Quality Formation Mechanism of Stiff Silkworm, Bombyx batryticatus Using UPLC-Q-TOF-MS-Based Metabolomics


Abstract
:
1. Introduction
2. Results
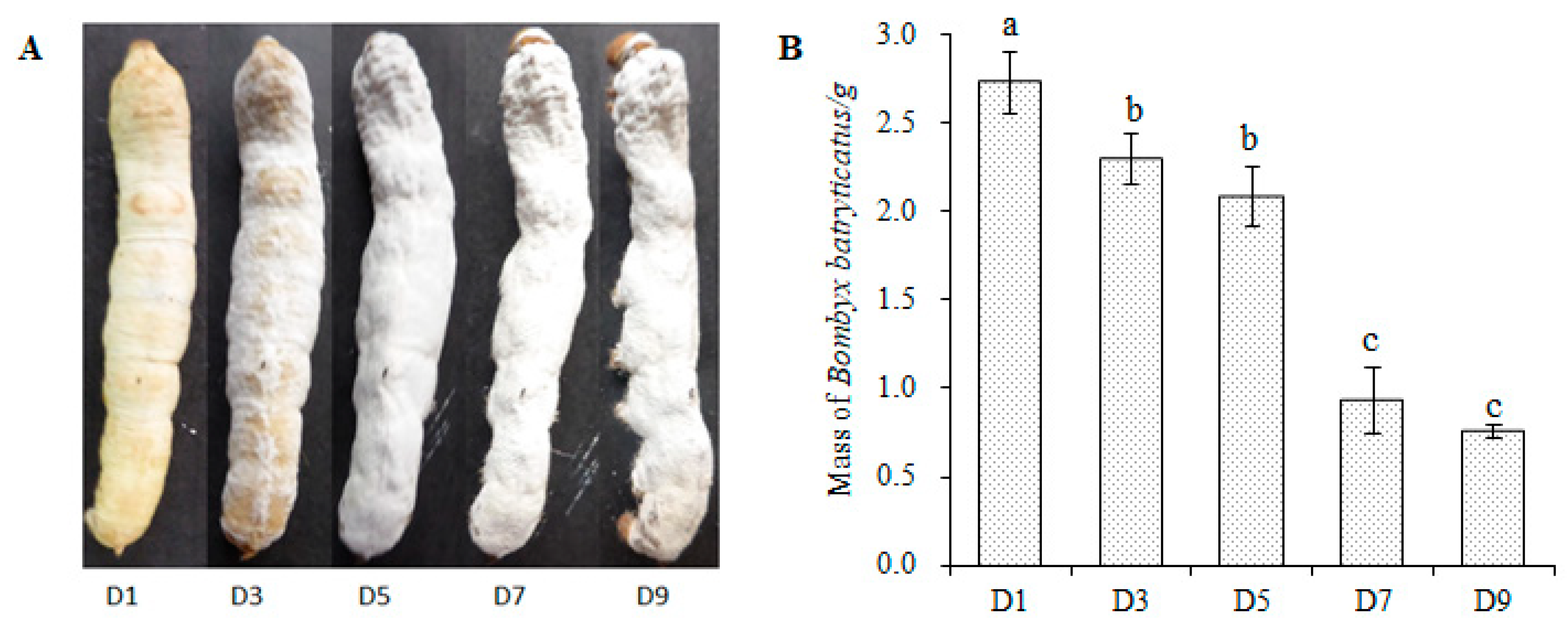
2.1. Changes in B. batryticatus during Stiff Stage
2.2. Effect of Stiff Time of B. batryticatus on Anticonvulsant Efficacy
2.3. Metabolic Profile of B. batryticatus during Stiff Stage
2.4. Analysis of Different Metabolites of B. batryticatus from Different Stiff Time
2.5. Changes in Chemical Constituents during the Stiff Stage of B. batryticatus
2.6. Screening of B. batryticatus Quality Control Index Component
3. Discussion
4. Materials and Methods
4.1. B. batryticatus
4.2. Sample Preparation
4.3. Animal Experiment
4.4. UPLC-Q/TOF-MS
4.5. Data Processing, Statistical Analysis and Metabolite Identification
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China; People’s Medical Publishing House: Beijing, China, 2015; Volume 1, p. 375. [Google Scholar]
- Namba, T. Coloured Illustrations of Wakan-Yaku; Hoikusha Publishing: Osaka, Japan, 1986; pp. 325–327. [Google Scholar]
- Lee, S.I. Clinical Application of Oriental Medicinal Herbs. Sungbo Publishing Company: Seoul, Korea, 1986; pp. 453–497. [Google Scholar]
- Pemberton, R.W. Insects and other arthropods used as drugs in Korean traditional medicine. J. Ethnopharmacol. 1999, 65, 207–216. [Google Scholar] [CrossRef]
- Yang, Q.; Liao, S.T.; Xing, D.X.; Luo, G.Q.; Wu, F.Q. Advances in chemical composition of Bombyx batryticatus and its identification techniques. Sci. Ser. 2009, 35, 696–699. [Google Scholar]
- Wang, J.X.; Zhu, C.L. The pharmacological research and clinical application of batryticated silkworms and muscardine pupae. Lishizhen Med. Mater. Med. Res. 1999, 10, 637–638. [Google Scholar]
- Kong, Y.; Xu, C.; He, Z.L.; Zhou, Q.M.; Wang, J.B.; Li, Z.Y.; Ming, X. A novel peptide inhibitor of platelet aggregation from stiff silkworm, Bombyx batryticatus. Peptides 2014, 53, 70–78. [Google Scholar] [CrossRef]
- Xu, C.; Shan, S.Y.; Liu, M.; Yang, M. Progress of researches on chemical constituents and pharmacological activities of Bombyx batryticatus. China Pharm. 2014, 25, 3732–3734. [Google Scholar]
