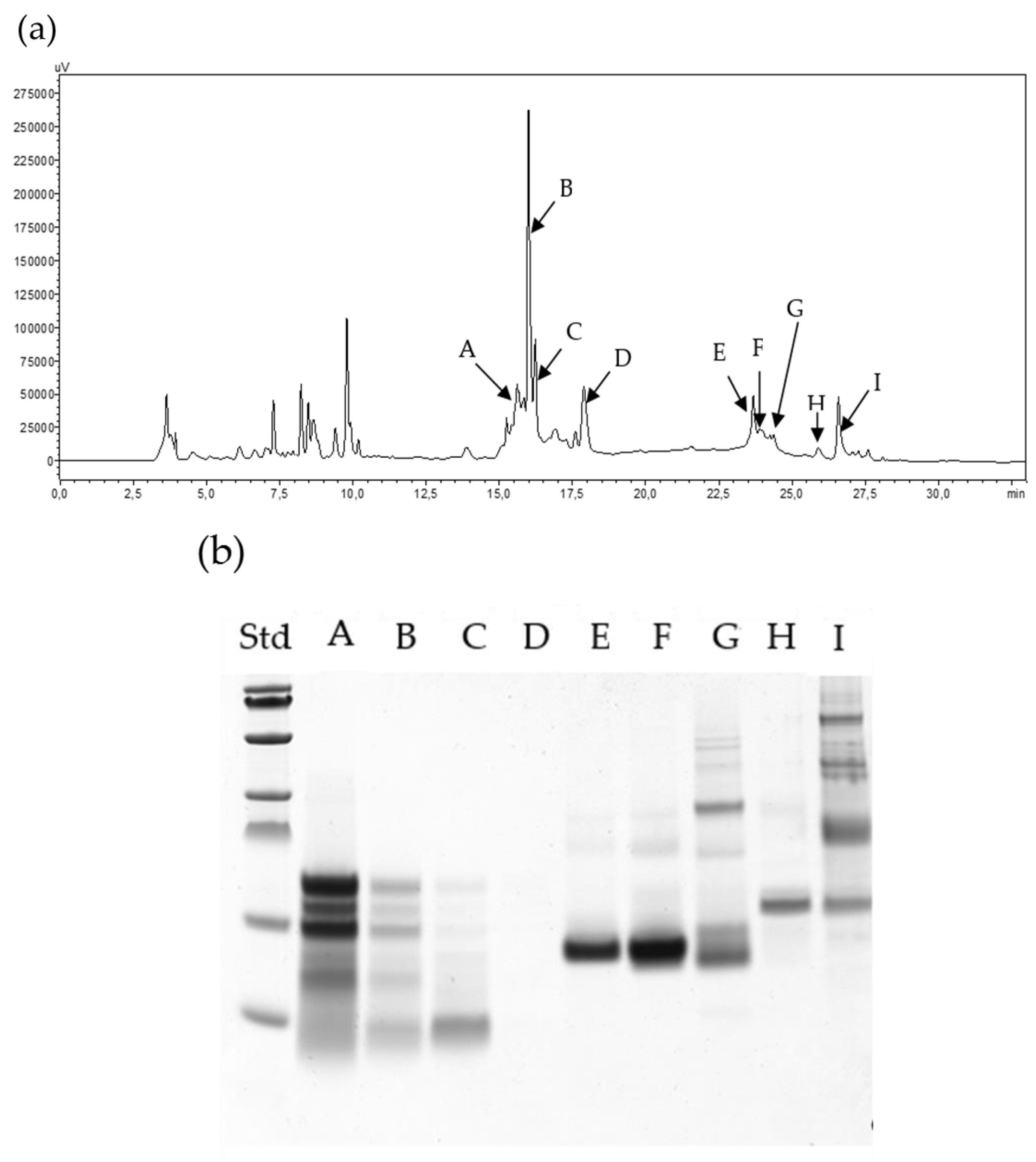
3.4. High-Performance Liquid Chromatography
3.5. Analyses
3.5.1. Determination of Protein Concentration
3.5.2. Inhibition of Amylase Activity
3.5.3. Trypsin Activity and Determination of it Inhibition
3.5.4. Electrophoresis
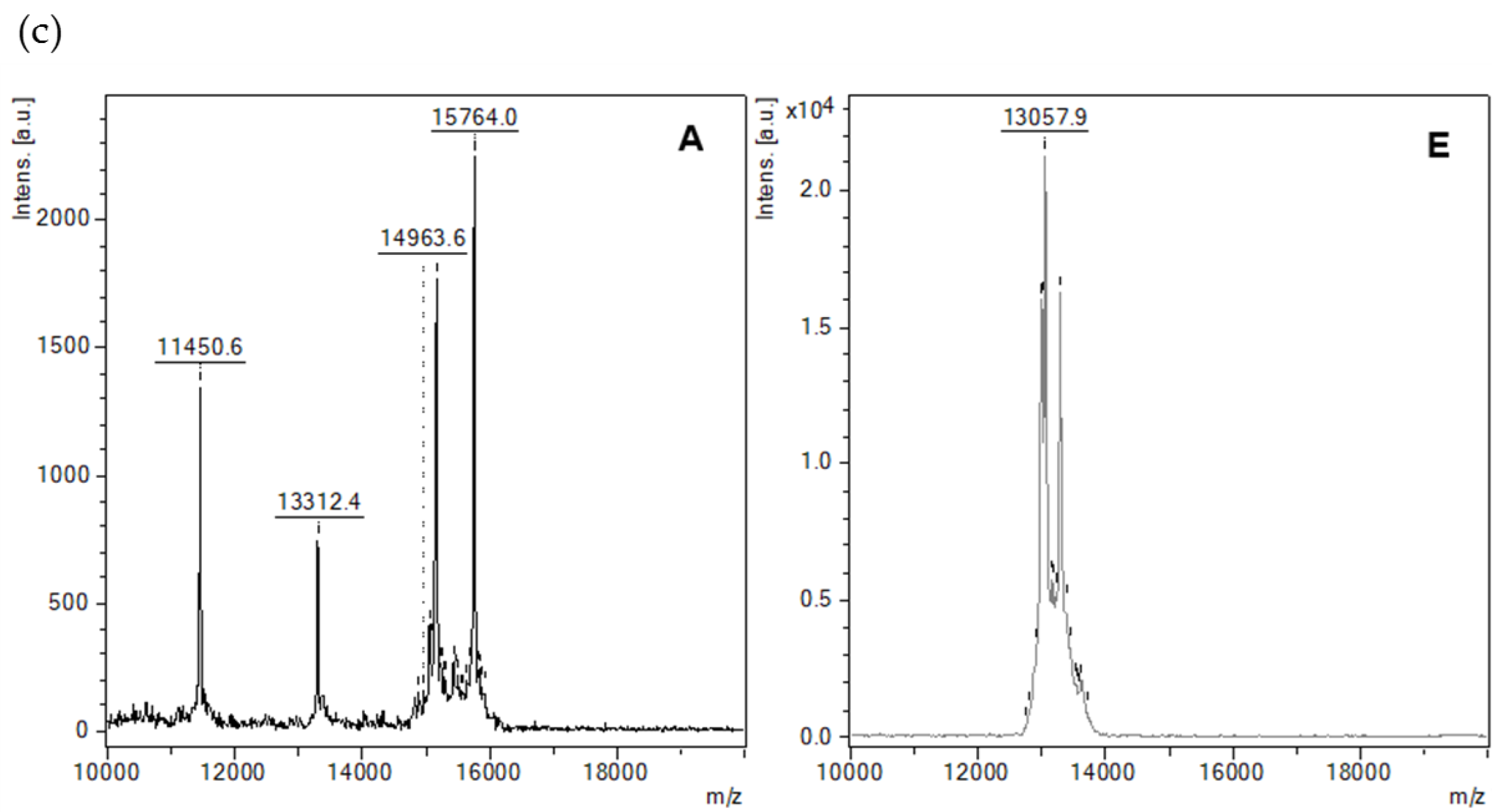
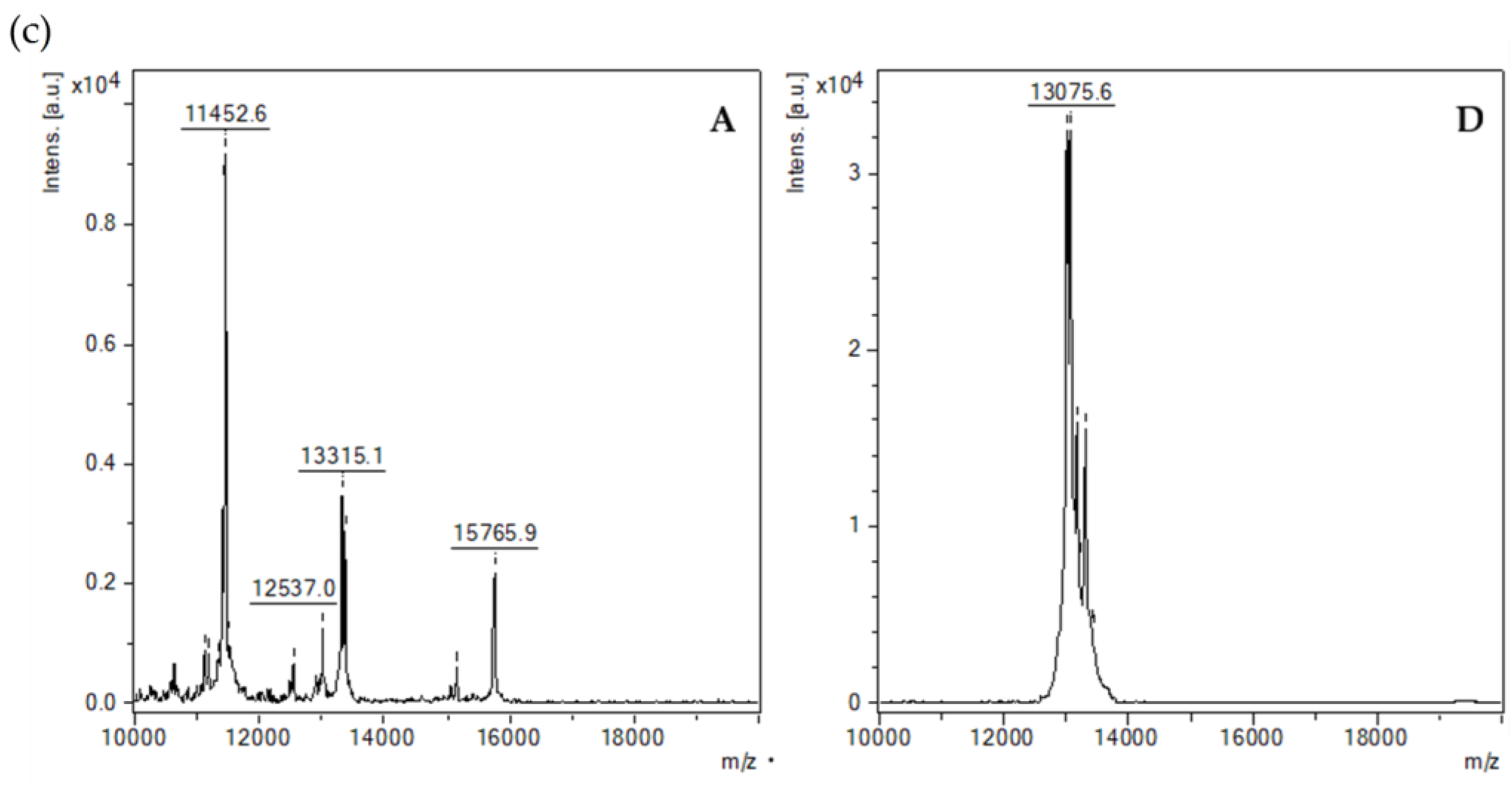
3.5.5. MALDI-TOF-MS Analysis
3.5.6. Protein Digestion and Untargeted Triple TOF/MS-Analysis
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACN | Acetonitrile |
| AF | Accuracy factor |
| AAC | Amylase activity of the control |
| AAT | Tested amylase activity |
| b0, b1, b2, … | Coefficients of models |
| ATIs | α-amylase/trypsin inhibitors |
| C/M | Chloroform/methanol mixture |
| D/M | Dichloromethane/methanol mixture |
| DD | Doehlert design |
| DNS | Dinitrosalicylic acid color reagent |
| DoE | Design of experiment |
| IAA | Inhibition of amylase activity |
| ITA | Inhibition of trypsin activity |
| LM | Linear mode |
| M | Molarity |
| MALDI-TOF | Matrix Assisted Laser Desorption Ionization-Time of Flight |
| MS | Mass Spectrometry |
| N | Number of parameters |
| p | Probability |
| PBD | Plackett - Burman design |
| SDS | Sodium dodecyl sulfate |
| X1,X2,X3 | Parameters |
| TAC | Trypsin activity of control |
| TAT | Tested trypsin activity |
| TFA | Trifluoroacetic |
| Y | Experimental response |
| 2,5-DHAP | 2,5-Dihydroxy actetophenone |
References
- Makela, M.J.; Eriksson, C.; Kotaniemi-Syrjanen, A.; Palosuo, K.; Marsh, J.; Borres, M.; Kuitunen, M.; Pelkonen, A.S. Wheat allergy in children-new tools for diagnostics. Clin. Exp. Allergy 2014, 44, 1420–1430. [Google Scholar] [CrossRef] [PubMed]
- Young, E.; Stoneham, M.D.; Petruckevitch, A.; Barton, J.; Rona, R. A population study of food intolerance. Lancet 1994, 343, 1127–1130. [Google Scholar] [CrossRef]
- Jin, Y.; Acharya, H.G.; Acharya, D.; Jorgensen, R.; Gao, H.; Secord, J.; Ng, P.K.W.; Gangur, V. Advances in Molecular Mechanisms of Wheat Allergenicity in Animal Models: A Comprehensive Review. Molecules 2019, 24, 1142. [Google Scholar] [CrossRef] [PubMed]
- Verhoeckx, K.C.M.; Vissers, Y.M.; Baumert, J.L.; Faludi, R.; Feys, M.; Flanagan, S.; Herouet-Guicheney, C.; Holzhauser, T.; Shimojo, R.; van der Bolt, N.; et al. Food processing and allergenicity. Food Chem. Toxicol. 2015, 80, 223–240. [Google Scholar] [CrossRef] [PubMed]
