Water Extracts of Hull-less Waxy Barley (Hordeum vulgare L.) Cultivar ‘Boseokchal’ Inhibit RANKL-induced Osteoclastogenesis

 ,
,  and
and
Abstract
:1. Introduction
2. Results
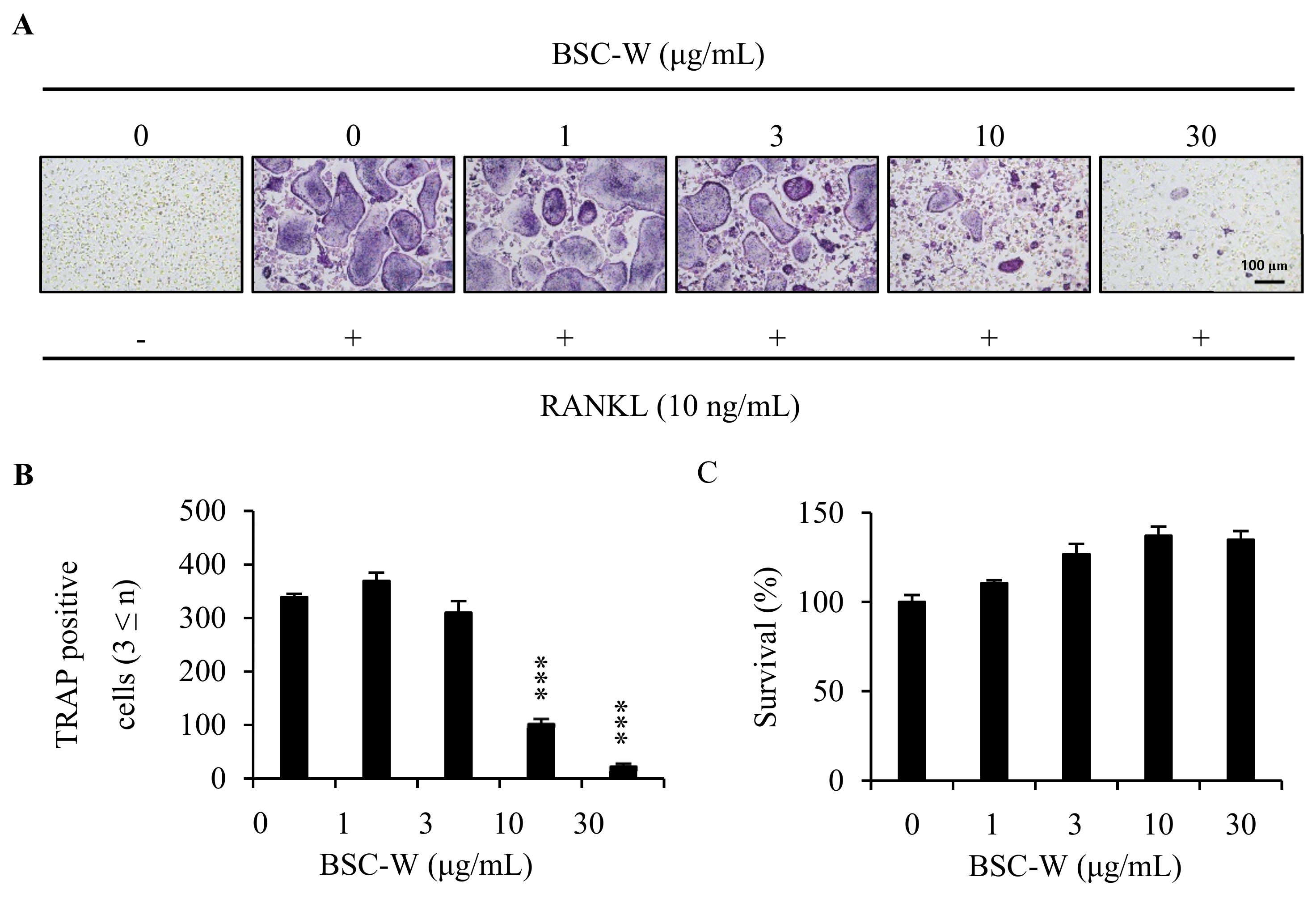
2.1. BSC-W Inhibited RANKL-Mediated Osteoclast Differentiation
2.2. BSC-W Had No Cytotoxic Effect On Bmms
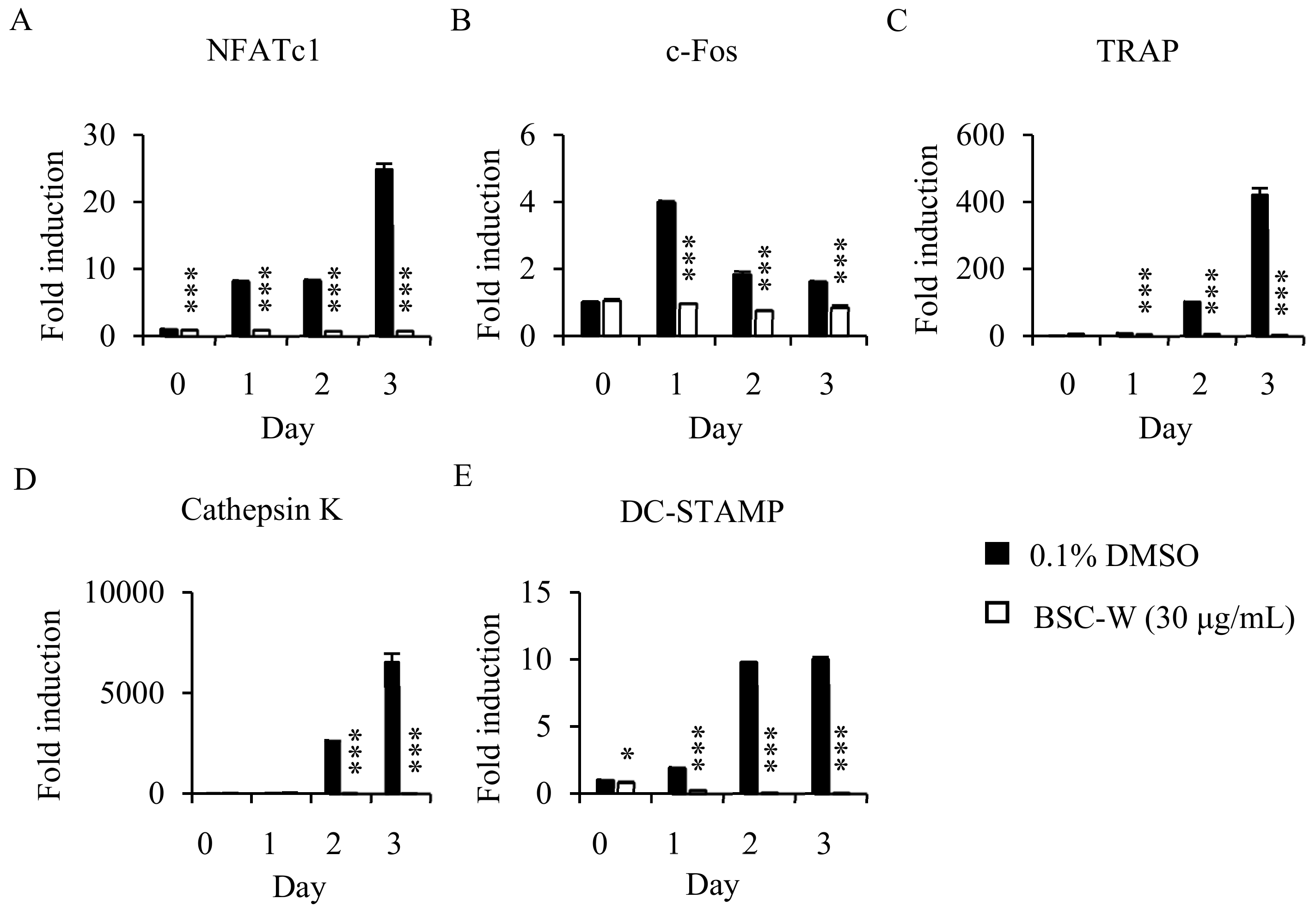
2.3. Effects of BSC-W on RANKL-Induced Gene Expression
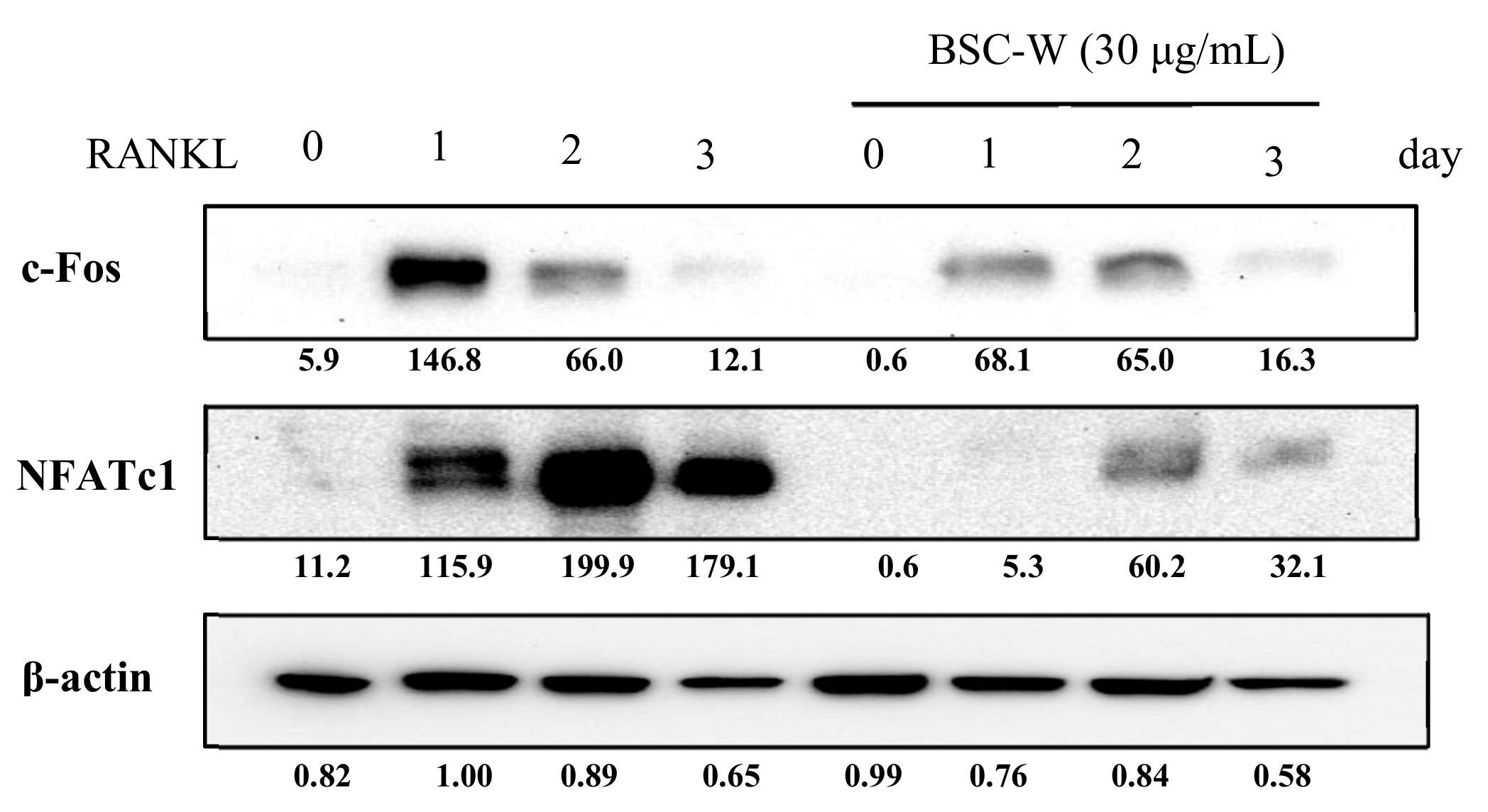
2.4. BSC-W Inhibited RANKL-Induced Protein Expression of NFATc1
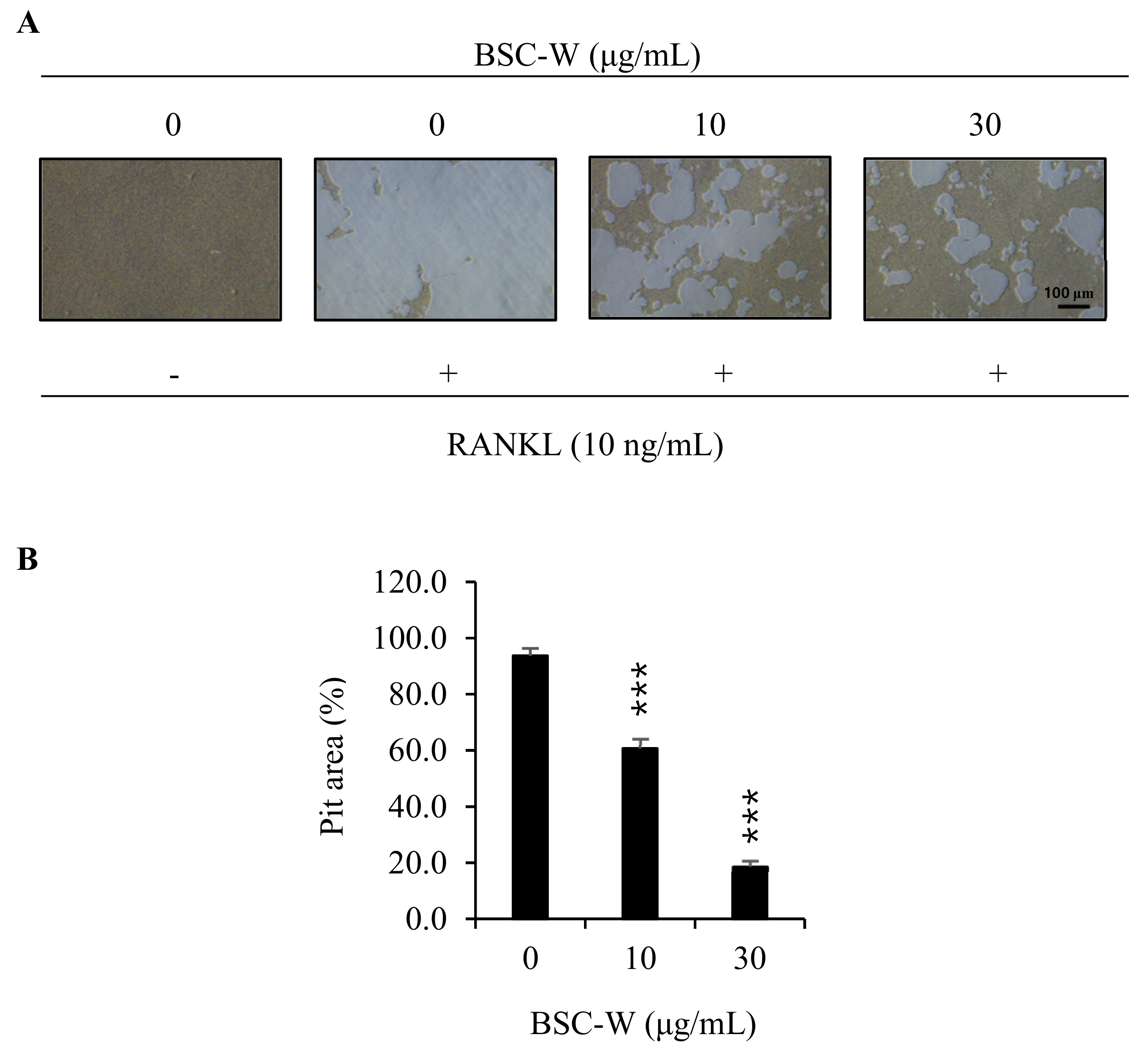
2.5. Effects of BSC-W on RANKL-Mediated Bone Resorptive Activity of Osteoclasts
3. Discussion
4. Materials and Methods
4.1. Preparation of the Boseokchal Water Extract
4.2. Cell Culture and Osteoclast Differentiation
4.3. Tartrate-Resistant Acid Phosphatase (TRAP) Staining
4.4. Cytotoxicity Assay for Extracts of BSC-W
4.5. Real-Time PCR
4.6. Western Blot
4.7. Bone Pit Formation Assay
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Goltzman, D. Discoveries, drugs and skeletal disorders. Nat. Rev. Drug Discov. 2002, 1, 784–796. [Google Scholar] [CrossRef] [PubMed]
- Rodan, G.A. Therapeutic Approaches to Bone Diseases. Science 2000, 289, 1508–1514. [Google Scholar] [CrossRef] [PubMed]
- NIH Consensus Development. Osteoporosis prevention, diagnosis, and therapy. JAMA 2000, 285, 785–795. [Google Scholar]
- Vaananen, H.K.; Laitala-Leinonen, T. Osteoclast lineage and function. Arch. Biochem. Biophys. 2008, 473, 132–138. [Google Scholar] [CrossRef]
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation and activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef]
