
Small Molecule Fluorescent Probes for G- Quadruplex Visualization as Potential Cancer Theranostic Agents

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
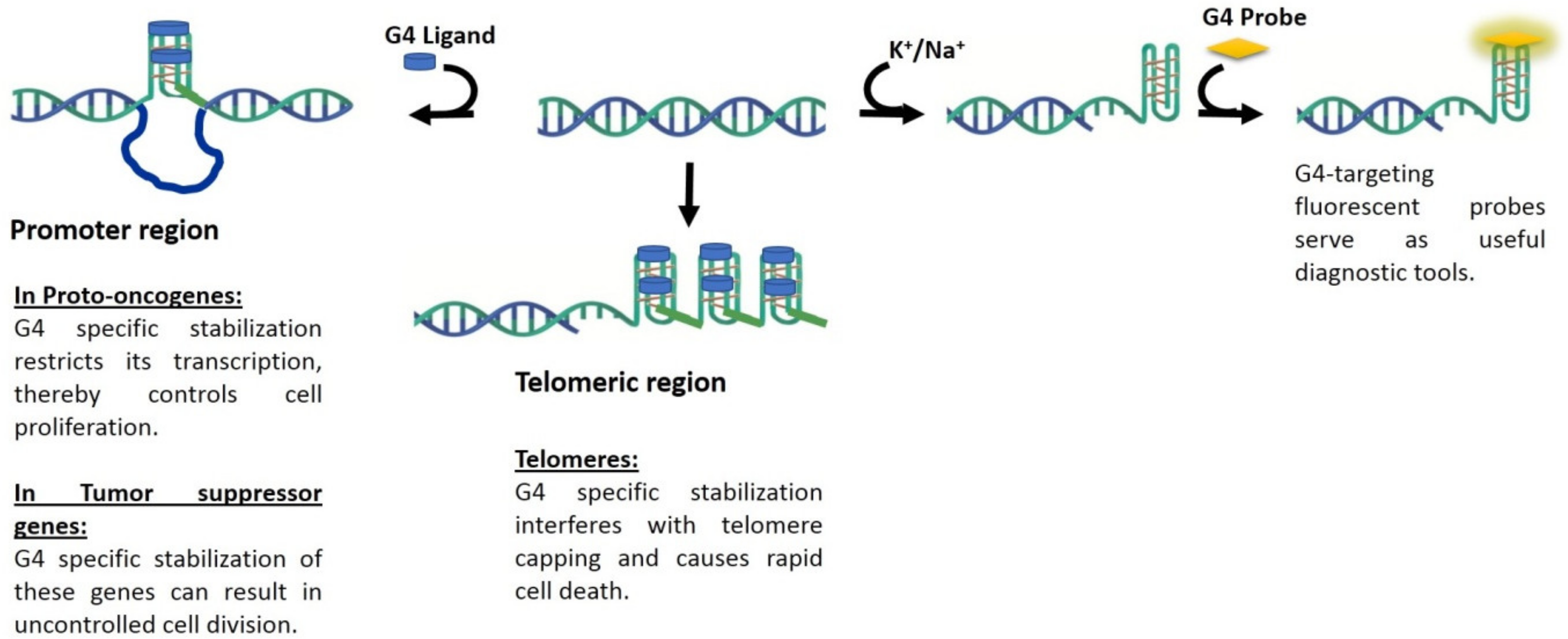
:1. G-Quadruplexes (G4) and Cancer
1.1. G4 as Molecular Targets
1.2. G4 Stabilizing Agents for Cancer Therapeutics
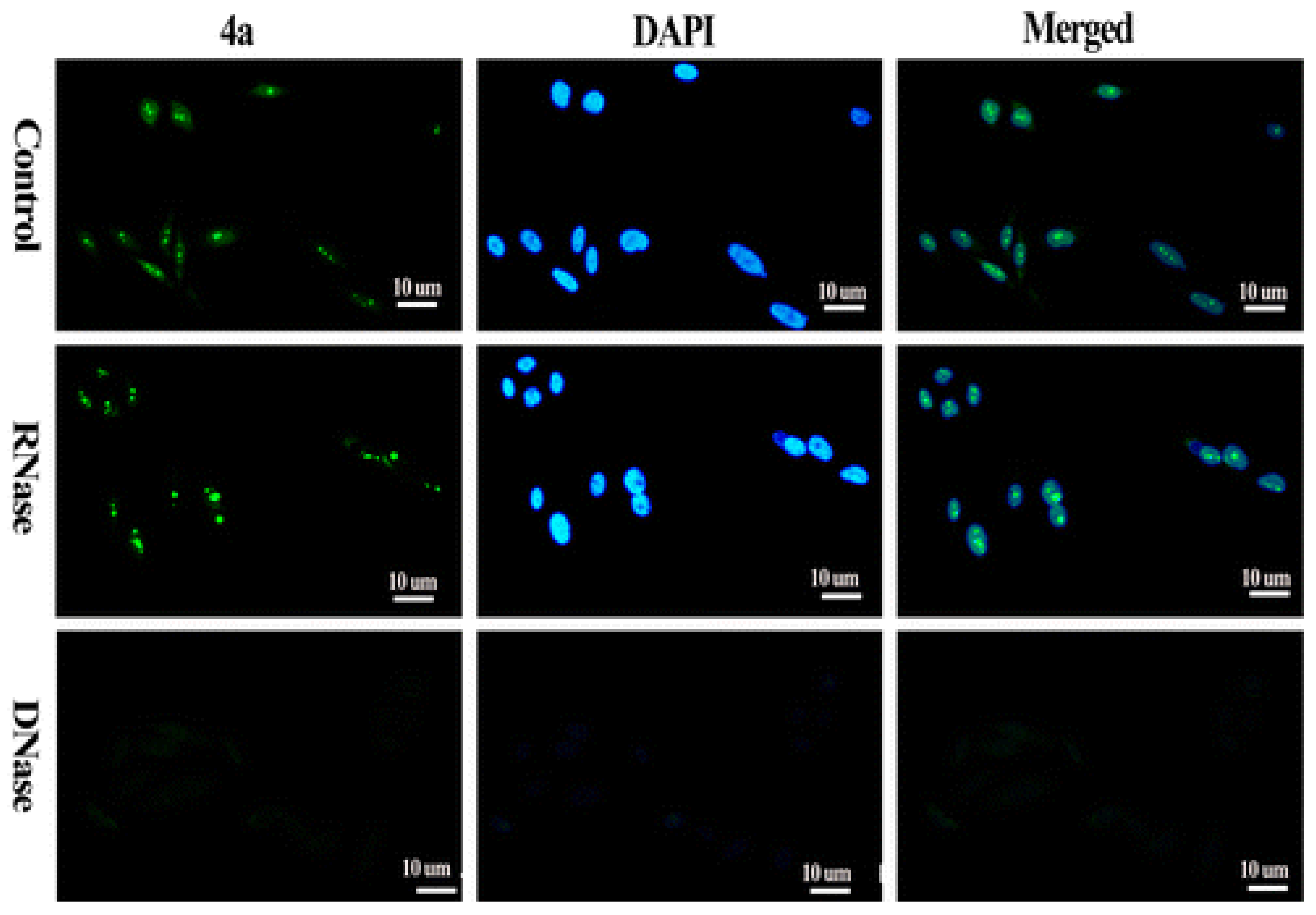
1.3. Visualizing G4 Structures in Cellular Context
2. Fluorescence Probes for G4 Visualization
2.1. BMVC
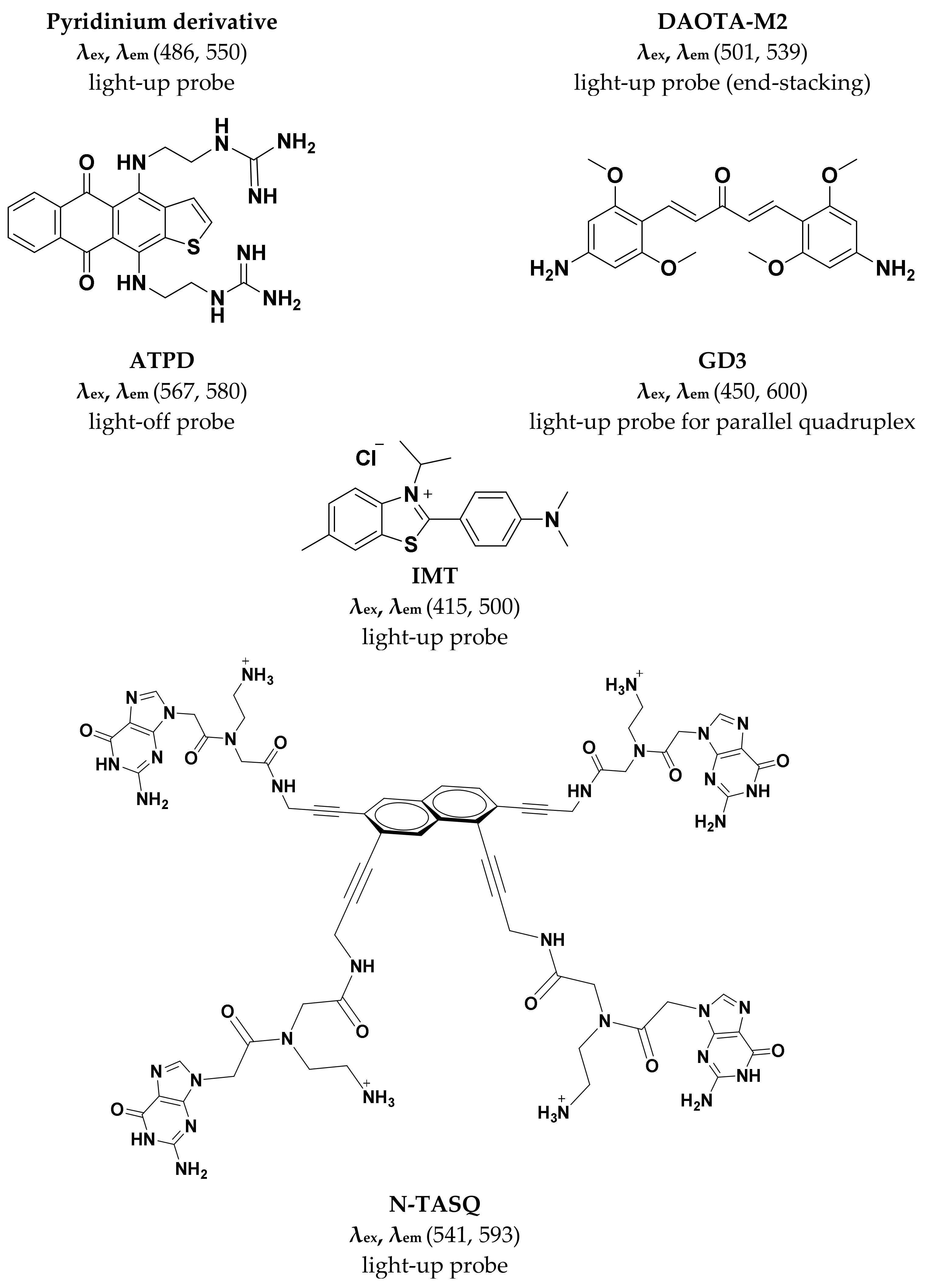
2.2. Pyridinium Derivatives
2.3. Thiazole Orange and Analogues
2.4. IMT
2.5. N-TASQ
2.6. GD3
2.7. Other Agents
3. Conclusions and Perspectives
Abbreviations of G4 Selective Fluorescence Probes
| BMVC | 3,6-bis(1-methyl-4-vinylpyridinium)carbazolediiodide |
| TO | Thiazole Orange |
| IMT | N-Isopropyl-2-(4-N,N-dimethylanilino)-6-methylbenzothiazole |
| N-TASQ | Naptho-Template Assisted Synthetic G-Quartet |
| GD3 | Tetra-methoxy bis(4-aminobenzylidene) acetone |
| DAOTA-M2 | Morpholino containing bis-substituted triangulenium |
| ADOTA-M | Morpholino containing mono-substituted triangulenium |
| BPBC | 9-methyl-3,6-bis[5-(4-methylpiperazin-1-yl)-1Hbenzo[d]imidazol-2-yl]-9H-carbazole |
| ATPDs | Guanidino anthrathiophenediones |
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Kastan, M.B.; Bartek, J. Cell-cycle checkpoints and cancer. Nature 2004, 432, 316. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, S.; Hurley, L.H.; Neidle, S. Targeting g-quadruplexes in gene promoters: A novel anticancer strategy? Nat. Rev. Drug Discov. 2011, 10, 261. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Hurley, L.H. G-quadruplex DNA: A potential target for anti-cancer drug design. Trends Pharmacol. Sci. 2000, 21, 136–142. [Google Scholar] [CrossRef]
- Murat, P.; Balasubramanian, S. Existence and consequences of g-quadruplex structures in DNA. Curr. Opin. Genet. Dev. 2014, 25, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Kwok, C.K.; Marsico, G.; Sahakyan, A.B.; Chambers, V.S.; Balasubramanian, S. Rg4-seq re-veals widespread formation of g-quadruplex structures in the human transcriptome. Nat. Methods 2016, 13, 841. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.Y.; Lejault, P.; Chevrier, S.; Boidot, R.; Robertson, A.G.; Wong, J.M.Y.; Monchaud, D. Transcriptome-wide identification of transient RNA g-quadruplexes in human cells. Nat. Commun. 2018, 9, 4730. [Google Scholar] [CrossRef] [PubMed]