- Berin, M.C.; Sampson, H.A. Mucosal immunology of food allergy. Curr. Biol. 2013, 23, R389–R400. [Google Scholar] [CrossRef]
- Bathrellou, E.; Kontogianni, M.D.; Panagiotakos, D.B. Celiac disease and non-celiac gluten or wheat sensitivity and health in later life: A review. Maturitas 2018, 112, 29–33. [Google Scholar] [CrossRef]
- Talley, N.J.; Walker, M.M. Celiac Disease and Nonceliac Gluten or Wheat Sensitivity: The Risks and Benefits of Diagnosis. JAMA Intern. Med. 2017, 177, 615–616. [Google Scholar] [CrossRef]
- Catassi, C.; Bai, J.C.; Bonaz, B.; Bouma, G.; Calabro, A.; Carroccio, A.; Castillejo, G.; Ciacci, C.; Cristofori, F.; Dolinsek, J.; et al. Non-Celiac Gluten sensitivity: The new frontier of gluten related disorders. Nutrients 2013, 5, 3839–3853. [Google Scholar] [CrossRef]
- Jang, Y.R.; Beom, H.R.; Altenbach, S.B.; Lee, M.K.; Lim, S.H.; Lee, J.Y. Improved Method for Reliable HMW-GS Identification by RP-HPLC and SDS-PAGE in Common Wheat Cultivars. Molecules 2017, 22, 1055. [Google Scholar] [CrossRef]
- Franco, O.L.; Rigden, D.J.; Melo, F.R.; Bloch, C., Jr.; Silva, C.P.; Grossi de Sa, M.F. Activity of wheat alpha-amylase inhibitors towards bruchid alpha-amylases and structural explanation of observed specificities. Eur. J. Biochem. 2000, 267, 2166–2173. [Google Scholar] [CrossRef]
- Oda, Y.; Matsunaga, T.; Fukuyama, K.; Miyazaki, T.; Morimoto, T. Tertiary and quaternary structures of 0.19 alpha-amylase inhibitor from wheat kernel determined by X-ray analysis at 2.06 A resolution. Biochemistry 1997, 36, 13503–13511. [Google Scholar] [CrossRef] [PubMed]
- Feng, G.H.; Richardson, M.; Chen, M.S.; Kramer, K.J.; Morgan, T.D.; Reeck, G.R. alpha-Amylase inhibitors from wheat: Amino acid sequences and patterns of inhibition of insect and human alpha-amylases. Insect. Biochem. Mol. Biol 1996, 26, 419–426. [Google Scholar] [CrossRef]
- Prandi, B.; Faccini, A.; Tedeschi, T.; Galaverna, G.; Sforza, S. LC/MS analysis of proteolytic peptides in wheat extracts for determining the content of the allergen amylase/trypsin inhibitor CM3: Influence of growing area and variety. Food Chem. 2013, 140, 141–146. [Google Scholar] [CrossRef]