- Feng, X. RANKing Intracellular Signaling in Osteoclasts. IUBMB Life 2005, 57, 389–395. [Google Scholar] [CrossRef]
- Kansara, M.; Teng, M.W.; Smyth, M.J.; Thomas, D.M. Translational biology of osteosarcoma. Nat. Rev. Cancer 2014, 14, 722–735. [Google Scholar] [CrossRef]
- Galibert, L. The Involvement of Multiple Tumor Necrosis Factor Receptor (TNFR)-associated Factors in the Signaling Mechanisms of Receptor Activator of NF-kappa B, a Member of the TNFR Superfamily. J. Boil. Chem. 1998, 273, 34120–34127. [Google Scholar] [CrossRef]
- Wong, B.R.; Josien, R.; Lee, S.Y.; Vologodskaia, M.; Steinman, R.M.; Choi, Y. The TRAF family of signal transducers mediates NF-kappaB activation by the TRANCE receptor. J. Boil. Chem. 1998, 273, 28355–28359. [Google Scholar] [CrossRef]
- Teitelbaum, S.L. RANKing c-Jun in osteoclast development. J. Clin. Investig. 2004, 114, 463–465. [Google Scholar] [CrossRef] [Green Version]
- Takayanagi, H. The Role of NFAT in Osteoclast Formation. Ann. New York Acad. Sci. 2007, 1116, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, H. Osteoimmunology: Shared mechanisms and crosstalk between the immune and bone systems. Nat. Rev. Immunol. 2007, 7, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Wagner, E.F.; Eferl, R. Fos/AP-1 proteins in bone and the immune system. Immunol. Rev. 2005, 208, 126–140. