Effects of Atmospheric and Room Temperature Plasma (ARTP) Mutagenesis on Physicochemical Characteristics and Immune Activity In Vitro of Hericium erinaceus Polysaccharides


 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Comparative Analysis of Agronomic Characters and Polysaccharide Content
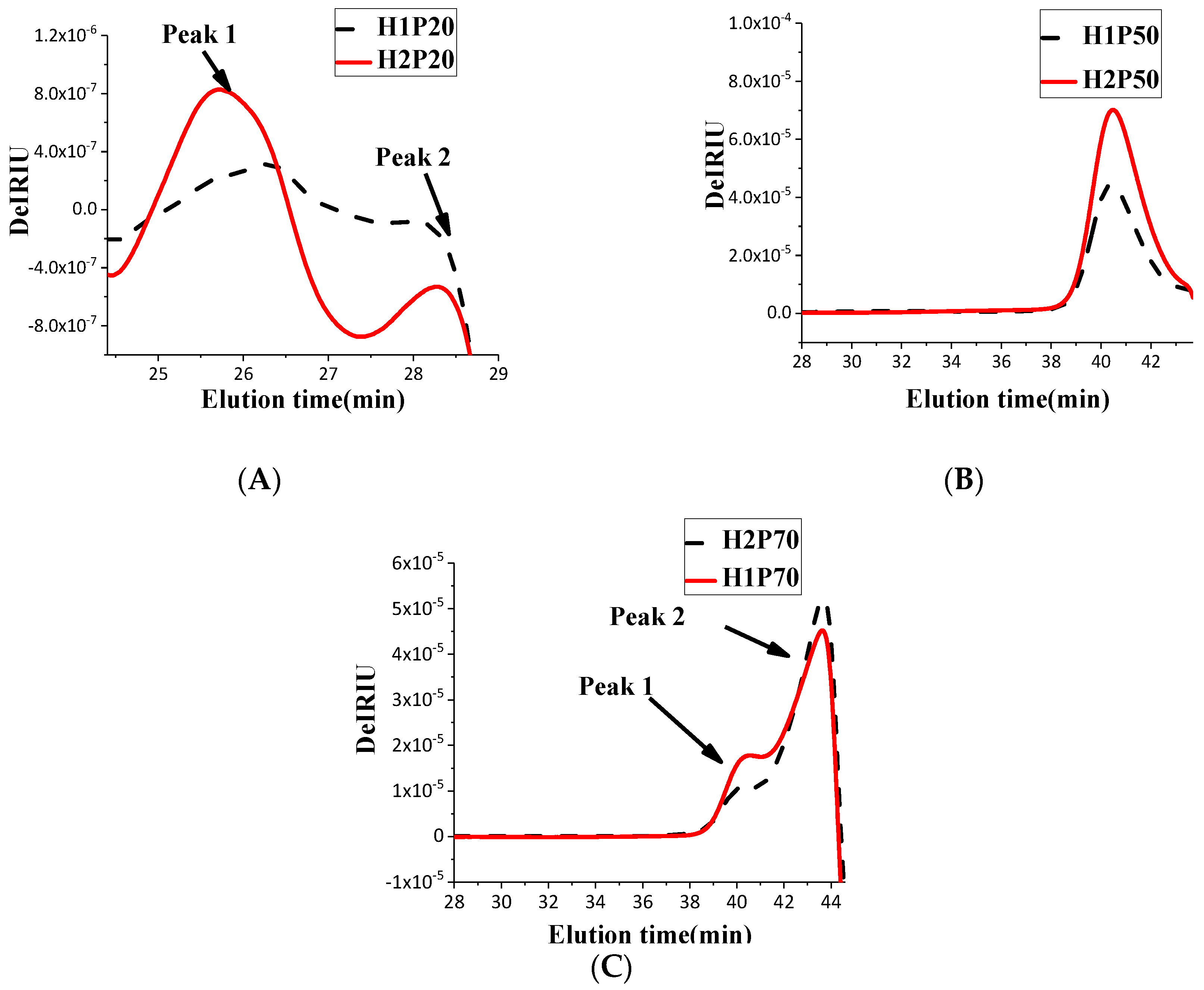
2.2. Effect of ARTP on Polysaccharide Molecular Weight Distribution Pattern
2.3. Effect of ARTP on Monosaccharide Composition of Hydrolyzed Polysaccharide Fractions
2.4. Fourier Transform Infrared Spectrum Analysis
2.5. Effect of ARTP on Nitric Oxide Production from Macrophages
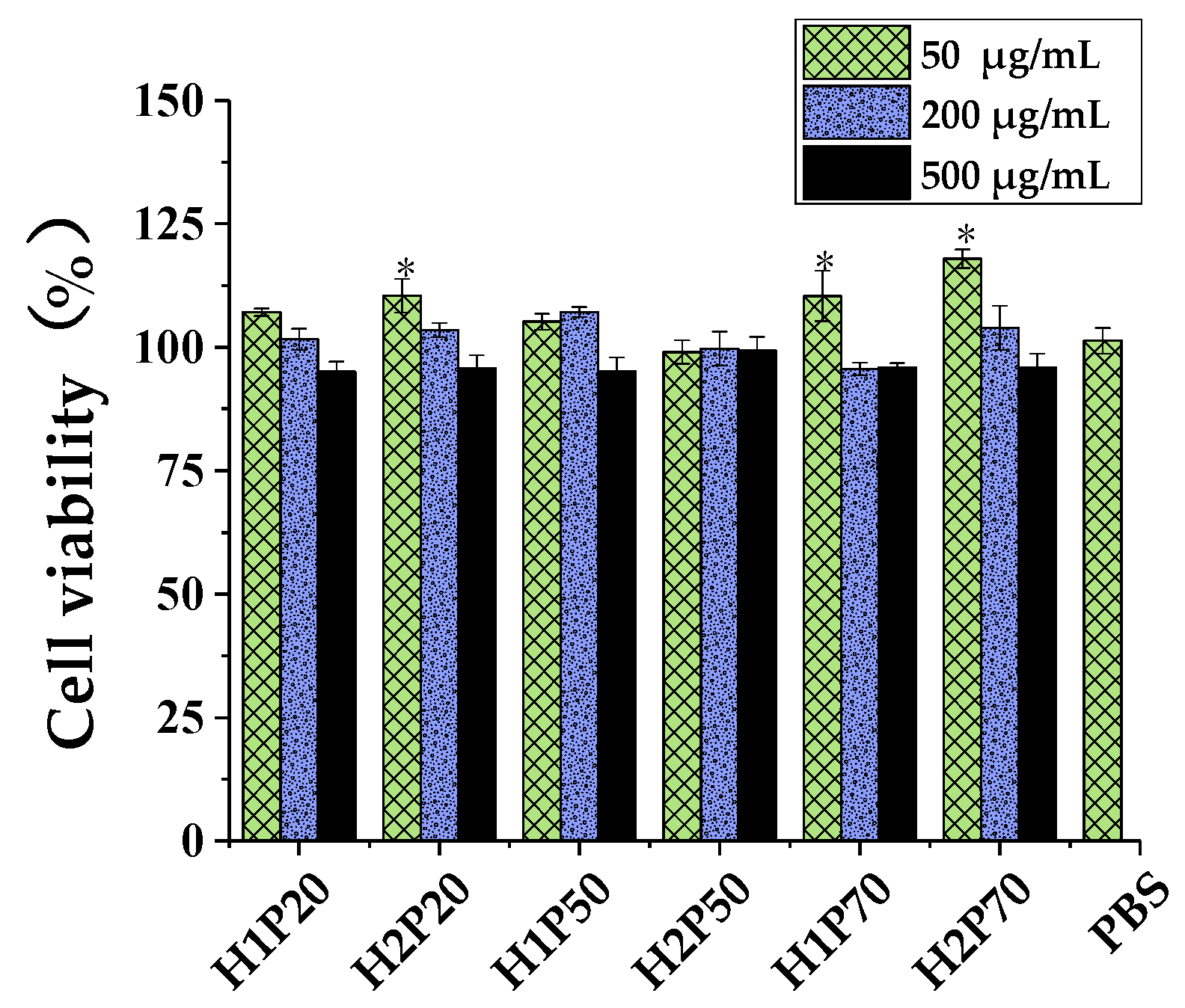
2.5.1. Cell Viability Assay
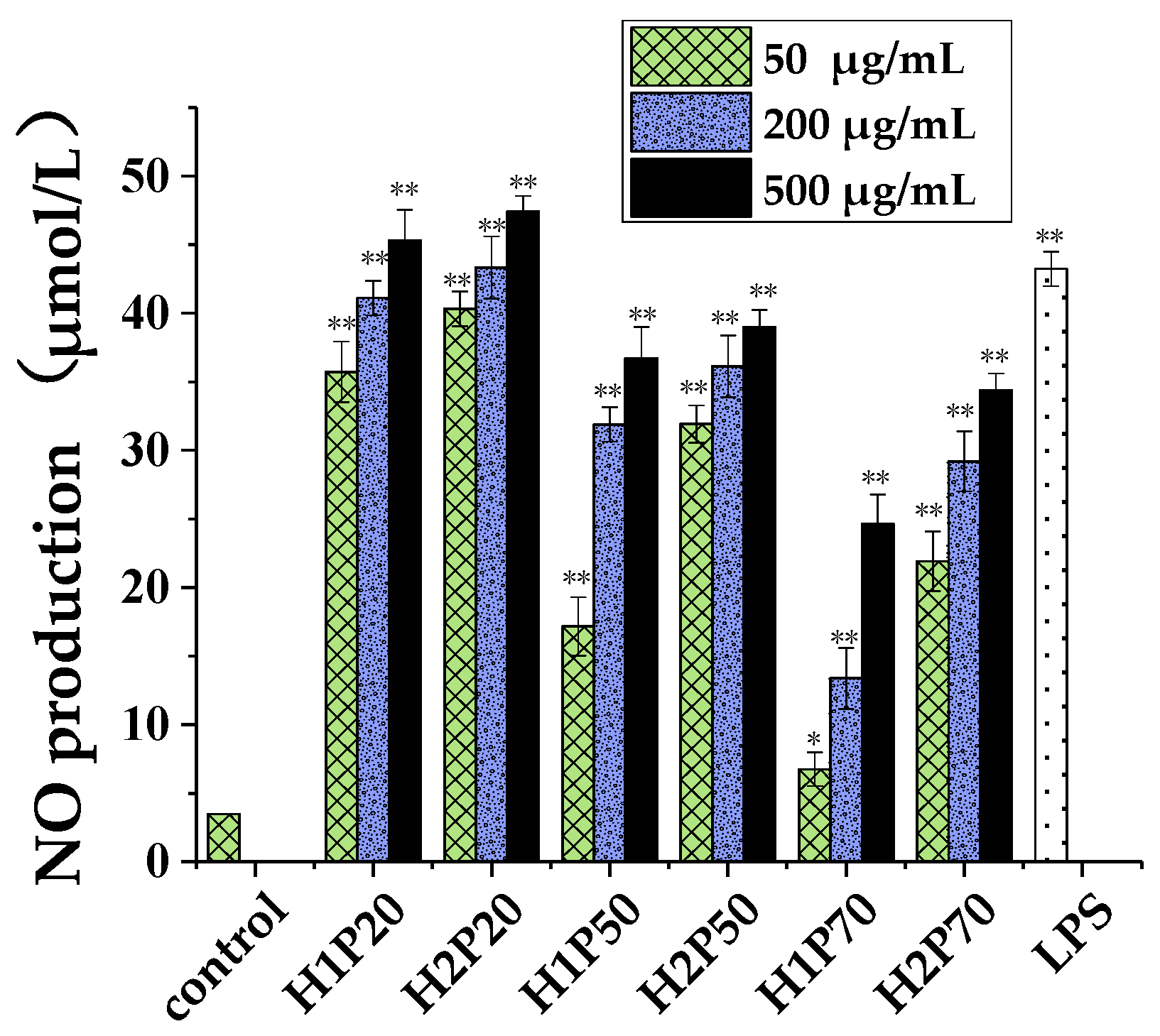
2.5.2. Nitric Oxide Released by RAW264.7 Macrophages
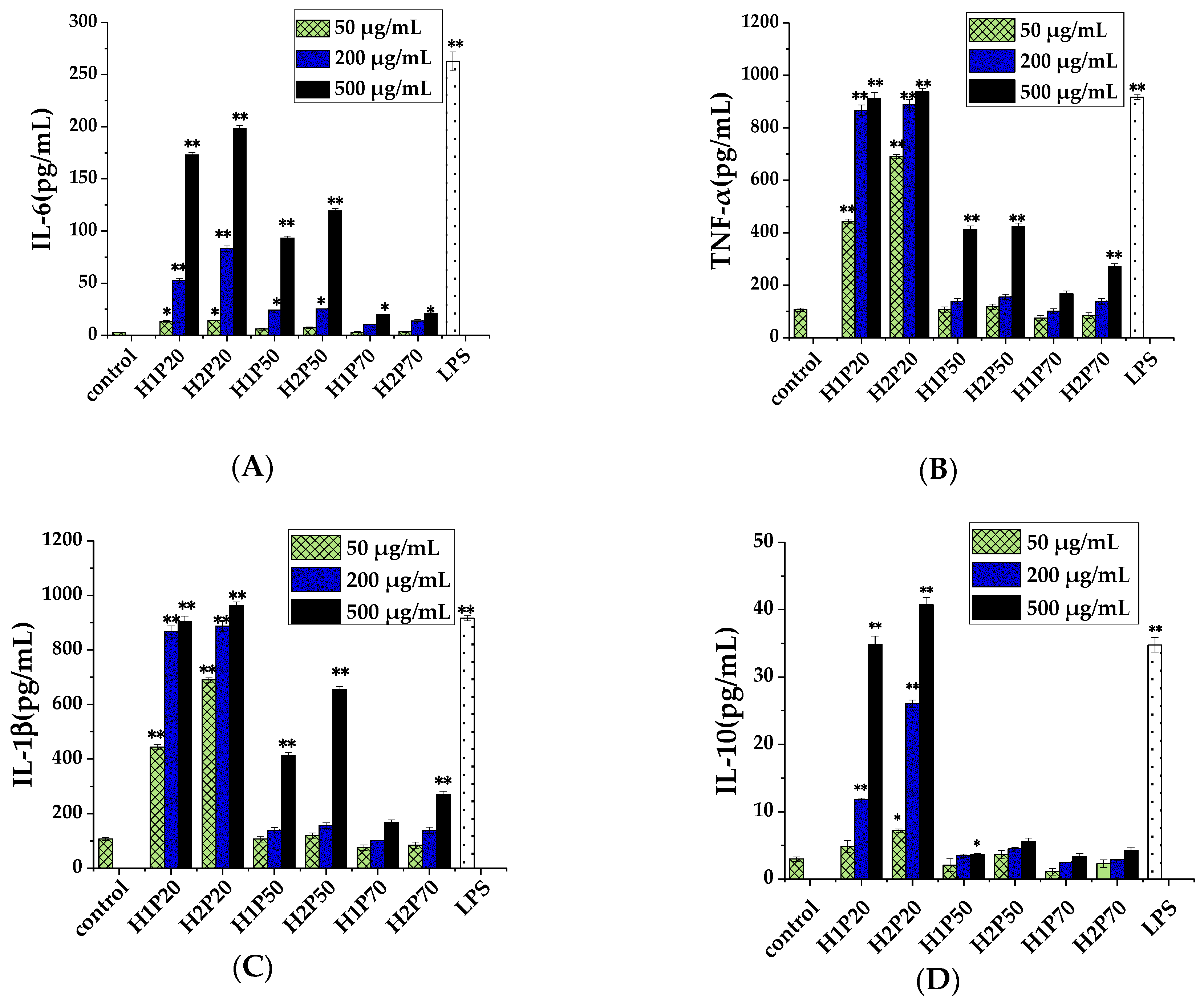
2.6. Effect of ARTP on Immunostimulatory Activity
3. Materials and Methods
3.1. Materials and Chemicals
3.2. Fruit Body Cultivation
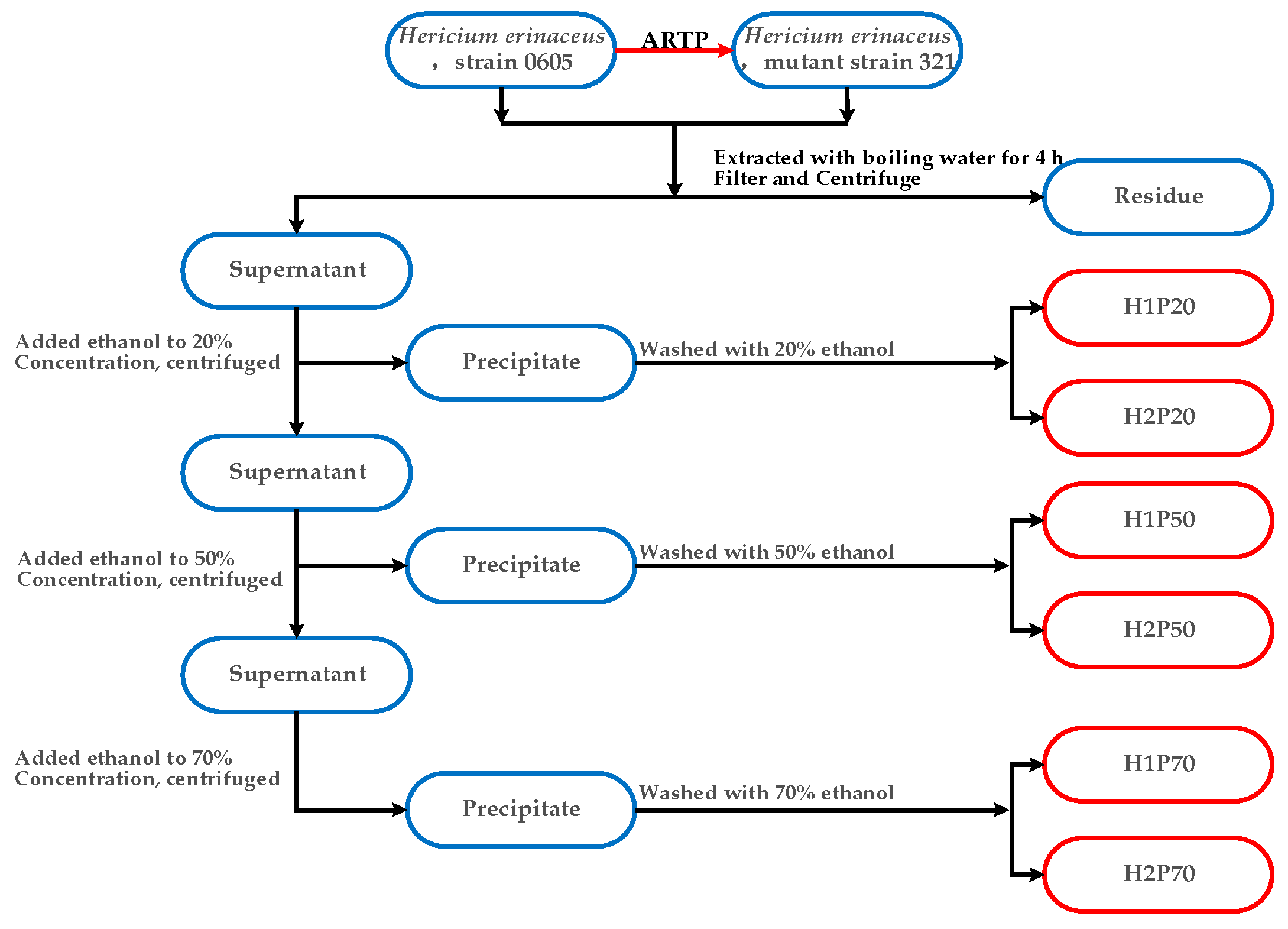
3.3. Extraction and Isolation of Polysaccharide Components
3.4. Total Polysaccharide Content
3.5. Fourier Transform Infrared Spectrum Analysis of Polysaccharide Components
3.6. Molecular Weight Distribution among the Polysaccharide Components
3.7. Monosaccharide Composition Analysis
3.8. RAW264.7 Macrophages Trial
3.8.1. Cell Viability Assay
3.8.2. Nitric Oxide Released by RAW264.7 Macrophages
3.9. THP-1 Macrophage Differentiation Trial
3.9.1. Cell Culture
3.9.2. Macrophage Differentiation and Quantitative Cytokines Trial
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhu, F.; Du, B.; Xu, B. Preparation and characterization of polysaccharides from mushrooms. In Polysaccharides: Bioactivity and Biotechnology; Ramawat, K.G., Mérillon, J.-M., Eds.; Springer International Publishing: New York, NY, USA, 2015; pp. 1009–1027. [Google Scholar] [CrossRef]
- Yang, B.-K.; Park, J.-B.; Song, C.-H. Hypolipidemic effect of an exo-biopolymer produced from a submerged mycelial culture of Hericium erinaceus. Biosci. Biotechnol. Biochem. 2003, 67, 1292–1298. [Google Scholar] [CrossRef]