- Zevallos, V.F.; Raker, V.; Tenzer, S.; Jimenez-Calvente, C.; Ashfaq-Khan, M.; Russel, N.; Pickert, G.; Schild, H.; Steinbrink, K.; Schuppan, D. Nutritional Wheat Amylase-Trypsin Inhibitors Promote Intestinal Inflammation via Activation of Myeloid Cells. Gastroenterology 2017, 152, 1100–1113.e1112. [Google Scholar] [CrossRef]
- Alves, T.O.; D’Almeida, C.T.S.; Victorio, V.C.M.; Souza, G.H.M.F.; Cameron, L.C.; Ferreira, M.S.L. Immunogenic and allergenic profile of wheat flours from different technological qualities revealed by ion mobility mass spectrometry. J. Food Comp. Analy. 2018, 73, 67–75. [Google Scholar] [CrossRef]
- Sotkovsky, P.; Sklenar, J.; Halada, P.; Cinova, J.; Setinova, I.; Kainarova, A.; Golias, J.; Pavlaskova, K.; Honzova, S.; Tuckova, L. A new approach to the isolation and characterization of wheat flour allergens. Clin. Exp. Allergy 2011, 41, 1031–1043. [Google Scholar] [CrossRef]
- Altenbach, S.B.; Vensel, W.H.; Dupont, F.M. The spectrum of low molecular weight alpha-amylase/protease inhibitor genes expressed in the US bread wheat cultivar Butte 86. BMC Res. Notes 2011, 4, 242. [Google Scholar] [CrossRef]
- Gomez, L.; Martin, E.; Hernandez, D.; Sanchez-Monge, R.; Barber, D.; del Pozo, V.; de Andres, B.; Armentia, A.; Lahoz, C.; Salcedo, G.; et al. Members of the alpha-amylase inhibitors family from wheat endosperm are major allergens associated with baker’s asthma. FEBS Lett. 1990, 261, 85–88. [Google Scholar] [CrossRef]
- Geisslitz, S.; Ludwig, C.; Scherf, K.A.; Koehler, P. Targeted LC-MS/MS Reveals Similar Contents of alpha-Amylase/Trypsin-Inhibitors as Putative Triggers of Nonceliac Gluten Sensitivity in All Wheat Species except Einkorn. J. Agric. Food Chem. 2018, 66, 12395–12403. [Google Scholar] [CrossRef]
- Tundo, S.; Lupi, R.; Lafond, M.; Giardina, T.; Larre, C.; Denery-Papini, S.; Morisset, M.; Kalunke, R.; Sestili, F.; Masci, S. Wheat ATI CM3, CM16 and 0.28 Allergens Produced in Pichia Pastoris Display a Different Eliciting Potential in Food Allergy to Wheat (double dagger). Plants 2018, 7, 101. [Google Scholar] [CrossRef]