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Kogawa, M.; Wada, S.; Takayanagi, H.; Tsujimoto, M.; Katayama, S.; Hisatake, K.; Nogi, Y. Essential Role of p38 Mitogen-activated Protein Kinase in Cathepsin K Gene Expression during Osteoclastogenesis through Association of NFATc1 and PU.1. J. Boil. Chem. 2004, 279, 45969–45979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yagi, M.; Miyamoto, T.; Sawatani, Y.; Iwamoto, K.; Hosogane, N.; Fujita, N.; Morita, K.; Ninomiya, K.; Suzuki, T.; Miyamoto, K.; et al. DC-STAMP is essential for cell–cell fusion in osteoclasts and foreign body giant cells. J. Exp. Med. 2005, 202, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Hyun, J.W.; Chung, H.S. Cyanidin and Malvidin from Oryza sativa cv. Heugjinjubyeo mediate cytotoxicity against human monocytic leukemia cells by arrest of G(2)/M phase and induction of apoptosis. J. Agric. Food Chem. 2004, 52, 2213–2217. [Google Scholar] [CrossRef]
- Ryu, S.; Han, S.; Park, S.; Kim, H. Antioxidant activity of blackish purple rice. Korean J. Crop. Sci. 2006, 51, 173–178. [Google Scholar]
- Ghosh, M.; Kim, I.S.; Lee, Y.M.; Hong, S.M.; Lee, T.H.; Lim, J.H.; Debnath, T.; Lim, B.O. The Effects of Aronia melanocarpa ‘Viking’ Extracts in Attenuating RANKL-Induced Osteoclastic Differentiation by Inhibiting ROS Generation and c-FOS/NFATc1 Signaling. Molecules 2018, 23, 615. [Google Scholar] [CrossRef]
- Dou, C.; Li, J.; Kang, F.; Cao, Z.; Yang, X.; Jiang, H.; Yang, B.; Xiang, J.; Xu, J.; Dong, S. Dual Effect of Cyanidin on RANKL-Induced Differentiation and Fusion of Osteoclasts. J. Cell Physiol. 2016, 231, 558–567. [Google Scholar] [CrossRef]
- Choi, J.; Suh, J.; Min, K.; Lee, M.; Park, J.; Kim, Y.; Park, T.; Hwang, J.; Kim, H.; Baek, S.; et al. A new colored naked barley cultivar” Zasoojeongchal” with waxy endosperm. Korean J. Breed. Sci. 2007, 39, 244–245. [Google Scholar]