- Li, G.; Yu, K.; Li, F.; Xu, K.; Li, J.; He, S.; Cao, S.; Tan, G. Anticancer potential of Hericium erinaceus extracts against human gastrointestinal cancers. J. Ethnopharmacol. 2014, 153, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.-C.; Yin, X.; Cao, C.Y.; Wei, J.; Zhang, Q.; Gao, J.-M. Chemical constituents from Hericium erinaceus and their ability to stimulate NGF-mediated neurite outgrowth on PC12 cells. Bioorg. Med. Chem. Lett. 2015, 25, 5078–5082. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Hong, E.K. Hericium erinaceus enhances doxorubicin-induced apoptosis in human hepatocellular carcinoma cells. Cancer Lett. 2010, 297, 144–154. [Google Scholar] [CrossRef]
- Friedman, M. Chemistry, nutrition, and health-promoting properties of Hericium erinaceus (Lion’s Mane) mushroom fruiting bodies and mycelia and their bioactive compounds. J. Agric. Food Chem. 2015, 63, 7108–7123. [Google Scholar] [CrossRef] [PubMed]
- Charumathy, M.; Sudha, G.; Balakrishnan, P. Detection of antioxidant activity and bioactive constituents in the fruiting bodies of Hericium erinaceus pers-an edible mushroom. J. Pharm. Pham. Sci. 2016, 8, 152–156. [Google Scholar]
- Li, Q.-Z.; Wu, D.; Chen, X.; Zhou, S.; Liu, Y.; Yang, Y.; Cui, F. Chemical compositions and macrophage activation of polysaccharides from Leon’s mane culinary-medicinal mushroom Hericium erinaceus (Higher Basidiomycetes) in different maturation stages. Int. J. Med. Mushrooms 2015, 17, 443–452. [Google Scholar] [CrossRef]
- Sun, L.; Liu, Q.; Bao, C.; Fan, J. Comparison of free total amino acid compositions and their functional classifications in 13 wild edible mushrooms. Molecules 2017, 22, 350. [Google Scholar] [CrossRef]
- Dong, X.-y.; Xiu, Z.L.; Min Hou, Y.; Li, S.; Jia Zhang, D.; Sheng Ren, C. Enhanced production of 1,3-propanediol in Klebsiella pneumoniae induced by dielectric barrier discharge plasma in atmospheric air. IEEE Trans. Plasma. Sci. 2009, 37, 920–926. [Google Scholar] [CrossRef]