- Tian, M.; Chen, F.; Yu, F. Progress of research on Bombyx batryticatus. Guid. J. Tradit. Chin. Med. Pharm. 2015, 21, 101–103. [Google Scholar]
- He, Q. Analysis and Comparison of Quality of Bombyx batryticatus from Different Origin. Master’s Thesis, South China Agricultural University, Guangzhou, China, 2016. [Google Scholar]
- Hu, M.B.; Yu, Z.J.; Wang, J.L.; Fan, W.X.; Liu, Y.J.; Li, J.H.; Xiao, H.; Li, Y.C.; Peng, W.; Wu, C.J. Traditional uses, origins, chemistry and pharmacology of Bombyx batryticatus: A review. Molecules 2017, 22, 1779. [Google Scholar] [CrossRef]
- Xing, D.X.; Li, L.; Liao, S.T.; Xiao, Y.; Li, Q.R.; Ye, M.Q.; Yang, Q. Identification of a Beauveria bassiana strain with high pathogenicity to Bombyx mori and its application in Bombyx Batryticatus production. Sci. Ser. 2014, 40, 879–883. [Google Scholar]
- Abdel-Halim, O.B.; Maatooq, G.T.; Marzouk, A.M. Metabolism of parthenin by Beauveria bassiana ATCC 7159. Die Pharmazie 2007, 62, 226–230. [Google Scholar]
- Herath, W.; Mikell, J.R.; Hale, A.L.; Ferreira, D.; Khan, I.A. Structure and antioxidant significance of the metabolites of 5,7-dihydroxyflavone (chrysin), and 5- and 6-hydroxyflavones. Chem. Pharm. Bull. 2008, 56, 418–422. [Google Scholar] [CrossRef]
- Yin, Z.Q.; Wang, L.; Ye, W.C.; Zhang, J.; Zhou, C.L.; Zhao, S.X. A new anthraquinone glycoside biotransformed by Beauveria bassiana. Chin. J. Nat. Med. 2008, 6, 103–104. [Google Scholar] [CrossRef]
- Holland, H.L.; Morris, T.A.; Nava, P.J.; Zabic, M. A new paradigm for biohydroxylation by Beauveria bassiana ATCC 7159. Tetrahedron 1999, 55, 7441–7460. [Google Scholar] [CrossRef]
- Grogan, G.J.; Holland, H.L. The biocatalytic reactions of Beauveria spp. J. Mol. Catal. B-Enzym. 2000, 9, 1–32. [Google Scholar] [CrossRef]
- Hou, C.X.; Qin, G.X.; Liu, T.; Geng, T.; Gao, K.; Pan, Z.H.; Qian, H.Y.; Guo, X.J. Transcriptome analysis of silkworm, Bombyx mori, during early response to Beauveria bassiana challenges. PLoS ONE 2014, 9, e91189. [Google Scholar] [CrossRef] [PubMed]
- Xing, D.X.; Yang, Q.; Jiang, L.; Li, Q.R.; Xiao, Y.; Ye, M.Q.; Xia, Q.Y. RNA-Seq analyses for two silkworm strains reveals insight into their susceptibility and resistance to Beauveria bassiana infection. Int. J. Mol. Sci. 2017, 18, 234. [Google Scholar] [CrossRef] [PubMed]