- Eddy, J.; Maizels, N. Gene function correlates with potential for g4 DNA formation in the human genome. Nucleic Acids Res. 2006, 34, 3887–3896. [Google Scholar] [CrossRef]
- Huppert, J.L. Hunting g-quadruplexes. Biochimie 2008, 90, 1140–1148. [Google Scholar] [CrossRef]
- Sun, D.; Thompson, B.; Cathers, B.E.; Salazar, M.; Kerwin, S.M.; Trent, J.O.; Jenkins, T.C.; Neidle, S.; Hurley, L.H. Inhibition of human telomerase by a g-quadruplex-interactive com-pound. J. Med. Chem. 1997, 40, 2113–2116. [Google Scholar] [CrossRef]
- Siddiqui-Jain, A.; Grand, C.L.; Bearss, D.J.; Hurley, L.H. Direct evidence for a g-quadruplex in a promoter region and its targeting with a small molecule to repress c-myc transcription. Proc. Natl. Acad. Sci USA 2002, 99, 11593–11598. [Google Scholar] [CrossRef]
- Cimino-Reale, G.; Zaffaroni, N.; Folini, M. Emerging role of g-quadruplex DNA as target in anticancer therapy. Curr. Pharm. Des. 2016, 22, 6612–6624. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Chen, M.; Wu, Y.L.; Tanaka, Y.; Ji, Y.J.; Zhang, S.L.; Wei, C.H.; Xu, Y. Formation and stabilization of the telomeric antiparallel g-quadruplex and inhibition of telomerase by novel benzothioxanthene derivatives with anti-tumour activity. Sci. Rep. 2015, 5, 13693. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-Y.; Vankayalapati, H.; Shin-Ya, K.; Wierzba, K.; Hurley, L.H. Telomestatin, a potent telomerase inhibitor that interacts quite specifically with the human telomeric intramolecu-lar g-quadruplex. J. Am. Chem. Soc. 2002, 124, 2098–2099. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.; Agarwala, P.; Maiti, S. Targeting RNA g-quadruplexes for potential therapeutic applications. In RNA Therapeutics; Garner, A., Ed.; Springer: Cham, Switzerland, 2017; Volume 27. [Google Scholar]
