
Total Phenols from Grape Leaves Counteract Cell Proliferation and Modulate Apoptosis-Related Gene Expression in MCF-7 and HepG2 Human Cancer Cell Lines

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Yield and Total Phenolic Content
2.2. DPPH and EPR Radical-Scavenging Activity
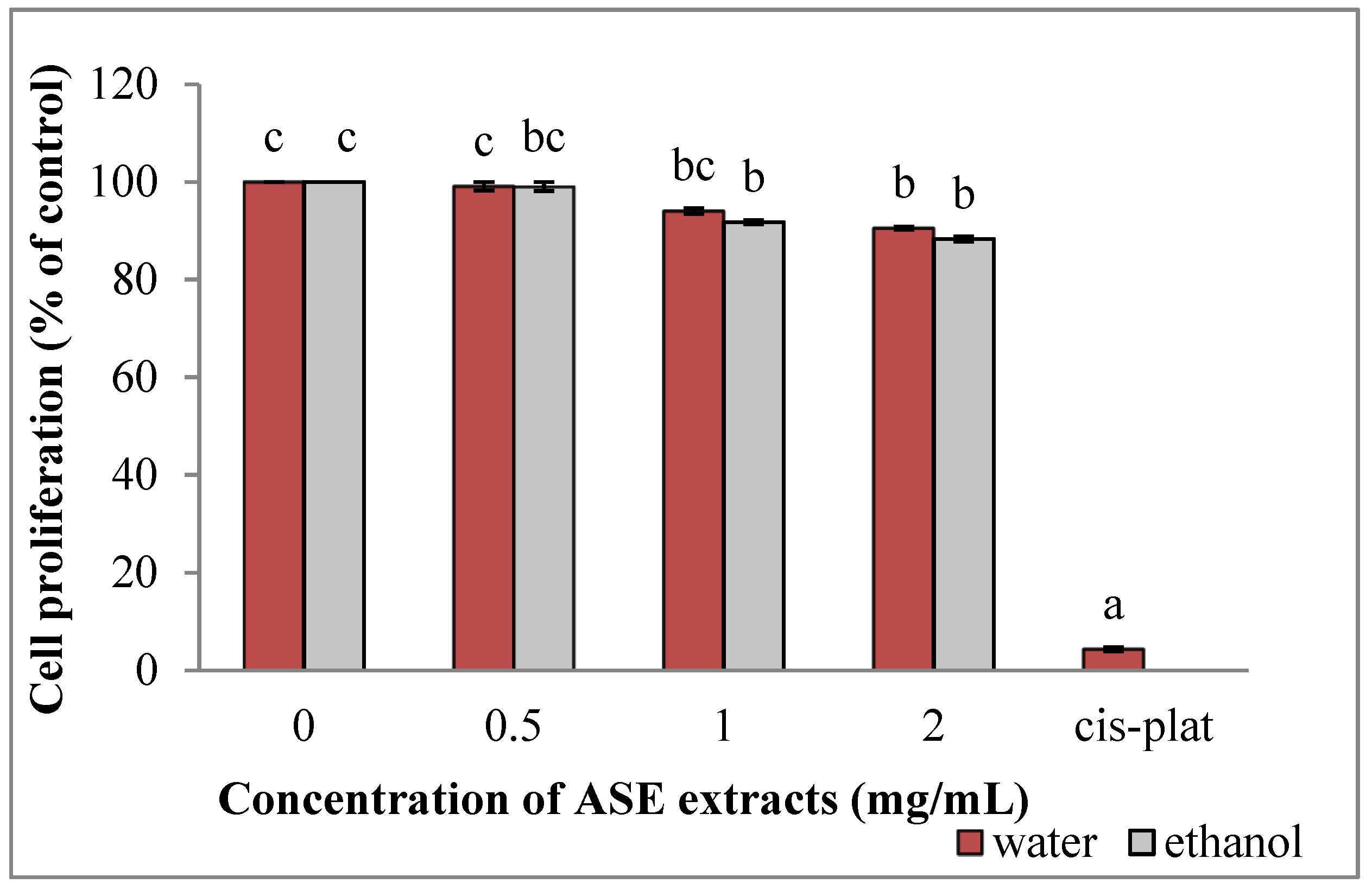
2.3. Effect of Grape Leaves EACE and WACE Extract on HUVEC Cell Proliferation
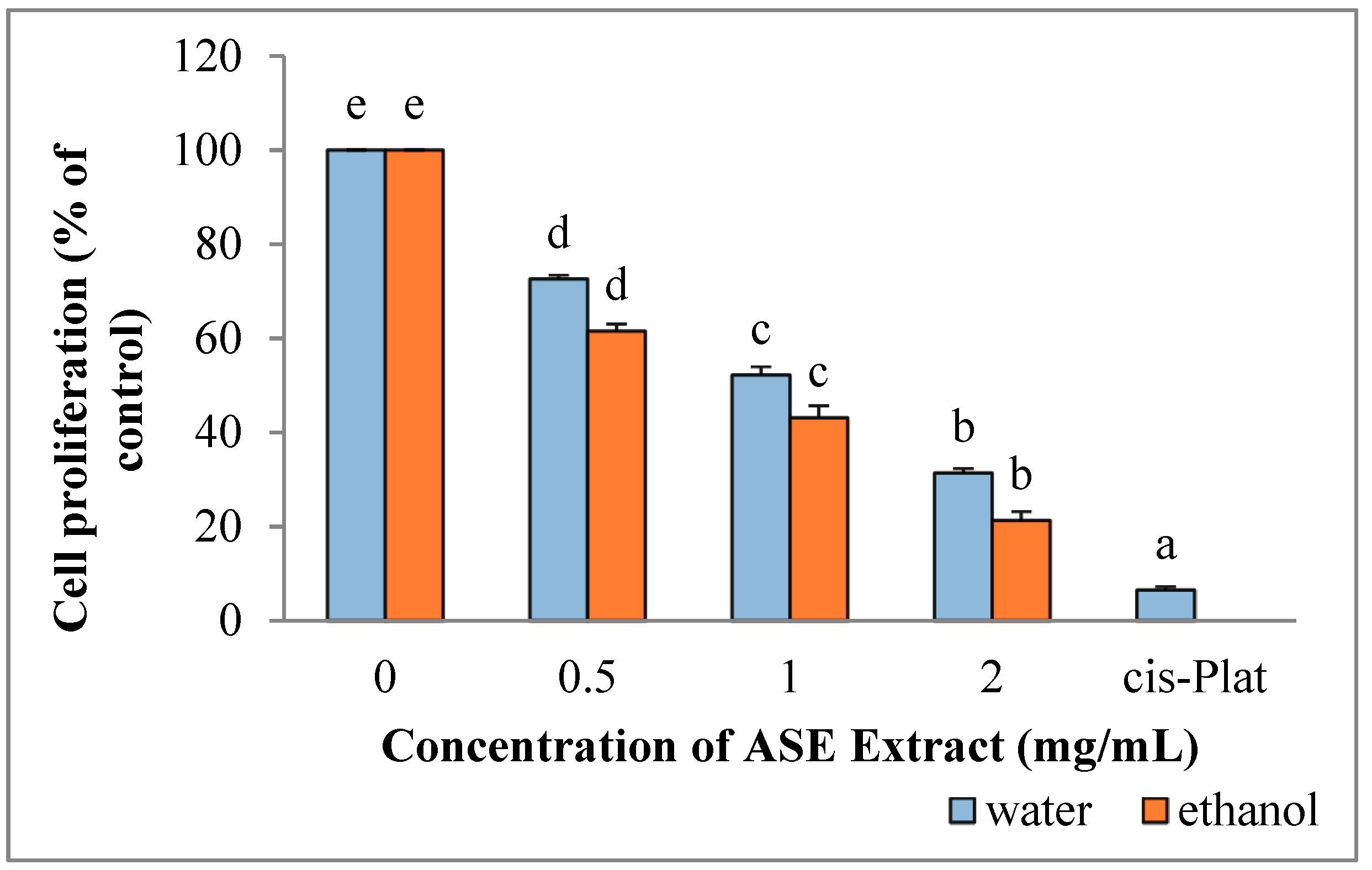
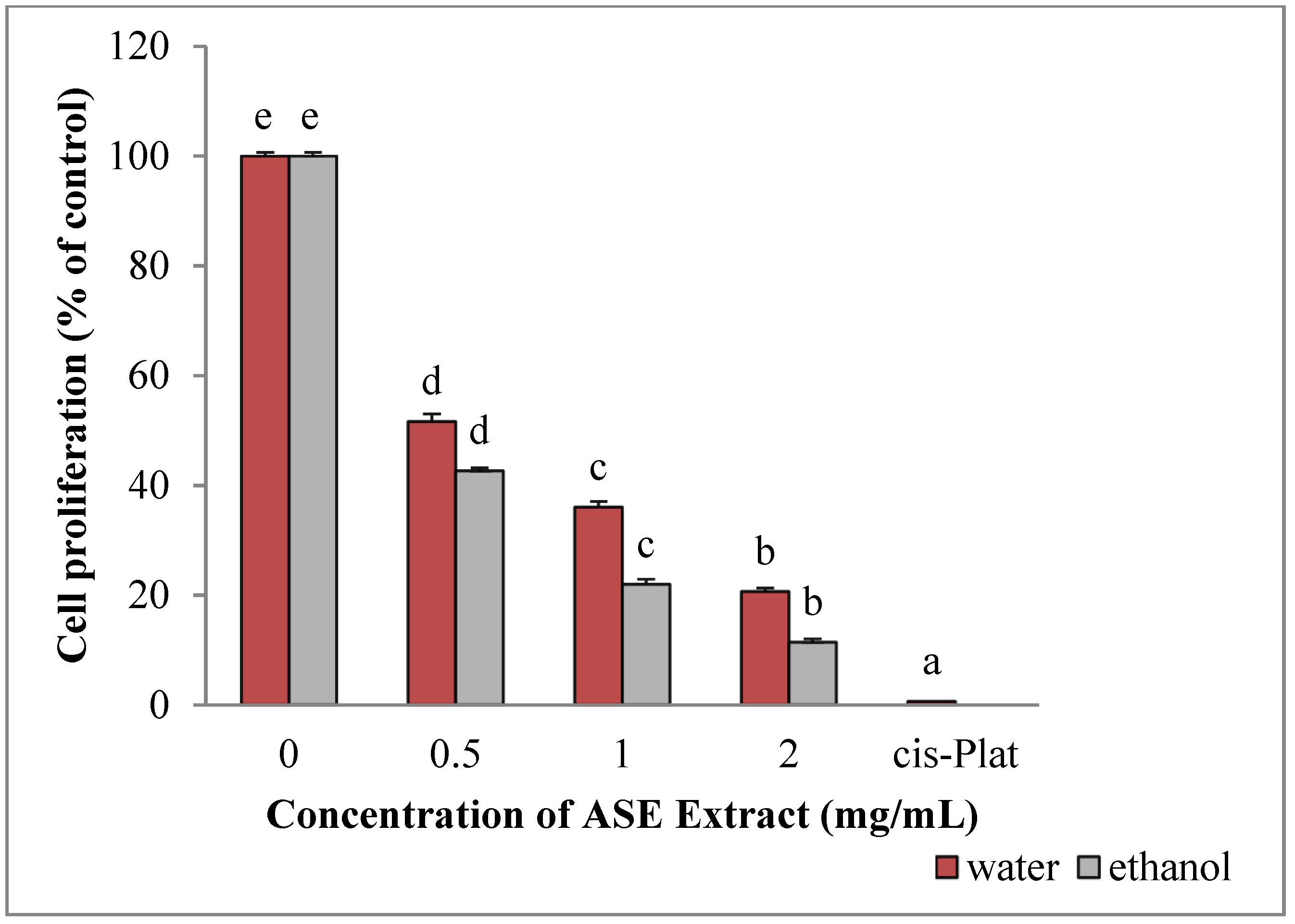
2.4. EACE and WACE Extract Counteract HepG2 Proliferation
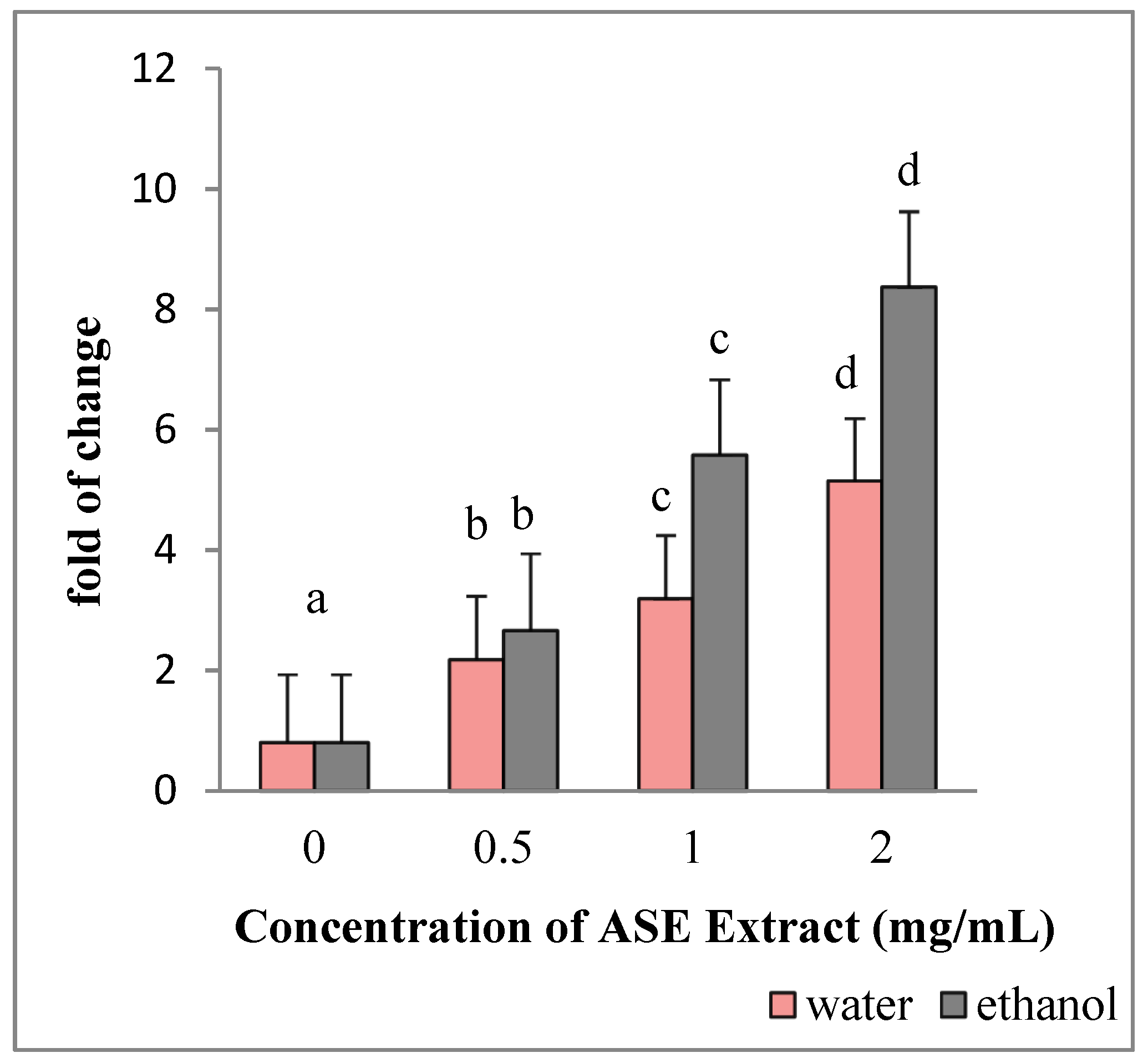
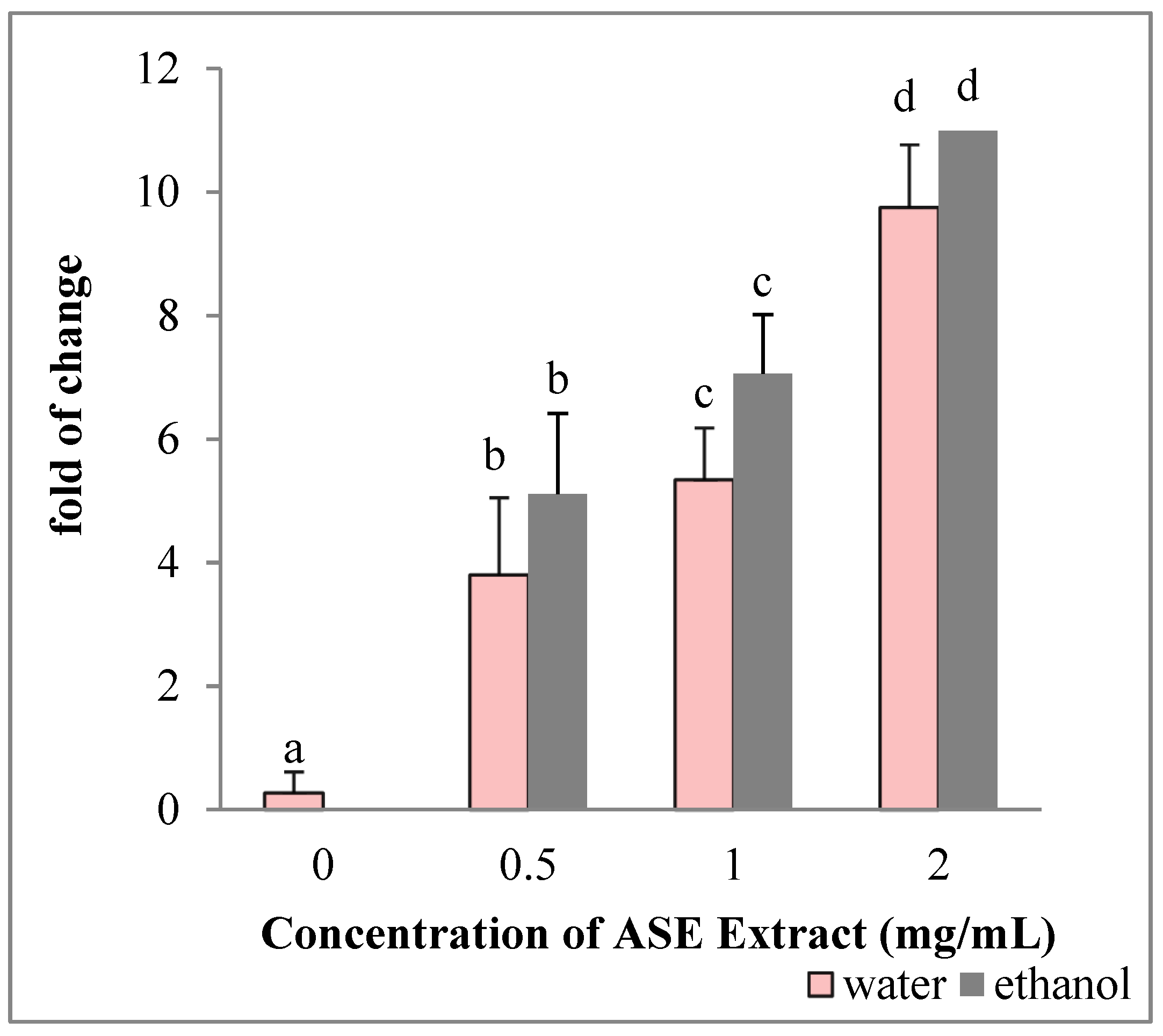
2.5. EACE and WACE Extracts Influence the Expression of Apoptosis-Related Genes in HepG2 Cells
2.6. EACE and WACE Extracts Influence MCF-7 Proliferation
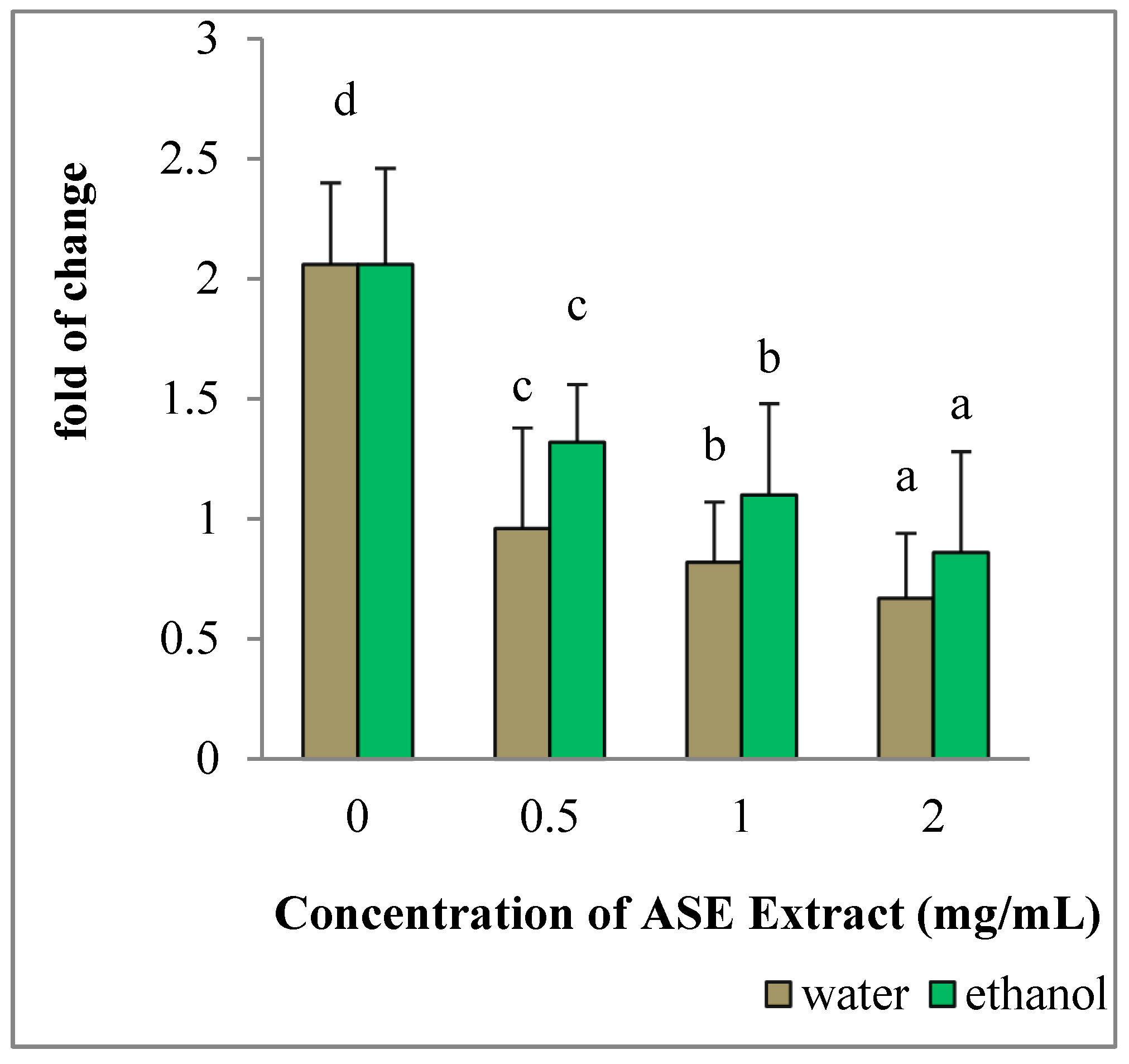
2.7. EACE and WACE Extracts Influenced the Expression Of Apoptosis-Related Genes in MCF-7 Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals and Cells
4.2. Plant Material
4.3. Extraction Procedure
4.4. Total Phenolic (TP) Content
4.5. Antioxidant Activity
4.5.1. Spin Trapping Assay of the •OH Radical
4.5.2. DPPH
4.6. Cell Culture
4.7. MTT Viability Assay
4.8. Gene Expression
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ďurackova, Z. Some Current Insights into Oxidative Stress. Physiol. Res. 2010, 59, 459–469. [Google Scholar] [PubMed]
- Barry, H. Oxidative stress and cancer: Have we moved forward? Biochem. J. 2007, 401, 1–11. [Google Scholar] [CrossRef]
- Ferlay, J.; Shin, H.R.; Bray, F.; Forman, D.; Mathers, C.; Parkin, D.M. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int. J. Cancer 2010, 127, 2893–2917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, J.S.; Jones, P.A. Cancer genetics and epigenetics: Two sides of the same coin? Cancer Cell 2012, 22, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.Y.; Hail, N., Jr.; Lotan, R. Apoptosis as a novel target for cancer chemoprevention. J. Natl. Cancer Inst. 2004, 96, 662–672. [Google Scholar] [CrossRef] [PubMed]