- Sisodiya, S.M.; Lin, W.R.; Harding, B.N.; Squier, M.V.; Thom, M. Drug resistance in epilepsy: Expression of drug resistance proteins in common causes of refractory epilepsy. Brain 2002, 125, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Curia, G.; Lucchi, C.; Vinet, J.; Gualtieri, F.; Marinelli, C.; Torsello, A.; Costantino, L.; Biagini, G. Pathophysiogenesis of mesial temporal lobe epilepsy: Is prevention of damage antiepileptogenic? Curr. Med. Chem. 2014, 21, 663–688. [Google Scholar] [CrossRef]
- Olivo, H.F.; Peeples, T.L.; Ríos, M.Y.; Velázquez, F.; Kim, J.W.; Narang, S. Microbial C-hydroxylation and β-4-O-methylglucosidation of methyl-benzamide 7-azanorbornane ethers with Beauveria bassiana. J. Mol. Catal. B-Enzym. 2003, 21, 97–105. [Google Scholar] [CrossRef]
- Zhan, J.; Gunatilaka, A.A.L. Selective 4’-O-methylglycosylation of the pentahydroxy-flavonoid quercetin by Beauveria bassiana ATCC 7159. Biocatal. Biotransfor. 2006, 24, 396–399. [Google Scholar] [CrossRef]
- Ibrahim, A.; Khalifa, S.I.; Khafagi, I.; Youssef, D.T.; Khan, S.; Mesbah, M. Microbial metabolism of biologically active secondary metabolites from Nerium oleander L. Chem. Pharm. Bull. 2008, 56, 1253–1258. [Google Scholar] [CrossRef]
- Guo, X.H.; Yan, Z.Y.; Liu, T.; Song, D.M.; Li, X.H. Anticonvulsive activity of three compounds isolated from Beauveria bassiana. Chin. J. Exp. Tradit. Med. Form. 2013, 19, 248–250. [Google Scholar]
- Jiang, X.; Chen, Y.; Shi, L.G. Optimization of flavonoids extraction from Bombyx batryticatus using response surface methodology and evaluation of their antioxidant and anticancer activities in vitro. Food Sci. Biotechnol. 2013, 22, 1707–1715. [Google Scholar] [CrossRef]
- Peng, Y.G.; Lei, T.X.; Fu, C.Y.; Zeng, X.Q.; Li, L.D. Effect of anticoagulant components in Bombyx batryticatus on thrombosis. Pharmacol. Clin. Chin. Mater. Med. 2007, 23, 27–29. [Google Scholar]
- Kim, Y.C.; Jung, Y.S.; Kim, S.K. Effect of betaine supplementation on changes in hepatic metabolism of sulfur-containing amino acids and experimental cholestasis induced by α-naphthylisothiocyanate. Food Chem. Toxicol. 2005, 43, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, R.; Merli, E.; Cicchitelli, G.; Mele, D.; Fucili, A.; Ceconi, C. Therapeutic effects of L-carnitine and propionyl-L-carnitine on cardiovascular diseases: A review. Ann. N. Y. Acad. Sci. 2010, 1033, 79–91. [Google Scholar] [CrossRef]