- Lee, M.; Kim, Y.; Park, J.; Kim, M.; Hyun, J. Hull-less waxy barley (Hordeum vulgare L.) cultivar ‘Boseokchal’ with high anthocyanin content and purple lemma. Korean J. Breed. Sci. 2014, 46, 456–462. [Google Scholar] [CrossRef]
- Wang, Y.; Harding, S.V.; Thandapilly, S.J.; Tosh, S.M. Jones PJH, Ames NP. Barley beta-glucan reduces blood cholesterol levels via interrupting bile acid metabolism. Br. J. Nutr. 2017, 118, 822–829. [Google Scholar] [CrossRef] [PubMed]
- Crabtree, G.R.; Olson, E.N. NFAT signaling: Choreographing the social lives of cells. Cell 2002, 109, 67–79. [Google Scholar] [CrossRef]
- Takayanagi, H.; Kim, S.; Koga, T.; Nishina, H.; Isshiki, M.; Yoshida, H.; Saiura, A.; Isobe, M.; Yokochi, T.; Inoue, J.-I.; et al. Induction and Activation of the Transcription Factor NFATc1 (NFAT2) Integrate RANKL Signaling in Terminal Differentiation of Osteoclasts. Dev. Cell 2002, 3, 889–901. [Google Scholar] [CrossRef] [Green Version]
- Hogan, P.G.; Chen, L.; Nardone, J.; Rao, A. Transcriptional regulation by calcium, calcineurin, and NFAT. Genes Dev. 2003, 17, 2205–2232. [Google Scholar] [CrossRef] [Green Version]
- Asagiri, M.; Sato, K.; Usami, T.; Ochi, S.; Nishina, H.; Yoshida, H.; Morita, I.; Wagner, E.F.; Mak, T.W.; Serfling, E.; et al. Autoamplification of NFATc1 expression determines its essential role in bone homeostasis. J. Exp. Med. 2005, 202, 1261–1269. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-H.; Kim, K.-J.; Kang, H.J.; Son, Y.-J.; Choi, S.-W.; Lee, M.-J. The Dual Role of Oat Bran Water Extract in Bone Homeostasis Through the Regulation of Osteoclastogenesis and Osteoblast Differentiation. Molecules 2018, 23, 3119. [Google Scholar] [CrossRef]