- Moller, J.; Schroer, M.A.; Erlkamp, M.; Grobelny, S.; Paulus, M.; Tiemeyer, S.; Wirkert, F.J.; Tolan, M.; Winter, R. The effect of ionic strength, temperature, and pressure on the interaction potential of dense protein solutions: From nonlinear pressure response to protein crystallization. Biophys. J. 2012, 102, 2641–2648. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.R.; Mount, E.M.; Giles, H.F. Design of Experiments. In Extrusion (Second Edition); Wagner, J.R., Mount, E.M., Giles, H.F., Eds.; William Andrew Publishing: Oxford, UK, 2014; pp. 291–308. [Google Scholar]
- Drouillard, K.G.; Hagen, H.; Haffner, G.D. Evaluation of chloroform/methanol and dichloromethane/hexane extractable lipids as surrogate measures of sample partition capacity for organochlorines in fish tissues. Chemosphere 2004, 55, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Cequier-Sanchez, E.; Rodriguez, C.; Ravelo, A.G.; Zarate, R. Dichloromethane as a solvent for lipid extraction and assessment of lipid classes and fatty acids from samples of different natures. J. Agric. Food Chem. 2008, 56, 4297–4303. [Google Scholar] [CrossRef] [PubMed]
- Vertommen, A.; Panis, B.; Swennen, R.; Carpentier, S.C. Evaluation of chloroform/methanol extraction to facilitate the study of membrane proteins of non-model plants. Planta 2010, 231, 1113–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salcedo, G.; Rodriguezloperena, M.A.; Aragoncillo, C. Relationships among Low Mw Hydrophobic Proteins from Wheat Endosperm. Phytochemistry 1978, 17, 1491–1494. [Google Scholar] [CrossRef]
- Guinn, E.J.; Pegram, L.M.; Capp, M.W.; Pollock, M.N.; Record, M.T., Jr. Quantifying why urea is a protein denaturant, whereas glycine betaine is a protein stabilizer. Proc. Natl. Acad. Sci. USA 2011, 108, 16932–16937. [Google Scholar] [CrossRef] [Green Version]
- Felitsky, D.J.; Record, M.T., Jr. Thermal and urea-induced unfolding of the marginally stable lac repressor DNA-binding domain: A model system for analysis of solute effects on protein processes. Biochemistry 2003, 42, 2202–2217. [Google Scholar] [CrossRef]
- Whitney, P.L.; Tanford, C. Solubility of amino acids in aqueous urea solutions and its implications for the denaturation of proteins by urea. J. Biol. Chem. 1962, 237, 1735–1737. [Google Scholar]
- Leberman, R. The Hofmeister series and ionic strength. FEBS Lett. 1991, 284, 293–294. [Google Scholar] [CrossRef] [Green Version]