- Chen, H.; Chen, Z.J.; Wu, M.B.; Deng, S.X. Screening the fusarium graminearum inhibitory mutant strain from Bacillus subtilis by atmospheric-pressure plasma jet. J. Appl. Microbiol. 2010, 108, 96–103. [Google Scholar] [CrossRef]
- Fang, M.; Jin, L.; Zhang, C.; Tan, Y.; Jiang, P.X.; Ge, N.; Li, H.P.; Xing, X. Rapid mutation of Spirulina platensis by a new mutagenesis system of Atmospheric and Room Temperature Plasmas (ARTP) and generation of a mutant library with diverse phenotypes. PLoS ONE 2013, 8, e77046. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.Y.; Huang, Z.; Li, G.; Zhao, H.X.; Xing, X.H.; Sun, W.; Li, H.P.; Gou, Z.; Bao, C.Y. Novel mutation breeding method for Streptomyces avermitilis using an atmospheric pressure glow discharge plasma. J. Appl. Microbiol. Biot. 2010, 108, 851–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Bai, F.; Xiu, Z.L. Oxidative stress induced in saccharomyces cerevisiae exposed to dielectric barrier discharge plasma in air at atmospheric pressure. IEEE Trans. Plasma Sci. 2010, 38, 1885–1891. [Google Scholar] [CrossRef]
- Heine, W.; Uhlemann, M.; Mohr, C. Physiologische besiedlung des darmtrakts in der kindheit, ihre pathologischen abweichungen und beeinflussung durch die ahrung. Monatsschr. Kinderh. 1998, 146. [Google Scholar] [CrossRef]
- Ottenheim, C.; Nawrath, M.; Chuan Wu, J. Microbial mutagenesis by atmospheric and room-temperature plasma (ARTP): The latest development. Bioresour. Bioprocess. 2018, 5, 12. [Google Scholar] [CrossRef]
- Sheng, X.; Yan, J.; Meng, Y.; Kang, Y.; Han, Z.; Tai, G.; Zhou, Y.; Cheng, H. Immunomodulatory effects of Hericium erinaceus derived polysaccharides are mediated by intestinal immunology. Food Funct. 2017, 8, 1020–1027. [Google Scholar] [CrossRef] [PubMed]
- Ren, F.; Chen, L.; Tong, Q. Highly improved acarbose production of Actinomyces through the combination of ARTP and penicillin susceptible mutant screening. World J. Microbiol. Biotechnol. 2017, 33, 16. [Google Scholar] [CrossRef] [PubMed]
- He, J.H.; Jiang, W.; Lv, B.B.; Li, P.; Wu, G.; Wang, J.B.; Zhu, Z.P.; Wu, X.; Tang, X.M. Screening and RAPD analysis of Volvariella volvacea ARTP mutants. Indian J. Agric. Sci. 2014, 28, 1950–1955. [Google Scholar]