- Kim, K.-J.; Yeon, J.-T.; Choi, S.-W.; Moon, S.-H.; Ryu, B.J.; Yu, R.; Park, S.-J.; Kim, S.H.; Son, Y.-J. Decursin inhibits osteoclastogenesis by downregulating NFATc1 and blocking fusion of pre-osteoclasts. Bone 2015, 81, 208–216. [Google Scholar] [CrossRef]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. Methods Mol. Boil. 2000, 132, 365–386. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Kim, H.; Kim, K.J.; Yeon, J.T.; Kim, S.H.; Won, D.H.; Choi, H.; Nam, S.J.; Son, Y.J.; Kang, H. Placotylene A, an inhibitor of the receptor activator of nuclear factor-kappaB ligand-induced osteoclast differentiation, from a Korean sponge Placospongia sp. Mar. Drugs 2014, 12, 2054–2065. [Google Scholar] [CrossRef] [PubMed]
- Johnell, O.; Kanis, J.A. An estimate of the worldwide prevalence and disability associated with osteoporotic fractures. Osteoporos. Int. 2006, 17, 1726–1733. [Google Scholar] [CrossRef] [PubMed]
- Harvey, N.; Dennison, E.; Cooper, C. Osteoporosis: Impact on health and economics. Nat. Rev. Rheumatol. 2010, 6, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Cauley, J.A. Public Health Impact of Osteoporosis. J. Gerontol. A Boil. Sci. Med. Sci. 2013, 68, 1243–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oka, Y.; Iwai, S.; Amano, H.; Irie, Y.; Yatomi, K.; Ryu, K.; Yamada, S.; Inaqaki, K.; Oquchi, K. Tea polyphenols inhibit rat osteoclast formation and