- Tateishi-Karimata, H.; Kawauchi, K.; Sugimoto, N. Destabilization of DNA g-quadruplexes by chemical environment changes during tumor progression facilitates transcription. J. Am. Chem. Soc. 2018, 140, 642–651. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.J.; Phan, A.T.P.; Kuryavyi, V. Human telomere, oncogenic promoter and 5′-utr g-quadruplexes: Diverse higher order DNA and RNA targets for cancer therapeutics. Nucleic Acids Res. 2007, 35, 7429–7455. [Google Scholar] [CrossRef] [PubMed]
- Eser, S.; Schnieke, A.; Schneider, G.; Saur, D. Oncogenic kras signalling in pancreatic cancer. Br. J. Cancer 2014, 111, 817–822. [Google Scholar] [CrossRef] [PubMed]
- Campani, D.; Esposito, I.; Boggi, U.; Cecchetti, D.; Menicagli, M.; De Negri, F.; Colizzi, L.; Del Chiaro, M.; Mosca, F.; Fornaciari, G.; et al. Bcl-2 expression in pancreas development and pancreatic cancer progression. J. Pathol. 2001, 194, 444–450. [Google Scholar] [CrossRef]
- Todd, A.K.; Johnstone, M.; Neidle, S. Highly prevalent putative quadruplex sequence mo-tifs in human DNA. Nucleic Acids Res. 2005, 33, 2901–2907. [Google Scholar] [CrossRef]
- Neidle, S. Quadruplex nucleic acids as novel therapeutic targets. J. Med. Chem. 2016, 59, 5987–6011. [Google Scholar] [CrossRef]
- Marchetti, C.; Zyner, K.G.; Ohnmacht, S.A.; Robson, M.; Haider, S.M.; Morton, J.P.; Marsi-co, G.; Vo, T.; Laughlin-Toth, S.; Ahmed, A.A.; et al. Targeting multiple effector pathways in pancreatic ductal adenocarcinoma with a g-quadruplex-binding small molecule. J. Med. Chem. 2018, 61, 2500–2517. [Google Scholar] [CrossRef]
- Zhou, G.; Liu, X.; Li, Y.; Xu, S.; Ma, C.; Wu, X.; Cheng, Y.; Yu, Z.; Zhao, G.; Chen, Y. Telomere targeting with a novel g-quadruplex-interactive ligand braco-19 induces t-loop disassembly and telomerase displacement in human glioblastoma cells. Oncotarget 2016, 7, 14925. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.-H.; Gu, L.-Q.; Wu, J.-Y. Design of selective g-quadruplex ligands as potential anti-cancer agents. Mini Rev. Med. Chem. 2008, 8, 1163–1178. [Google Scholar] [CrossRef] [PubMed]
- Ruggiero, E.; Richter, S.N. G-quadruplexes and g-quadruplex ligands: Targets and tools in antiviral therapy. Nucleic Acids Res. 2018, 46, 3270–3283. [Google Scholar] [CrossRef] [PubMed]