- Calzone, F.J.; Angerer, R.C.; Gorovsky, M.A. Regulation of protein synthesis in Tetrahymena: Isolation and characterization of polysomes by gel filtration and precipitation at pH 5.3. Nucl. Acids Res. 1982, 10, 2145–2161. [Google Scholar] [CrossRef]
- Maity, H.; Karkaria, C.; Davagnino, J. Mapping of solution components, pH changes, protein stability and the elimination of protein precipitation during freeze-thawing of fibroblast growth factor 20. Int. J. Pharm. 2009, 378, 122–135. [Google Scholar] [CrossRef] [PubMed]
- Henning, A.K.; Albrecht, D.; Riedel, K.; Mettenleiter, T.C.; Karger, A. An alternative method for serum protein depletion/enrichment by precipitation at mildly acidic pH values and low ionic strength. Proteomics 2015, 15, 1935–1940. [Google Scholar] [CrossRef] [PubMed]
- Dexter, J.E.; Matsuo, R.R. Influence of Protein Content on Some Durum Wheat Quality Parameters. Canadian J. Plant Sci. 1977, 57, 717–727. [Google Scholar] [CrossRef]
- Terman, G.L. Yields and Protein Content of Wheat Grain as Affected by Cultivar, N, and Environmental Growth Factors1. Agronomy J. 1979, 71, 437. [Google Scholar] [CrossRef]
- Plackett, R.L.; Burman, J.P. The Design of Optimum Multifactorial Experiments. Biometrika 1946, 33, 305–325. [Google Scholar] [CrossRef]
- Pinkowska, H.; Krzywonos, M.; Wolak, P.; Zlocinska, A. Pectin and Neutral Monosaccharides Production during the Simultaneous Hydrothermal Extraction of Waste Biomass from Refining of Sugar-Optimization with the Use of Doehlert Design. Molecules 2019, 24, 472. [Google Scholar] [CrossRef] [PubMed]
- Fauduet, H.; Porte, C.; Havet, J.L.; Daguet, D. Modelling of influential parameters on a continuous evaporation process by Doehlert shells. J. Autom. Methods Manag. Chem. 2003, 25, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Bernfeld, P. Amylases, α and β. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1955; Volume 1, pp. 149–158. [Google Scholar]
- Sagu, S.T.; Nso, E.J.; Homann, T.; Kapseu, C.; Rawel, H.M. Extraction and purification of beta-amylase from stems of Abrus precatorius by three phase partitioning. Food Chem. 2015, 183, 144–153. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Ali, M.; Homann, T.; Khalil, M.; Kruse, H.P.; Rawel, H. Milk whey protein modification by coffee-specific phenolics: Effect on structural and functional properties. J. Agric. Food Chem. 2013, 61, 6911–6920. [Google Scholar] [CrossRef] [PubMed]