- Hu, M.B.; Liu, Y.J.; Xiao, H.; Wu, C.J.; Zhang, J.X. Advance and thinking of researches on artificial culture of Bombyx batryticatus. J. Chin. Med. Mater. 2016, 39, 930–933. [Google Scholar]
- Huang, J.M.; Su, M.S.; Zhang, Y.M.; Shao, F.; Yang, M.; Zhang, P.Z. Progress of research on phenolic transformation and 4-O-methyl-glucoside products in Bombyx bassiana. Chin. Trad. Pat. Med. 2017, 39, 1029–1033. [Google Scholar]
- Racine, R.J. Modification of seizure activity by electrical stimulation. II. Motor seizure. Electroencephalogr. Clin. Neurophysiol. 1972, 32, 281–294. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Clonic Seizure Latency/s | Tonic Seizure Latency/s | Mortality Protection Rate/% |
|---|---|---|---|
| Control | ns | ns | 100 |
| Model | 163.00 ± 12.60 | 223.90 ± 26.57 | 10 |
| SW | 177.4 ± 19.29 | 356.6 ± 160.63 | 20 |
| D1 | 187.70 ± 19.78 | 387.10 ± 157.66 | 20 |
| D5 | 222.10 ± 16.93 * | 435.80 ± 152.97 | 30 |
| D9 | 280.30 ± 21.86 ** | 684.60 ± 188.78 * | 40 |
| Classification | Metabolites | VIP Value | p-Value | FC(SW/D1) |
|---|---|---|---|---|
| Flavonoids | Unknown flavonoid | 1.93 | 3.51 × 10−3 | −6.06 |
| Rutin | 1.13 | 6.01 × 10−3 | −5.86 | |
| Unknown flavonoid | 1.8 | 1.63 × 10−2 | −4.51 | |
| Quercetin | 1.68 | 4.04 × 10−2 | −1.08 | |
| Kaempferol | 1.79 | 3.58 × 10−2 | 0.92 | |
| Amino acid | L-glutamine | 4.71 | 1.01 × 10−4 | −2.62 |
| L-asparagine | 1.18 | 3.10 × 10−3 | −2.37 | |
| L-arginine | 2.1 | 5.87 × 10−3 | −2.14 | |
| L-tryptophan | 1.95 | 4.20 × 10−3 | −1.02 | |
| L-histidine | 3.12 | 3.17 × 10−6 | −0.72 | |
| L-proline | 4.45 | 5.95 × 10−3 | −0.58 | |
| Lipids | TG (56:8) | 8.76 | 1.66 × 10−3 | −1.94 |
| TG (52:5) | 6.83 | 1.21 × 10−2 | −1.28 | |
| TG (54:6) | 7.25 | 6.51 × 10−3 | −1.24 | |
| TG (54:8) | 5.23 | 3.54 × 10−2 | −1.17 | |
| TG (56:7) | 9.91 | 1.89 × 10−2 | −1.11 | |
| TG (52:4) | 7.99 | 2.10 × 10−3 | −0.98 | |
| PC (36:2) | 0.73 | 1.38 × 10−3 | −0.96 | |