- Salgado, G.F.; Cazenave, C.; Kerkour, A.; Mergny, J.L. G-quadruplex DNA and ligand inter-action in living cells using nmr spectroscopy. Chem. Sci. 2015, 6, 3314–3320. [Google Scholar] [CrossRef] [PubMed]
- Largy, E.; Granzhan, A.; Hamon, F.; Verga, D.; Teulade-Fichou, M.P. Visualizing the quadru-plex: From fluorescent ligands to light-up probes. Top. Curr. Chem. 2013, 330, 111–177. [Google Scholar] [PubMed]
- Huang, F.-C.; Chang, C.-C.; Lou, P.-J.; Kuo, I.-C.; Chien, C.-W.; Chen, C.-T.; Shieh, F.-Y.; Chang, T.-C.; Lin, J.-J. G-quadruplex stabilizer 3, 6-bis (1-methyl-4-vinylpyridinium) carba-zole diiodide induces accelerated senescence and inhibits tumorigenic properties in cancer cells. Mol. Cancer Res. 2008, 6, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Biffi, G.; Tannahill, D.; Miller, J.; Howat, W.J.; Balasubramanian, S. Elevated levels of g-quadruplex formation in human stomach and liver cancer tissues. PLoS ONE 2014, 9, e102711. [Google Scholar] [CrossRef]
- Biffi, G.; Tannahill, D.; McCafferty, J.; Balasubramanian, S. Quantitative visualization of DNA g-quadruplex structures in human cells. Nat. Chem. 2013, 5, 182–186. [Google Scholar] [CrossRef]
- Laguerre, A.; Wong, H.M.Y.; Monchaud, D. Direct visualization of both DNA and RNA quad-ruplexes in human cells via an uncommon spectroscopic method. Sci. Rep. 2016, 6, 32141. [Google Scholar] [CrossRef]
- Biffi, G.; Di Antonio, M.; Tannahill, D.; Balasubramanian, S. Visualization and selective chemical targeting of RNA g-quadruplex structures in the cytoplasm of human cells. Nat. Chem. 2014, 6, 75–80. [Google Scholar] [CrossRef]
- Chang, C.-C.; Wu, J.-Y.; Chang, T.-C. A carbazole derivative synthesis for stabilizing the quadruplex structure. J. Chin. Chem. Soc. 2003, 50, 185–188. [Google Scholar] [CrossRef]
- Wang, Y.; Patel, D.J. Solution structure of the human telomeric repeat d[ag3(t2ag3)3] g-tetraplex. Structure 1993, 1, 263–281. [Google Scholar] [CrossRef]
- Balagurumoorthy, P.; Brahmachari, S.K. Structure and stability of human telomeric se-quence. J. Biol. Chem. 1994, 269, 21858–21869. [Google Scholar] [PubMed]