- Fresco, P.; Borges, F.; Marques, M.P.M.; Diniz, C. The Anticancer Properties of Dietary Polyphenols and its Relation with Apoptosis. Curr. Pharm. Des. 2010, 16, 114–134. [Google Scholar] [CrossRef] [PubMed]
- Martin, K.R. Targeting apoptosis with dietary bioactive agents. J. Med. Food 2006, 231, 117–129. [Google Scholar] [CrossRef]
- Chandrasekara Reddy, G.; Shiva Prakash, S.; Diwakar, L. Stilbene heterocycles: Synthesis, antimicrobial, antioxidant and anticancer activities. J. Pharm. Innov. 2015, 3, 24–30. [Google Scholar]
- Kalra Ekta, K. Nutraceutical-Definition and Introduction. AAPS Pharm. Sci. 2003, 5. [Google Scholar] [CrossRef]
- Basoli, V.; Santaniello, S.; Cruciani, S.; Ginesu, G.C.; Cossu, M.L.; Delitala, A.P.; Serra, P.A.; Ventura, C.; Maioli, M. Melatonin and Vitamin D Interfere with the Adipogenic Fate of Adipose-Derived Stem Cells. Int. J. Mol. Sci. 2017, 18, 981. [Google Scholar] [CrossRef]
- Santaniello, S.; Cruciani, S.; Basoli, V.; Balzano, F.; Bellu, E.; Garroni, G.; Ginesu, G.C.; Cossu, M.L.; Facchin, F.; Delitala, A.P.; et al. Melatonin and Vitamin D Orchestrate Adipose Derived Stem Cell Fate by Modulating Epigenetic Regulatory Genes. Int. J. Med. Sci. 2018, 15, 1631–1639. [Google Scholar] [CrossRef] [PubMed]
- Mazza, G.; Miniati, E. Anthocyanins in Fruits, Vegetables and Grains; CRC Press: Boca Raton, FL, USA, 1993. [Google Scholar]
- Felicio, J.D.; Santos, R.S.; Gonzalez, E. Chemical constituents from Vitis Vinifera (Vitaceae). Arg. Inst. Biol. 2001, 68, 47–50. [Google Scholar]
- Monagas, M.; Garrido, I.; Bartolomé, B.; Gomez-Cordovés, B. Chemical characterization of commercial dietary ingredients from Vitis vinifera L. Anal. Chim. Acta 2006, 563, 401–410. [Google Scholar] [CrossRef]