- Qiang, W.; Ling-Ran, F.; Luo, W.; Han-Guang, L.; Lin, W.; Ya, Z.; Yu, X.B. Mutation breeding of lycopene-producing strain Blakeslea Trispora by a novel Atmospheric and Room Temperature Plasma (ARTP). Appl. Biochem. Biotech. 2014, 174, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Trowsdale, J.; Parham, P.; Trowsdale, J.; Parham, P. Mini-review: Defense strategies and immunity-related genes. Eur. J. Immunol. 2004, 34, 7–17. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, X.F.; Li, H.-P.; Wang, L.Y.; Zhang, C.; Xing, X.H.; Bao, C.Y. Atmospheric and room temperature plasma (ARTP) as a new powerful mutagenesis tool. Appl. Microbiol. Biot. 2014, 98, 5387–5396. [Google Scholar] [CrossRef]
- Wang, H.X.; Ng, T.B. A new laccase from dried fruiting bodies of the monkey head mushroom Hericium erinaceum. Biochem. Bioph. Res. Commun. 2004, 322, 17–21. [Google Scholar] [CrossRef]
- Porcheray, F.; Viaud, S.; Rimaniol, A.C.; Léone, C.; Samah, B.; Dereuddre, N.; Dormont, D.; Gras, G. Macrophage activation switching: An asset for the resolution of inflammation. Clin. Exp. Immunol. 2006, 142, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.Z.; Hao, R.; Zha, X.Q.; Pan, L.H.; Liu, J.; Luo, J.P. Polysaccharide of dendrobium huoshanense activates macrophages via toll-like receptor 4-mediated signaling pathways. Carbohyd. Polym. 2016, 146, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xie, J.H.; Jia, S.; Huang, L.; Wang, Z.; Li, C.; Xie, M. Immunomodulatory effects of an acetylated Cyclocarya paliurus polysaccharide on murine macrophages RAW264.7. Int. J. Biol. Macromol. 2017, 98, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Surayot, U.; You, S. Structural effects of sulfated polysaccharides from Codium fragile on NK cell activation and cytotoxicity. Int. J. Biol. Macromol. 2017, 98, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhou, C.; Zhou, D.; Ou, S.; Huang, H. Structural characterization of a novel polysaccharide fraction from Hericium erinaceus and its signaling pathways involved in macrophage immunomodulatory activity. J. Funct. Foods 2017, 37, 574–585. [Google Scholar] [CrossRef]