| Nucleosides | Adenine | 3.25 | 3.06 × 10−2 | −1.05 |
| dTDP | 1.37 | 8.34 × 10−4 | −0.94 | |
| Uracil | 2.45 | 1.64 × 10−4 | 1.26 | |
| Hypoxanthine | 6.23 | 1.11 × 10−2 | 2.09 | |
| Others | Benzoic acid | 1.56 | 2.88 × 10−2 | −1.39 |
| 2-Hydroxycinnamic acid | 1.73 | 3.08 × 10−2 | −1.37 | |
| 2-pyrrolidone-5-carboxylic acid | 5.79 | 2.84 × 10−4 | −1.02 | |
| Betaine | 4.53 | 3.37 × 10−3 | 0.65 | |
| Acetylcarnitine | 1.76 | 4.71 × 10−3 | 0.66 |
| Classification | Metabolites | VIP Value | p-Value | FC(D1/D5) |
|---|---|---|---|---|
| Flavonoids | Kaempferol | 1.34 | 5.31 × 10−3 | −1.82 |
| Unknown flavonoid | 1.12 | 4.26 × 10−2 | −1.25 | |
| Unknown flavonoid | 1.07 | 3.08 × 10−4 | 4.41 | |
| Kaempferol-7-O-β-d-4-O-methylglucoside | 1.39 | 8.75 × 10−5 | 6.75 | |
| Quercetin-7-O-β-d-4-O-methylglucoside | 1.67 | 1.22 × 10−6 | 6.82 | |
| Amino acid | L-histidine | 1.81 | 3.67 × 10−8 | −1.8 |
| L-phenylalanine | 3.04 | 2.59 × 10−3 | 0.57 | |
| L-isoleucine | 1.5 | 2.61 × 10−2 | 0.82 | |
| L-valine | 1.79 | 1.51 × 10−5 | 0.84 | |
| L-tryptophan | 1.39 | 6.71 × 10−6 | 1.27 | |
| L-proline | 5.38 | 1.03 × 10−6 | 3.22 | |
| Lipids | DG (34:3) | 3.2 | 3.81 × 10−4 | −2.51 |
| DG (36:6) | 5.96 | 9.74 × 10−4 | −2.27 | |
| DG (36:4) | 5.67 | 5.92 × 10−4 | −1.82 | |
| DG (34:1) | 2.69 | 4.10 × 10−4 | −1.58 | |
| DG (36:5) | 2.49 | 3.61 × 10−3 | −1.34 | |
| DG (36:3) | 2.22 | 1.72 × 10−3 | −1.1 | |
| DG (36:2) | 2.85 | 1.65 × 10−3 | −1.01 | |
| DG (33:4) | 1.96 | 6.75 × 10−3 | 1.91 | |
| DG (35:5) | 2.3 | 8.92 × 10−6 | 3.37 | |
| DG (35:6) | 1.94 | 4.41 × 10−6 | 3.43 | |
| MG (18:2) | 1.67 | 3.00 × 10−2 | −0.95 | |
| Eicosatrienoic acid (C20:3) | 1.93 | 1.71 × 10−3 | −0.93 | |
| Octadecapentaenoic acid (C18:5) | 4.89 | 3.03 × 10−2 | −3.26 | |
| Octadecatrienoic acid (C18:3) | 5.32 | 9.96 × 10−4 | −0.68 | |
| Octadecenoic acid (C18:1) | 2.98 | 4.21 × 10−2 | −0.62 | |
| Stearic acid (C18:0) | 1.96 | 3.51 × 10−3 | −2.28 | |
| Octadecatetraenoic acid (C18:4) | 3.43 | 1.31 × 10−6 | 3.9 | |
| Octadecadienoic acid (C18:2) | 1.85 | 1.10 × 10−4 | 4.33 | |
| (4E,2S,3R)-2-N-octadecanoyl- 4-tetradecasphingenine | 1.63 | 1.93 × 10−4 | −1.53 | |
| (4E,6E,2S,3R)-2-N-eicosanoyl- 4,6-tetradecasphingadienine | 1.46 | 3.05 × 10−3 | −1.07 | |