- Monagas, M.; Hernandez-Ledesma, B.; Gomez-Cordovés, C.; Bartolomé, B. Commercial dietary ingredients from Vitisvinifera L. leaves and grape skins: Antioxidant and chemical characterization. J. Agric. Food Chem. 2006, 54, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.J.; Weller, C.L. Recent advances in extraction of nutraceuticals from plants. Trends Food Sci. Technol. 2006, 17, 300–312. [Google Scholar] [CrossRef]
- Romanik, G.; Gilgenast, E.; Przyjazny, A.; Kamiński, M. Techniques of preparing plant material for chromatographic separation and analysis. J. Biochem. Biophys. Methods 2007, 70, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.; Russell, J.M. Plant Phenolics: Extraction, Analysis and Their Antioxidant and Anticancer Properties. Moeclues 2010, 15, 7313–7352. [Google Scholar] [CrossRef] [Green Version]
- Asma, H.A.-S.; Mohammad, A.H. Total phenols, total flavonoids contents and free radical scavenging activity of seeds crude extracts of pigeon pea traditionally used in Oman for the treatment of several chronic diseases. Asian Pac. J. Trop. Dis. 2015, 5, 316–321. [Google Scholar] [CrossRef]
- Pari, L.; Suresh, A. Effect of grape (Vitisvinifera L.) leaf extract on alcohol induced oxidative stress in rats. Food Chem. Toxic. 2008, 46, 1627–1634. [Google Scholar] [CrossRef]
- Deliorman, O.D.; Orhan, N.; Ergun, E.; Ergun, F. Hepatoprotective effect of Vitisvinifera L. leaves on carbon tetrachloride-induced acute liver damage in rats. J. Ethnopharmacol. 2007, 112, 145–151. [Google Scholar] [CrossRef]
- Richter, B.E.; Jones, B.A.; Ezzell, J.L.; Porter, N.L.; Avdalovic, N.; Pohl, C. Accelerated solvent extraction: A technology for sample preparation. Anal. Chem. 1996, 68, 1033–1039. [Google Scholar] [CrossRef]
- Brachet, A.; Rudaz, S.; Mateus, L.; Christen, P.; Veuthey, J. Optimisation of accelerated solvent extraction of cocaine and benzoylecgonine from coca leaves. J. Sep. Sci. 2001, 24, 865–873. [Google Scholar] [CrossRef]
- Kaufmann, B.; Christen, P.; Veuthey, J.L. Study of factors influencing pressurized solvent extraction of polar steroids from plant material Application to the Recovery of Withanolides. Chromatographia 2001, 54, 394–398. [Google Scholar] [CrossRef]