- Liao, W.; Luo, Z.; Liu, D.; Ning, Z.; Yang, J.; Ren, J. Structure characterization of a novel polysaccharide from Dictyophora indusiata and its macrophage immunomodulatory activities. J. Agric. Food Chem. 2015, 63, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Li, S.; Lin, Y.; Liu, R.; Gu, Y.; Liao, D. Effectiveness of cultured Cordyceps sinensis combined with glucocorticosteroid on pulmonary fibrosis induced by bleomycin in rats. China J. Chin. Mater. Med. 2011, 36, 2265–2270. [Google Scholar]
- Cheong, K.L.; Meng, L.Z.; Chen, X.Q.; Wang, L.Y.; Wu, D.T.; Zhao, J.; Li, S.P. Structural elucidation, chain conformation and immuno-modulatory activity of glucogalactomannan from cultured Cordyceps sinensis fungus UM01. J. Funct. Foods 2016, 25, 174–185. [Google Scholar] [CrossRef]
- Yu, Q.; Nie, S.P.; Wang, J.; Huang, D.; Li, W.; Xie, M.Y. Signaling pathway involved in the immunomodulatory effect of Ganoderma atrum polysaccharide in Spleen Lymphocytes. J. Agric. Food Chem. 2015, 63, 2734–3740. [Google Scholar] [CrossRef] [PubMed]
- Ghonime, M.; Emara, M.; Shawky, R.; Soliman, H.; El-Domany, R.; Abdelaziz, A. Immunomodulation of RAW 264.7 murine macrophage functions and antioxidant activities of 11 plant extracts. Immunol. Investig. 2015, 44, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Schepetkin, I.; Quinn, M.; Schepetkin, I.A.; Quinn, M.T. Botanical polysaccharides: Macrophage immunomodulation and therapeutic potential. Int. Immunopharmacol. 2006, 6, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.-Z.; Wu, D.; Zhou, S.; Liu, Y.F.; Li, Z.p.; Feng, J.; Yan, Y. Structure elucidation of a bioactive polysaccharide from fruiting bodies of Hericium erinaceus in different maturation stages. Carbohydr. Polym. 2016, 144, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.Y. Comparison of yield of different Ganoderma lucidum fruiting bodies and content of polysaccharides in fruiting bodies. J. Changsha Univ. 2009, 23, 18–19. [Google Scholar]