differentiation. J. Pharmacol. Sci. 2012, 118, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Londzin, P.; Siudak, S.; Cegieła, U.; Pytlik, M.; Janas, A.; Waligóra, A.; Folwarczna, J. Phloridzin, an Apple Polyphenol, Exerted Unfavorable Effects on Bone and Muscle in an Experimental Model of Type 2 Diabetes in Rats. Nutrients 2018, 10, E1701. [Google Scholar] [CrossRef]
- Feng, Y.L.; Jiang, X.T.; Ma, F.F.; Han, J.; Tang, X.L. Resveratrol prevents osteoporosis by upregulating FoxO1 transcriptional activity. Int. J. Mol. Med. 2018, 41, 202–212. [Google Scholar] [CrossRef]
- Wang, X.; Batubara, I.; Yamauchi, K.; Mitsunaga, T. Identification and structure-activity relationship (SAR) of chemical constituents from Daemonorops draco (Willd.) Blume and selected commercial flavonoids on anti-osteoclastogenesis activity. Fitoterapia 2019, 138, 104280. [Google Scholar] [CrossRef]
- Yao, W.; Zhang, H.; Jiang, X.; Mehmood, K.; Iqbal, M.; Li, A.; Zhang, J.; Wang, Y.; Waqas, M.; Shen, Y.; et al. Effect of Total Flavonoids of Rhizoma drynariae on Tibial Dyschondroplasia by Regulating BMP-2 and Runx2 Expression in Chickens. Front. Pharmacol. 2018, 9, 9. [Google Scholar] [CrossRef]
- Adhikary, S.; Choudhary, D.; Ahmad, N.; Karvande, A.; Kumar, A.; Banala, V.T.; Mishra, P.R.; Trivedi, R. Dietary flavonoid kaempferol inhibits glucocorticoid-induced bone loss by promoting osteoblast survival. Nutrition 2018, 53, 64–76. [Google Scholar] [CrossRef]