- Ju, Z.Y.; Howard, L.R. Effects of solvent and temperature on pressurized liquid extraction of anthocyanins and total phenolics from dried red grape skin. J. Agric. Food Chem. 2003, 51, 5207–5213. [Google Scholar] [CrossRef] [PubMed]
- Pineiro, Z.; Palma, M.; Barroso, C.G. Determination of trans-resveratrol in grapes by pressurised liquid extraction and fast high-performance liquid chromatography. J. Chromatogr. A 2006, 1110, 61–65. [Google Scholar] [CrossRef]
- Luque-Rodriguez, J.M.; Luque de Castro, M.D.; Perez-Juan, P. Dynamic superheated liquid extraction of anthocyanins and other phenolics from red grape skins of winemaking residues. Bioresour. Technol. 2007, 98, 2705–2713. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Salces, R.M.; Korta, E.; Barranco, A.; Berrueta, L.A.; Gallo, B.; Vicente, F. Pressurized liquid extraction for the determination of polyphenols in apple. J. Chromatogr. A 2001, 933, 37–43. [Google Scholar] [CrossRef]
- Howard, L.; Pandjaitan, N. Pressurized liquid extraction of flavonoids from spinach. J. Food Sci. 2008, 73, 151–157. [Google Scholar] [CrossRef]
- Luthria, D.L.; Mukhopadhyay, S. Influence of sample preparation on assay of phenolic acids from eggplant. J. Agric. Food Chem. 2006, 54, 41–47. [Google Scholar] [CrossRef]
- Bonoli, M.; Marconi, E.; Caboni, M.F. Free and bound phenolic compounds in barley (Hordeumvulgare L.) flours. J. Chromatogr. A 2004, 1057, 1–12. [Google Scholar] [CrossRef]
- Hawkins, C.L.; Davies, M.J. Detection and characterization of radicals in biological materials using EPR methodology. Biochim. Biophys. Acta 2014, 1840, 708–721. [Google Scholar] [CrossRef] [PubMed]