- Huschek, G.; Bonick, J.; Merkel, D.; Huschek, D.; Rawel, H. Authentication of leguminous-based products by targeted biomarkers using high resolution time of flight mass spectrometry. Lwt-Food Sci. Technol. 2018, 90, 164–171. [Google Scholar] [CrossRef]
- Bonick, J.; Huschek, G.; Rawel, H.M. Determination of wheat, rye and spelt authenticity in bread by targeted peptide biomarkers. J. Food Comp. Analy. 2017, 58, 82–91. [Google Scholar] [CrossRef]
Sample Availability: Samples are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nb | Parameters | Experimental Responses | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Julius | Ponticus | ||||||||||||||
| X1 | X2 | X3 | X4 | X5 | X6 | X7 | X8 | X9 | X10 | X11 | Protein conc. (mg/mL) | IAA (%) | Protein conc. (mg/mL) | IAA (%) | |
| 1 | D/M | 2:1 | 125:1 | 50 | 4 | 60 | 30 | 9 | (NH4)2SO4 | 1 | 10000 | 0.22 ± 0.03 | 29.3 ± 1.6 | 0.27 ± 0.03 | 26.6 ± 1.6 |
| 2 | D/M | 1:1 | 250:1 | 0 | 4 | 60 | 80 | 9 | (NH4)2SO4 | 0.5 | 12000 | 0.31 ± 0.01 | 63.3 ± 2.2 | 0.27 ± 0.01 | 65.0 ± 3.0 |
| 3 | C/M | 2:1 | 250:1 | 50 | 4 | 180 | 80 | 8 | (NH4)2SO4 | 0.5 | 10000 | 0.20 ± 0.04 | 61.9 ± 5.3 | 0.24 ± 0.08 | 29.0 ± 1.4 |
| 4 | D/M | 2:1 | 250:1 | 0 | 25 | 180 | 30 | 9 | NaCl | 0.5 | 10000 | 0.63 ± 0.04 | 87.8 ± 3.2 | 0.61 ± 0.04 | 90.3 ± 2.7 |
| 5 | D/M | 1:1 | 250:1 | 50 | 4 | 180 | 30 | 8 | NaCl | 1 | 12000 | 0.49 ± 0.01 | 90.2 ± 0.7 | 0.52 ± 0.01 | 62.0 ± 2.6 |
| 6 | C/M | 1:1 | 250:1 | 50 | 25 | 60 | 80 | 9 | NaCl | 1 | 10000 | 0.56 ± 0.05 | 89.8 ± 2.4 | 0.40 ± 0.03 | 87.0 ± 3.5 |
| 7 | C/M | 2:1 | 250:1 | 0 | 25 | 60 | 30 | 8 | (NH4)2SO4 | 1 | 12000 | 0.18 ± 0.01 | 17.6 ± 0.9 | 0.16 ± 0.03 | 35.7 ± 1.0 |
| 8 | C/M | 1:1 | 125:1 | 50 | 25 | 180 | 30 | 9 | (NH4)2SO4 | 0.5 | 12000 | 0.32 ± 0.01 | 56.0 ± 1.7 | 0.35 ± 0.03 | 39.5 ± 0.9 |
| 9 | C/M | 2:1 | 125:1 | 0 | 4 | 180 | 80 | 9 | NaCl | 1 | 12000 | 0.50 ± 0.02 | 85.1 ± 5.2 | 0.53 ± 0.05 | 83.4 ± 1.7 |
| 10 | D/M | 1:1 | 125:1 | 0 | 25 | 180 | 80 | 8 | (NH4)2SO4 | 1 | 10000 | 0.16 ± 0.01 | 29.8 ± 4.1 | 0.21 ± 0.02 | 33.8 ± 1.9 |
| 11 | C/M | 1:1 | 125:1 | 0 | 4 | 60 | 30 | 8 | NaCl | 0.5 | 10000 | 0.62 ± 0.07 | 78.7 ± 2.5 | 0.55 ± 0.01 | 87.3 ± 2.7 |