- Kang, C.-C.; Huang, W.-C.; Kouh, C.-W.; Wang, Z.-F.; Cho, C.-C.; Chang, C.-C.; Wang, C.-L.; Chang, T.-C.; Seeman, J.; Huang, L.J. Chemical principles for the design of a novel fluores-cent probe with high cancer-targeting selectivity and sensitivity. Integr. Boil. 2013, 5, 1217–1228. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.-L.; Lin, L.; Lou, P.-J.; Chang, T.-C.; Young, T.-H. Detection of cell carcinogenic trans-formation by a quadruplex DNA binding fluorescent probe. PLoS ONE 2014, 9, e86143. [Google Scholar]
- Huang, W.-C.; Tseng, T.-Y.; Chen, Y.-T.; Chang, C.-C.; Wang, Z.-F.; Wang, C.-L.; Hsu, T.-N.; Li, P.-T.; Chen, C.-T.; Lin, J.-J. Direct evidence of mitochondrial g-quadruplex DNA by using flu-orescent anti-cancer agents. Nucleic Acids Res. 2015, 43, 10102–10113. [Google Scholar] [PubMed]
- Lu, Y.-J.; Hu, D.-P.; Zhang, K.; Wong, W.-L.; Chow, C.-F. New pyridinium-based fluorescent dyes: A comparison of symmetry and side-group effects on g-quadruplex DNA binding se-lectivity and application in live cell imaging. Biosens. Bioelectron. 2016, 81, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Nygren, J.; Svanvik, N.; Kubista, M. The interactions between the fluorescent dye thiazole orange and DNA. Biopolymers 1998, 46, 39–51. [Google Scholar] [CrossRef]
- Rye, H.S.; Quesada, M.A.; Peck, K.; Mathies, R.A.; Glazer, A.N. High-sensitivity two-color detection of double-stranded DNA with a confocal fluorescence gel scanner using ethidium homodimer and thiazole orange. Nucleic Acids Res. 1991, 19, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Rye, H.S.; Yue, S.; Wemmer, D.E.; Quesada, M.A.; Haugland, R.P.; Mathies, R.A.; Glazer, A.N. Stable fluorescent complexes of double-stranded DNA with bis-intercalating asym-metric cyanine dyes: Properties and applications. Nucleic Acids Res. 1992, 20, 2803–2812. [Google Scholar] [CrossRef] [PubMed]