- Goupy, P.; Dufour, C.; Loonis, M.; Dangles, O. Quantitative kinetic analysis of hydrogen transfer reactions from dietary polyphenols to the DPPH radical. J. Agric. Food Chem. 2003, 51, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Fadda, A.; Barberis, A.; Sanna, D. Influence of pH, buffers and role of quinolinic acid, a novel iron chelating agent, in the determination of hydroxyl radical scavenging activity of plant extracts by Electron Paramagnetic Resonance (EPR). Food Chem. 2018, 240, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Samoticha, J.; Wojdyło, A.; Golis, T. Phenolic composition, physicochemical properties and antioxidant activity of interspecific hybrids of grapes growing in Poland. Food Chem. 2017, 215, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Farhadi, K.; Esmaeilzadeh, F.; Hatami, M.; Forough, M.; Molaie, R. Determination of phenolic compounds content and antioxidant activity in skin, pulp, seed, cane and leaf of five native grape cultivars in West Azerbaijan province. Iran. Food Chem. 2016, 15, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free-radical method to evaluate antioxidant activity. Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Tan, J.; Wang, B.C.; Zhu, L.C. Regulation of Survivin and Bcl-2 in HepG2 Cell Apoptosis Induced by Quercetin. Chem. Biodevers. 2009, 6, 1101–1110. [Google Scholar] [CrossRef]
- Farahmandzad, F.; Ahmadi, R.; Riyahi, N.; Mahdavi, E. The Effects of Scrophulariastriata extract on Apoptosis in Glioblastoma Cells in Cell Culture. Int. J. Adv. Chem. Eng. Bio. Sci. 2015, 2, 62–63. [Google Scholar] [CrossRef]
- Morak-Młodawska, B.; Pluta, K.; Latocha, M.; Suwińska, K.; Jeleń, M.; Kuśmierz, D. 3,6-Diazaphenothiazines as potential lead molecules – synthesis, characterization and anticancer activity. J. Enzyme Inhib. Med. Chem. 2016, 31, 1512–1519. [Google Scholar] [CrossRef]
- Morak-Młodawska, B.; Pluta, K.; Latocha, M.; Jelén, M.; Kùsmierz, D. Synthesis, Anticancer Activity, and Apoptosis Induction of Novel 3,6-Diazaphenothiazines. Molecules 2019, 24, 267. [Google Scholar] [CrossRef]