| 12 | D/M | 2:1 | 125:1 | 50 | 25 | 60 | 80 | 8 | (NH4)2SO4 | 1 | 12000 | 0.23 ± 0.02 | 20.4 ± 0.8 | 0.19 ± 0.01 | 26.6 ± 1.4 |
| Nb | Experimental Responses | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Coded Values | Real Values | Julius | Ponticus | |||||||||
| X10 | X6 | X8 | Protein Concentration | IAA | ITA | Protein Concentration | IAA | ITA | ||||
| x10 | x6 | x8 | (M) | (min) | (pH) | (mg/mL) | (%) | (%) | (mg/mL) | (%) | (%) | |
| 1 | 0 | 0 | 0 | 1 | 105 | 8 | 0.50 ± 0.05 | 73.1 ± 0.5 | 1.6 ± 0.6 | 0.48 ± 0.09 | 60.4 ± 3.1 | 3.8 ± 0.1 |
| 2 | 1 | 0 | 0 | 1.8 | 105 | 8 | 0.41 ± 0.05 | 82.2 ± 4.7 | 2.0 ± 0.1 | 0.32 ± 0.02 | 44.5 ± 2.3 | 2.1 ± 0.1 |
| 3 | −1 | 0 | 0 | 0.2 | 105 | 8 | 0.58 ± 0.02 | 83.7 ± 1.2 | 3.3 ± 0.2 | 0.37 ± 0.05 | 76.3 ± 1.0 | 3.5 ± 0.3 |
| 4 | 0.5 | 0.866 | 0 | 1.4 | 180 | 8 | 0.38 ± 0.03 | 81.0 ± 1.5 | 3.0 ± 0.5 | 0.65 ± 0.05 | 48.8 ± 0.8 | 5.8 ± 1.0 |
| 5 | −0.5 | −0.866 | 0 | 0.6 | 30 | 8 | 0.49 ± 0.01 | 83.7 ± 3.6 | 1.8 ± 0.0 | 0.36 ± 0.13 | 49.1 ± 0.7 | 6.6 ± 0.9 |
| 6 | 0.5 | −0.866 | 0 | 1.4 | 30 | 8 | 0.37 ± 0.02 | 84.2 ± 3.0 | 4.7 ± 0.9 | 0.77 ± 0.15 | 40.3 ± 1.1 | 7.1 ± 1.1 |
| 7 | −0.5 | 0.866 | 0 | 0.6 | 180 | 8 | 0.53 ± 0.03 | 83.3 ± 0.9 | 2.2 ± 0.3 | 0.66 ± 0.16 | 60.9 ± 0.1 | 5.3 ± 0.8 |
| 8 | 0.5 | 0.289 | 0.816 | 1.4 | 130 | 9 | 0.47 ± 0.04 | 75.0 ± 1.0 | 2.3 ± 0.9 | 0.49 ± 0.02 | 37.1 ± 1.5 | 6.4 ± 0.5 |
| 9 | −0.5 | −0.289 | −0.816 | 0.6 | 80 | 7 | 0.61 ± 0.03 | 80.9 ± 2.7 | 4.0 ± 0.2 | 0.79 ± 0.18 | 72.3 ± 1.7 | 5.7 ± 0.6 |
| 10 | 0.5 | −0.289 | −0.816 | 1.4 | 80 | 7 | 0.36 ± 0.03 | 85.3 ± 2.5 | 5.0 ± 0.3 | 0.34 ± 0.15 | 67.1 ± 1.1 | 3.5 ± 0.2 |
| 11 | 0 | 0.577 | −0.816 | 1 | 155 | 7 | 0.52 ± 0.02 | 77.5 ± 0.7 | 4.3 ± 0.7 | 0.40 ± 0.09 | 69.4 ± 1.2 | 4.1 ± 0.2 |
| 12 | −0.5 | 0.289 | 0.816 | 0.6 | 130 | 9 | 0.68 ± 0.02 | 82.5 ± 1.4 | 1.2 ± 0.1 | 0.58 ± 0.05 | 57.8 ± 2.1 | 3.6 ± 0.5 |
| 13 | 0 | −0.577 | 0.816 | 1 | 55 | 9 | 0.56 ± 0.04 | 79.3 ± 0.4 | 3.0 ± 0.2 | 0.33 ± 0.06 | 42.3 ± 2.5 | 3.5 ± 0.8 |
| 14 | 0 | 0 | 0 | 1 | 105 | 8 | 0.43 ± 0.04 | 75.4 ± 2.1 | 1.8 ± 0.5 | 0.42 ± 0.02 | 59.7 ± 4.2 | 4.4 ± 0.4 |
| 15 | 0 | 0 | 0 | 1 | 105 | 8 | 0.51 ± 0.05 | 69.8 ± 4.3 | 2.2 ± 0.6 | 0.39 ± 0.01 | 62.2 ± 1.9 | 2.9 ± 0.3 |
| 16 | 0 | 0 | 0 | 1 | 105 | 8 | 0.45 ± 0.01 | 73.7 ± 0.7 | 1.5 ± 0.1 | 0.43 ± 0.09 | 61.9 ± 2.5 | 3.9 ± 0.5 |