- Teitelbaum, S.L. Bone resorption by osteoclasts. Science 2000, 289, 1504–1508. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.-Y.; Hwang, Y.-H.; Lee, S.-J.; Kim, J.-J.; Nam, S.-J.; Yee, S.-T. Verification of the antioxidant activity of a subterranean part of Suaeda japonica Makino. Ind. Crop. Prod. 2017, 109, 836–842. [Google Scholar] [CrossRef]
- Herald, T.J.; Gadgil, P.; Tilley, M. High-throughput micro plate assays for screening flavonoid content and DPPH-scavenging activity in sorghum bran and flour. J. Sci. Food Agric. 2012, 92, 2326–2331. [Google Scholar] [CrossRef] [PubMed]
- Grigoriadis, A.; Wang, Z.; Cecchini, M.; Hofstetter, W.; Félix, R.; Fleisch, H.; Wagner, E.F. c-Fos: A key regulator of osteoclast-macrophage lineage determination and bone remodeling. Science 1994, 266, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Mansky, K.C.; Sankar, U.; Han, J.; Ostrowski, M.C. Microphthalmia transcription factor is a target of the p38 MAPK pathway in response to receptor activator of NF-kappa B ligand signaling. J. Biol. Chem. 2002, 277, 11077–11083. [Google Scholar] [CrossRef] [PubMed]
- Gingery, A.; Bradley, E.; Shaw, A.; Oursler, M.J. Phosphatidylinositol 3-kinase coordinately activates the MEK/ERK and AKT/NFkappaB pathways to maintain osteoclast survival. J. Cell Biochem. 2003, 89, 165–179. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Harding, S.V.; Eck, P.; Thandapilly, S.J.; Gamel, T.H.; Abdel-Aal, E.-S.M.; Grow, G.H.; Tosh, S.M.; Jones, P.J.; Ames, N.P. High-Molecular-Weight beta-Glucan Decreases Serum Cholesterol Differentially Based on the CYP7A1 rs3808607 Polymorphism in Mildly Hypercholesterolemic Adults. J. Nutr. 2016, 146, 720–727. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene of Interest | Primer Sequence (5’→3’) | |