- Deepak, B.; Animesh, M.; Eviatar, N.; Anupam, B. Apoptosis-inducing effects of extracts from desert plants in HepG2 human hepatocarcinoma cells. Asian Pac. J. Trop. Biomed. 2015, 5, 87–92. [Google Scholar] [CrossRef] [Green Version]
- Khazaei, S.; Abdul Hamid, R.; MohdEsa, N.; Ramachandran, V.; GhomiTabatabaee, F.; Aalam, A.; Ismail, P. Promotion of HepG2 cell apoptosis by flower of Allium atroviolaceum and the mechanism of action. BMC Complement. Altern. Med. 2017, 17, 1594–1596. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhang, B.Y.; Jia, Z.X.; Wu, W.J.; Lu, Z.Q. Hepatocellular carcinoma HepG2 cell apoptosis and caspase-8 and Bcl-2 expression induced by injectable seed extract of Coixlacryma-jobi. Hepatob. Pancreat. Dis. Int. 2011, 10, 303–307. [Google Scholar] [CrossRef]
- Blassan, P.G.; Abrahamse, H.; Parimelazhagan, T. Caspase dependent apoptotic activity of Rubus fairholmianus Gard.on MCF-7 human breast cancer cell lines. J. Appl. Biomed. 2016, 89, 1–9. [Google Scholar] [CrossRef]
- Filipa, S.; Reis, D.S.; Lillian, B.; Anabela, M.; Patricia, M.; Isabel, C.F.R.; Vasconcelos, M.H. Leccinumvulpinum Watling induces DNA damage, decreases cell proliferation and induces apoptosis on the human MCF-7 breast cancer cell line. Food Chem. Toxicol. 2013, 90, 45–54. [Google Scholar] [CrossRef]
- Dikmen, M.; Ozturk, N.; Ozturk, Y. The Antioxidant Potency of Punicagranatum, L. Fruit Peel Reduces Cell Proliferation and Induces Apoptosis on Breast Cancer. J. Med. Food. 2011, 14, 1638–1646. [Google Scholar] [CrossRef] [PubMed]
- Atmaca, H.; Bozkurt, E. Apoptotic and anti-angiogenic effects of Salvia triloba extract in prostate cancer cell lines. Tumor Biol. 2015, 37, 3639–4657. [Google Scholar] [CrossRef] [PubMed]
- Amirala, A.; Speidel, V.H.; Abbas, D.; Nader, K.; Gharavi, F.; Zare, S.; Mona, O.; Ali, B.; Behzad, B. Induction of apoptosis by grape seed extract (Vitisvinifera) in oral squamous cell carcinoma. Bosn. J. Basic Med. Sci. 2013, 13, 186–191. [Google Scholar] [CrossRef]