- Lubitz, I.; Zikich, D.; Kotlyar, A. Specific high-affinity binding of thiazole orange to triplex and g-quadruplex DNA. Biochemistry 2010, 49, 3567–3574. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.-J.; Deng, Q.; Hou, J.-Q.; Hu, D.-P.; Wang, Z.-Y.; Zhang, K.; Luyt, L.G.; Wong, W.-L.; Chow, C.-F. Molecular engineering of thiazole orange dye: Change of fluorescent signaling from universal to specific upon binding with nucleic acids in bioassay. ACS Chem. Boil. 2016, 11, 1019–1029. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; De Cian, A.; Teulade-Fechou, M.P.; Mergny, J.L.; Monchaud, D. Engineering bisquinolinium/thiazole orange conjugates for fluorescent sensing of g-quadruplex DNA. Angew. Chem. Int. Ed. 2009, 48, 2188–2191. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.-J.; Yan, S.-C.; Chan, F.-Y.; Zou, L.; Chung, W.-H.; Wong, W.-L.; Qiu, B.; Sun, N.; Chan, P.-H.; Huang, Z.-S. Benzothiazole-substituted benzofuroquinolinium dye: A selective switch-on fluorescent probe for g-quadruplex. Chem. Commun. 2011, 47, 4971–4973. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.-J.; Deng, Q.; Hu, D.-P.; Wang, Z.-Y.; Huang, B.-H.; Du, Z.-Y.; Fang, Y.-X.; Wong, W.-L.; Zhang, K.; Chow, C.-F. A molecular fluorescent dye for specific staining and imaging of RNA in live cells: A novel ligand integration from classical thiazole orange and styryl compounds. Chem. Commun. 2015, 51, 15241–15244. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Sun, H.; Wang, L.; Liu, Y.; Chen, H.; Li, Q.; Guan, A.; Liu, M.; Tang, Y. Real-time monitoring of DNA g-quadruplexes in living cells with a small-molecule fluorescent probe. Nucleic Acids Res. 2018, 46, 7522–7532. [Google Scholar] [CrossRef] [PubMed]
- Nikan, M.; Sherman, J.C. Template assembled synthetic g-quartets (tasqs). Angew. Chem. Int. Ed. 2008, 47, 4900–4902. [Google Scholar] [CrossRef]
- Laguerre, A.; Desbois, N.; Stefan, L.; Richard, P.; Gros, C.P.; Monchaud, D. Porphyrin-based design of bioinspired multitarget quadruplex ligands. ChemMedChem 2014, 9, 2035–2039. [Google Scholar] [CrossRef]
- Laguerre, A.; Hukezalie, K.; Winckler, P.; Katranji, F.; Chanteloup, G.; Pirrotta, M.; Perrier-Cornet, J.-M.; Wong, J.M.Y.; Monchaud, D. Visualization of RNA-quadruplexes in live cells. J. Am. Chem. Soc. 2015, 137, 8521–8525. [Google Scholar] [CrossRef]