|---|---|---|
| Sense | Anti-Sense | |
| NFATc1 | GGGTCAGTGTGACCGAAGAT | GGAAGTCAGAAGTGGGTGGA |
| c-Fos | CCAGTCAAGAGCATCAGCAA | AAGTAGTGCAGCCCGGAGTA |
| Cathepsin K | GGCCAACTCAAGAAGAAAAC | GTGCTTGCTTCCCTTCTGG |
| DC-STAMP | CCAAGGAGTCGTCCATGATT | GGCTGCTTTGATCGTTTCTC |
| TRAP | GATGACTTTGCCAGTCAGCA | ACATAGCCCACACCGTTCTC |
| GAPDH | AACTTTGGCATTGTGGAAGG | ACACATTGGGGGTAGGAACA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, K.-J.; Lee, Y.; Son, S.-R.; Lee, H.; Son, Y.-J.; Lee, M.-K.; Lee, M. Water Extracts of Hull-less Waxy Barley (Hordeum vulgare L.) Cultivar ‘Boseokchal’ Inhibit RANKL-induced Osteoclastogenesis. Molecules 2019, 24, 3735. https://doi.org/10.3390/molecules24203735
Kim K-J, Lee Y, Son S-R, Lee H, Son Y-J, Lee M-K, Lee M. Water Extracts of Hull-less Waxy Barley (Hordeum vulgare L.) Cultivar ‘Boseokchal’ Inhibit RANKL-induced Osteoclastogenesis. Molecules. 2019; 24(20):3735. https://doi.org/10.3390/molecules24203735
Chicago/Turabian StyleKim, Kwang-Jin, Yongjin Lee, So-Ri Son, Hyunjin Lee, Young-Jin Son, Mi-Kyung Lee, and Mija Lee. 2019. "Water Extracts of Hull-less Waxy Barley (Hordeum vulgare L.) Cultivar ‘Boseokchal’ Inhibit RANKL-induced Osteoclastogenesis" Molecules 24, no. 20: 3735. https://doi.org/10.3390/molecules24203735
APA StyleKim, K. -J., Lee, Y., Son, S. -R., Lee, H., Son, Y. -J., Lee, M. -K., & Lee, M. (2019). Water Extracts of Hull-less Waxy Barley (Hordeum vulgare L.) Cultivar ‘Boseokchal’ Inhibit RANKL-induced Osteoclastogenesis. Molecules, 24(20), 3735. https://doi.org/10.3390/molecules24203735