| Phytosphingosine | 3.76 | 3.41 × 10−5 | 4.64 | |
| Nucleosides | Hypoxanthine | 3.59 | 7.35 × 10−3 | −2.48 |
| Adenine | 3.41 | 2.57 × 10−3 | 2.05 | |
| Others | 2-pyrrolidone-5-carboxylic acid | 2.27 | 3.23 × 10−4 | −1.09 |
| Choline | 3.76 | 2.54 × 10−3 | −0.91 | |
| Acetylcarnitine | 1.79 | 1.80 × 10−5 | 1.03 | |
| Betaine | 6.16 | 1.79 × 10−7 | 1.48 | |
| Benzoic acid | 1.23 | 9.91 × 10−4 | 1.83 | |
| L-carnitine | 1.97 | 6.95 × 10−9 | 1.84 | |
| 2-Hydroxycinnamic acid | 1.43 | 1.10 × 10−3 | 1.9 | |
| 6-Hydroxynicotinic acid | 1.36 | 4.38 × 10−5 | 2.86 | |
| Beauverin | 10.84 | 2.04 × 10−5 | 9.36 |
| Classification | Metabolites | VIP Value | p-Value | FC(D5/D9) |
|---|---|---|---|---|
| Flavonoids | Unknown flavonoid | 1.2 | 3.05 × 10−2 | 1.39 |
| Quercetin-7-O-β-d-4-O-methylglucoside | 1.85 | 1.38 × 10−2 | 1.4 | |
| Kaempferol--7-O-β-d-4-O-methylglucoside | 1.64 | 1.68 × 10−2 | 1.47 | |
| Unknown flavonoid | 0.25 | 9.55 × 10−4 | 1.51 | |
| Amino acid | L-leucine | 2.49 | 7.59 × 10−5 | −3.75 |
| L-isoleucine | 2.66 | 1.64 × 10−5 | −3.43 | |
| L-phenylalanine | 5.61 | 1.11 × 10−7 | −2.94 | |
| L-tryptophan | 1.64 | 1.95 × 10−8 | −2.01 | |
| L-histidine | 0.62 | 3.54 × 10−3 | −0.64 | |
| L-valine | 1.56 | 6.66 × 10−4 | 0.44 | |
| Lipid | TG (54:8) | 1.68 | 1.35 × 10−2 | −3.46 |
| TG (52:6) | 3.62 | 4.51 × 10−3 | −3.73 | |
| TG (52:4) | 3.15 | 2.75 × 10−3 | −3.64 | |
| TG (56:7) | 3.56 | 3.18 × 10−3 | −3.43 | |
| TG (52:5) | 1.89 | 5.91 × 10−3 | −3.22 | |
| TG (54:6) | 2.03 | 4.74 × 10−3 | −2.75 | |
| TG (56:8) | 1.64 | 5.63 × 10−3 | −2.53 | |
| DG (35:5) | 1.8 | 9.32 × 10−4 | −1.35 | |
| DG (35:6) | 1.17 | 7.87 × 10−3 | −0.79 | |
| DG (33:4) | 1.5 | 5.56 × 10−3 | −0.76 | |
| DG (36:6) | 2.91 | 6.60 × 10−3 | 0.99 | |
| DG (36:2) | 3.95 | 2.45 × 10−5 | 1.4 | |
| DG (34:1) | 2.6 | 1.87 × 10−5 | 1.42 | |
| DG (34:3) | 2.96 | 3.16 × 10−6 | 2.17 | |
| DG (36:4) | 7.5 | 2.69 × 10−7 | 2.24 | |
| DG (36:3) | 4.48 | 1.01 × 10−6 | 2.25 | |
| DG (36:5) | 4.48 | 2.10 × 10−5 | 2.32 | |
| MG (22:6) | 1.79 | 1.60 × 10−2 | 0.44 | |
| MG (18:1) | 3.84 | 1.54 × 10−2 | 1.82 | |
| MG (20:3) | 1.95 | 8.64 × 10−5 | 1.89 | |
| MG (18:2) | 6 | 3.01 × 10−5 | 3.17 | |