- Maioli, M.; Basoli, V.; Carta, P.; Fabbri, D.; Antonietta, D.M.; Cruciani, S.; Andrea, S.P.; Delogu, G. Synthesis of magnolol and honokiol derivatives and their effect against hepatocarcinoma cells. PLoS ONE 2018, 13, e0192178. [Google Scholar] [CrossRef]
- Zhou, K.Q.; Julian, J.R. Potential Anticancer Properties of Grape Antioxidants. J. Oncol. 2012. [Google Scholar] [CrossRef]
- Folin, D.; Denis, W. On phosphotungstic-phosphomolybdic compounds as color reagents. J. Bio. Chem. 1912, 12, 239–243. [Google Scholar]
- Folin, D.; Ciocalteu, V. On tyrosine and tryptophan determinations in proteins. J. Bio. Chem. 1927, 73, 627–650. [Google Scholar]
- Singleton, V.; Rossi, J.A., Jr. Colorimetry of total phenolic with phosphotungstic-phosphomolybdic acid reagents. Am. J. Enol. Viticult. 1965, 16, 144–157. [Google Scholar]
- Fadda, A.; Bernardo, P.; Alberto, A.; Antonio, B.; Maria, C. Suitability for ready-to-eat processing and preservation of six green and red baby leaves cultivars and evaluation of their antioxidant value during storage and after the expiration date. J. Food Process Preserv. 2015, 40, 550–558. [Google Scholar] [CrossRef]
- Choi, C.W.; Kim, S.C.; Hwang, S.S.; Choi, B.K. Antioxidant activity and free radical scavenging capacity between Korean medicinal plants and flavonoids by assay guided comparison. Plant Sci. 2002, 163, 1161–1168. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Kenneth, J.L.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds from grape leaves are available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Extracts | Total Phenols (mg GAE/gr DW ± SD) y | Yield (% ± SD) | IC50·OH (mg/mL ± SD) | IC50 DPPH (mg/mL ± SD) |
|---|---|---|---|---|
| WACE | 55.41 ± 0.11 a | 22.8 ± 3.21 a | 0.67 ± 0.53 a R2 = 0.9791 | 0.15 ± 0.41 a R2 = 0.9711 |
| EACE | 155.73 ± 1.20 b | 18.87 ± 0.6 a | 0.64 ± 0.71 a R2 = 0.9989 | 0.09 ± 0.32 b R2 = 09922 |
| Extract | MCF-7 | HepG2 | HUVEC | |
|---|---|---|---|---|
| Cells | ||||