| 17 | 0 | 0 | 0 | 1 | 105 | 8 | 0.47 ± 0.03 | 65.7 ± 1.5 | 1.3 ± 0.1 | 0.41 ± 0.08 | 58.9 ± 2.3 | 3.6 ± 0.2 |
| Factors | Coeff. | Julius | Ponticus | ||
|---|---|---|---|---|---|
| Proteins conc. (mg/mL) | IAA (%) | Proteins conc. (mg/mL) | IAA (%) | ||
| Constants | |||||
| b0 | 0.472 | 71.5 | 0.406 | 60.6 | |
| Linear | |||||
| X10 | b1 | −0.071 ** | −1.0 | −0.212 *** | −13.8 *** |
| X6 | b2 | 0.086 ** | −1.7 | −0.115 * | 5.1 ** |
| X8 | b3 | 0.102 *** | −1.4 | −0.149 ** | −14.6 *** |
| Quadratic | |||||
| X10* X10 | b11 | 0.023 | 11.4** | 0.089 | −0.2 |
| X6* X6 | b22 | −0.047 | 11.5** | 0.156 * | −14.4 |
| X8* X8 | b33 | 0.098 ** | 7.1* | 0.112 | −0.8 |
| Interaction | |||||
| X10* X6 | b12 | −0.017 | −1.6 | −0.208 * | −2.0 |
| X10* X8 | b13 | 0.031 | −6.7 | 0.049 | −8.8 * |
| X6* X8 | b23 | −0.039 | 3.6 | 0.356 ** | −1.1 |
| R2 | 0.94 | 0.91 | 0.93 | 0.98 | |
| AF | 1.05 | 1.02 | 1.08 | 1.03 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tchewonpi Sagu, S.; Huschek, G.; Bönick, J.; Homann, T.; Rawel, H.M. A New Approach of Extraction of α-Amylase/trypsin Inhibitors from Wheat (Triticum aestivum L.), Based on Optimization Using Plackett–Burman and Box–Behnken Designs. Molecules 2019, 24, 3589. https://doi.org/10.3390/molecules24193589
Tchewonpi Sagu S, Huschek G, Bönick J, Homann T, Rawel HM. A New Approach of Extraction of α-Amylase/trypsin Inhibitors from Wheat (Triticum aestivum L.), Based on Optimization Using Plackett–Burman and Box–Behnken Designs. Molecules. 2019; 24(19):3589. https://doi.org/10.3390/molecules24193589
Chicago/Turabian StyleTchewonpi Sagu, Sorel, Gerd Huschek, Josephine Bönick, Thomas Homann, and Harshadrai M. Rawel. 2019. "A New Approach of Extraction of α-Amylase/trypsin Inhibitors from Wheat (Triticum aestivum L.), Based on Optimization Using Plackett–Burman and Box–Behnken Designs" Molecules 24, no. 19: 3589. https://doi.org/10.3390/molecules24193589
APA StyleTchewonpi Sagu, S., Huschek, G., Bönick, J., Homann, T., & Rawel, H. M. (2019). A New Approach of Extraction of α-Amylase/trypsin Inhibitors from Wheat (Triticum aestivum L.), Based on Optimization Using Plackett–Burman and Box–Behnken Designs. Molecules, 24(19), 3589. https://doi.org/10.3390/molecules24193589