| Octadecadienoic acid (C18:2) | 1.84 | 1.45 × 10−4 | −3.63 | |
| Octadecatetraenoic acid (C18:4) | 3.3 | 4.34 × 10−6 | −2.71 | |
| Eicosatetraenoic acid (C20:4) | 1.4 | 3.33 × 10−2 | −0.2 | |
| LPC (14:1) | 1.46 | 2.92 × 10−3 | 3.19 | |
| LPC (18:1) | 2.53 | 2.84 × 10−6 | 3.2 | |
| LPC (18:2) | 3.2 | 7.82 × 10−9 | 5.17 | |
| PC (36:2) | 3.07 | 2.06 × 10−6 | 6.2 | |
| PC (36:4) | 4.35 | 3.20 × 10−7 | 6.69 | |
| PC (36:5) | 3.98 | 1.85 × 10−5 | 7.04 | |
| (4E,6E,2S,3R)-2-N-eicosanoyl- 4,6-tetradecasphingadienine | 1.23 | 7.35 × 10−5 | −1.48 | |
| Phytosphingosine | 2.75 | 2.02 × 10−3 | −1.22 | |
| Nucleosides | Sphinganine | 1.36 | 1.23 × 10−2 | −1.12 |
| Xanthine | 2.45 | 2.83 × 10−3 | −4.5 | |
| Hypoxanthine | 1.43 | 2.58 × 10−2 | −2.38 | |
| Adenine | 3.32 | 3.28 × 10−3 | −1.76 | |
| Acetylcarnitine | 2.48 | 4.63 × 10−9 | −3.23 | |
| Others | 2-Hydroxycinnamic acid | 1.51 | 1.97 × 10−4 | −1.95 |
| Benzoic acid | 1.32 | 8.65 × 10−5 | −1.96 | |
| 2-pyrrolidone-5-carboxylic acid | 1.7 | 9.20 × 10−6 | −1.19 | |
| Betaine | 7.79 | 2.43 × 10−8 | 0.99 | |
| L-carnitine | 2.4 | 3.99 × 10−8 | 1.03 | |
| 6-Hydroxynicotinic acid | 2.1 | 5.97 × 10−3 | 1.78 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xing, D.; Shen, G.; Li, Q.; Xiao, Y.; Yang, Q.; Xia, Q. Quality Formation Mechanism of Stiff Silkworm, Bombyx batryticatus Using UPLC-Q-TOF-MS-Based Metabolomics. Molecules 2019, 24, 3780. https://doi.org/10.3390/molecules24203780
Xing D, Shen G, Li Q, Xiao Y, Yang Q, Xia Q. Quality Formation Mechanism of Stiff Silkworm, Bombyx batryticatus Using UPLC-Q-TOF-MS-Based Metabolomics. Molecules. 2019; 24(20):3780. https://doi.org/10.3390/molecules24203780
Chicago/Turabian StyleXing, Dongxu, Guanwang Shen, Qingrong Li, Yang Xiao, Qiong Yang, and Qingyou Xia. 2019. "Quality Formation Mechanism of Stiff Silkworm, Bombyx batryticatus Using UPLC-Q-TOF-MS-Based Metabolomics" Molecules 24, no. 20: 3780. https://doi.org/10.3390/molecules24203780
APA StyleXing, D., Shen, G., Li, Q., Xiao, Y., Yang, Q., & Xia, Q. (2019). Quality Formation Mechanism of Stiff Silkworm, Bombyx batryticatus Using UPLC-Q-TOF-MS-Based Metabolomics. Molecules, 24(20), 3780. https://doi.org/10.3390/molecules24203780