| WACE y IC50* (mg/mL) | 0.71 | 1.1 | >>2 | |
| EACE x IC50* (mg/mL) | 0.43 | 0.7 | >>2 | |
| Primers | Forward | Reverse |
|---|---|---|
| hGAPDH | GAGTCAACGGATTTGGTCGT | GACAAGCTTCCCGTTCTCAG |
| BAX | TCTGACGGCAACTTCAACTG | TTGAGGAGTCTCACCCAACC |
| BCL-2 | AGGATTGTGGCCTTCTTTGA | ACAGTTCCACAAAGGCATCC |
| Temperature (°C) | 40 |
|---|---|
| Pressure (PSI) | 1500 |
| Number of Cycle | 2 |
| Extraction time of one cycle (min) | 5 |
| Concentration of Ethanol (%) | 60 Ethanol/40 water |
| Type of water used | Ultrapure |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferhi, S.; Santaniello, S.; Zerizer, S.; Cruciani, S.; Fadda, A.; Sanna, D.; Dore, A.; Maioli, M.; D’hallewin, G. Total Phenols from Grape Leaves Counteract Cell Proliferation and Modulate Apoptosis-Related Gene Expression in MCF-7 and HepG2 Human Cancer Cell Lines. Molecules 2019, 24, 612. https://doi.org/10.3390/molecules24030612
Ferhi S, Santaniello S, Zerizer S, Cruciani S, Fadda A, Sanna D, Dore A, Maioli M, D’hallewin G. Total Phenols from Grape Leaves Counteract Cell Proliferation and Modulate Apoptosis-Related Gene Expression in MCF-7 and HepG2 Human Cancer Cell Lines. Molecules. 2019; 24(3):612. https://doi.org/10.3390/molecules24030612
Chicago/Turabian StyleFerhi, Selma, Sara Santaniello, Sakina Zerizer, Sara Cruciani, Angela Fadda, Daniele Sanna, Antonio Dore, Margherita Maioli, and Guy D’hallewin. 2019. "Total Phenols from Grape Leaves Counteract Cell Proliferation and Modulate Apoptosis-Related Gene Expression in MCF-7 and HepG2 Human Cancer Cell Lines" Molecules 24, no. 3: 612. https://doi.org/10.3390/molecules24030612
APA StyleFerhi, S., Santaniello, S., Zerizer, S., Cruciani, S., Fadda, A., Sanna, D., Dore, A., Maioli, M., & D’hallewin, G. (2019). Total Phenols from Grape Leaves Counteract Cell Proliferation and Modulate Apoptosis-Related Gene Expression in MCF-7 and HepG2 Human Cancer Cell Lines. Molecules, 24(3), 612. https://doi.org/10.3390/molecules24030612