- Ferreira, S.S.; Passos, C.P.; Madureira, P.; Vilanova, M.; Coimbra, M.A. Structure–function relationships of immunostimulatory polysaccharides: A review. Carbohydr. Polym. 2015, 132, 378–396. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Sun, Z.; Ma, X.; Yang, B.; Jiang, Y.; Wei, D.; Chen, F. Mutation breeding of extracellular polysaccharide-producing microalga Crypthecodinium cohnii by a novel mutagenesis with atmospheric and room temperature plasma. Int. J. Mol. Sci. 2015, 16, 8201–8212. [Google Scholar] [CrossRef]
- Xu, M.; McCanna, D.J.; Sivak, J.G. Use of the viability reagent PrestoBlue in comparison with alamarBlue and MTT to assess the viability of human corneal epithelial cells. J. Pharmacol. Toxicol. Met. 2015, 71, 1–7. [Google Scholar] [CrossRef]
- Al-Nasiry, S.; Geusens, N.; Hanssens, M.; Luyten, C.; Pijnenborg, R. The use of Alamar Blue assay for quantitative analysis of viability, migration and invasion of choriocarcinoma cells. Hum. Reprod. 2007, 22, 1304–1309. [Google Scholar] [CrossRef] [Green Version]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Šafránková, B.; Hermannová, M.; Nesporova, K.; Velebný, V.; Kubala, L. Absence of differences among low, middle, and high molecular weight hyaluronan in activating murine immune cells in vitro. Int. J. Biol. Macromol. 2018, 107, 1–8. [Google Scholar] [CrossRef] [PubMed]
- A Cleary, J.; E Kelly, G.; Husband, A.J. The effect of molecular weight and β-1,6-linkages on priming of macrophage function in mice by (1,3)-β-d-glucan. Immunol. Cell Biol. 1999, 77, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Kao, P.F.; Wang, S.H.; Hung, W.T.; Liao, Y.-H.; Lin, C.M.; Yang, W.B. Structural characterization and antioxidative activity of low-molecular-weights β-1,3-glucan from the residue of extracted Ganoderma lucidum fruiting bodies. J. Biomed. Biotechnol. 2012, 673764. [Google Scholar] [CrossRef]
- Krizková, L.; Durackova, Z.; Sandula, J.; Slamenová, D.; Sasinkova, V.; Sivonová, M.; Krajcovic, J. Fungal beta-(1-3)-D-glucan derivatives exhibit high antioxidative and antimutagenic activity in vitro. Anticancer Res. 2003, 23, 2751–2756. [Google Scholar] [CrossRef]
- Al-Banna, N.A.; Cyprian, F.; Albert, M.J. Cytokine responses in campylobacteriosis: Linking pathogenesis to immunity. Cytokine Growth FR. 2018, 41, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Fukuhara, Y.D.M.; Reis, M.L.; Dellalibera-Joviliano, R.; Cunha, F.Q.C.; Donadi, E.A. Increased plasma levels of IL-1β, IL-6, IL-8, IL-10 and TNF-α in patients moderately or severely envenomed by Tityus serrulatus scorpion sting. Toxicon 2003, 41, 49–55. [Google Scholar] [CrossRef]
- Wu, Y.; Cui, S.W.; Tang, J.; Wang, Q.; Gu, X. Preparation, partial characterization and bioactivity of water-soluble polysaccharides from boat-fruited sterculia seeds. Carbohydr. Polym. 2007, 70, 437–443. [Google Scholar] [CrossRef]
- Wang, Y.; Shao, J.; Yao, S.; Zhang, S.; Yan, J.; Wang, H.; Chen, Y. Study on the antithrombotic activity of Umbilicaria esculenta polysaccharide. Carbohydr. Polym. 2014, 105, 231–236. [Google Scholar] [CrossRef]
- Chen, Y.C.; Shen, S.-C.; Chen, L.G.; Lee, T.; Yang, L. Wogonin, baicalin, and baicalein inhibition of inducible nitric oxide synthase and cyclooxygenase-2 gene expressions induced by nitric oxide synthase inhibitors and lipopolysaccharide. Biochem. Pharmacol. 2001, 61, 1417–1427. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Shape | Fleshy Quality | Spiny Length/cm | Yield/g | Polysaccharide content (%) |
|---|---|---|---|---|---|
| 0605 | Round | Tight relatively | 1.31 ± 1.09 | 113 ± 11.16 a | 5.16 ± 0.25 c |
| 321 | Large and round | Tight | 1.83 ± 0.22 | 138 ± 5.37 b | 6.03 ± 0.09 d |
| Fraction | Peak1 | Peak2 | ||||||
|---|---|---|---|---|---|---|---|---|
| Mw (Da) | Mn (Da) | Mw/Mn | Ratio (%) | Mw (Da) | Mn (Da) | Mw/Mn | Ratio (%) | |
| H1P20 | 6.30 × 106 | 5.84 × 106 | 1.16 | 51.7 | 2.33 × 105 | 1.87 × 105 | 1.87 | 48.3 |
| H2P20 | 2.18 × 107 | 2.04 × 107 | 1.07 | 78.8 | 2.80 × 105 | 2.49 × 105 | 1.50 | 21.2 |
| H1P50 | 2.27 × 104 | 2.22 × 104 | 1.11 | 100 | - | - | - | - |
| H2P50 | 6.52 × 104 | 4.27 × 104 | 1.47 | 100 | - | - | - | - |
| H1P70 | 1.65 × 104 | 1.33 × 104 | 1.23 | 15.5 | 3.97 × 104 | 3.08 × 104 | 1.28 | 84.5 |
| H2P70 | 9.78 × 104 | 6.96 × 104 | 1.40 | 28.1 | 3.77 × 104 | 2.50 × 104 | 1.50 | 71.9 |
| Sample | Fuc | Ara | GlcN | Gal | Glu | Xyl | Man | Fru |
|---|---|---|---|---|---|---|---|---|
| H1P20 | 1.89 | 1.02 | 0.86 | 0.73 | 3.91 | 0.18 | 1.00 | - |
| H2P20 | 2.00 | 0.63 | 0.41 | 1.30 | 4.98 | 0.75 | 1.00 | 0.56 |
| H1P50 | 1.29 | - | 0.19 | 1.74 | 2.45 | - | 1.00 | - |
| H2P50 | 7.12 | 0.16 | 0.29 | 3.72 | 4.35 | 0.21 | 1.00 | - |
| H1P70 | 0.81 | - | - | 2.46 | 2.82 | - | 1.00 | - |
| H2P70 | 3.71 | - | 0.21 | 2.33 | 9.99 | - | 1.00 | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, L.; Wu, D.; Zhang, H.; Li, Q.; Zhang, Z.; Liu, Y.; Zhou, S.; Wang, W.; Li, Z.; Yang, Y. Effects of Atmospheric and Room Temperature Plasma (ARTP) Mutagenesis on Physicochemical Characteristics and Immune Activity In Vitro of Hericium erinaceus Polysaccharides. Molecules 2019, 24, 262. https://doi.org/10.3390/molecules24020262
Zhu L, Wu D, Zhang H, Li Q, Zhang Z, Liu Y, Zhou S, Wang W, Li Z, Yang Y. Effects of Atmospheric and Room Temperature Plasma (ARTP) Mutagenesis on Physicochemical Characteristics and Immune Activity In Vitro of Hericium erinaceus Polysaccharides. Molecules. 2019; 24(2):262. https://doi.org/10.3390/molecules24020262
Chicago/Turabian StyleZhu, Lingli, Di Wu, Henan Zhang, Qiaozhen Li, Zhong Zhang, Yanfang Liu, Shuai Zhou, Wenhan Wang, Zhengpeng Li, and Yan Yang. 2019. "Effects of Atmospheric and Room Temperature Plasma (ARTP) Mutagenesis on Physicochemical Characteristics and Immune Activity In Vitro of Hericium erinaceus Polysaccharides" Molecules 24, no. 2: 262. https://doi.org/10.3390/molecules24020262
APA StyleZhu, L., Wu, D., Zhang, H., Li, Q., Zhang, Z., Liu, Y., Zhou, S., Wang, W., Li, Z., & Yang, Y. (2019). Effects of Atmospheric and Room Temperature Plasma (ARTP) Mutagenesis on Physicochemical Characteristics and Immune Activity In Vitro of Hericium erinaceus Polysaccharides. Molecules, 24(2), 262. https://doi.org/10.3390/molecules24020262