- Yang, C.; Hu, R.; Li, Q.; Li, S.; Xiang, J.; Guo, X.; Wang, S.; Zeng, Y.; Li, Y.; Yang, G. Visualiza-tion of parallel g-quadruplexes in cells with a series of new developed bis(4-aminobenzylidene)acetone derivatives. ACS Omega 2018, 3, 10487–10492. [Google Scholar] [CrossRef]
- Shivalingam, A.; Izquierdo, M.A.; Marois, A.L.; Vysniauskas, A.; Suhling, K.; Kuimova, M.K.; Vilar, R. The interactions between a small molecule and g-quadruplexes are visualized by fluorescence lifetime imaging microscopy. Nat. Commun. 2015, 6, 8178. [Google Scholar] [CrossRef] [PubMed]
- Dash, J.; Das, R.N.; Hegde, N.; Dan Pantos, G.; Shirude, P.S.; Rodriguez, R.; Shahid, R.; Bal-asubramanian, S. Synthesis of bis-indole carboxamides as g-quadruplex stabilizing and in-ducing ligands. Chem. Eur. J. 2012, 18, 554–564. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Zhang, X.; Wang, L.; Liu, Y.; Bing, T.; Liu, X.; Shangguan, D. Interaction of bisbenzim-idazole-substituted carbazole derivatives with g-quadruplexes and living cells. RSC Adv. 2015, 5, 75911–75917. [Google Scholar] [CrossRef]
- Cogoi, S.; Shchekotokhin, A.E.; Membrino, A.; Sinkevich, Y.B.; Xodo, L.E. Guanidino an-thrathiphenediones as g-quadruplex binders: Uptake, intracellular localization, and anti-harvey-ras gene activity in bladder cancer cells. J. Med. Chem. 2013, 56, 2764–2778. [Google Scholar] [CrossRef] [PubMed]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chilka, P.; Desai, N.; Datta, B. Small Molecule Fluorescent Probes for G- Quadruplex Visualization as Potential Cancer Theranostic Agents. Molecules 2019, 24, 752. https://doi.org/10.3390/molecules24040752
Chilka P, Desai N, Datta B. Small Molecule Fluorescent Probes for G- Quadruplex Visualization as Potential Cancer Theranostic Agents. Molecules. 2019; 24(4):752. https://doi.org/10.3390/molecules24040752
Chicago/Turabian StyleChilka, Pallavi, Nakshi Desai, and Bhaskar Datta. 2019. "Small Molecule Fluorescent Probes for G- Quadruplex Visualization as Potential Cancer Theranostic Agents" Molecules 24, no. 4: 752. https://doi.org/10.3390/molecules24040752
APA StyleChilka, P., Desai, N., & Datta, B. (2019). Small Molecule Fluorescent Probes for G- Quadruplex Visualization as Potential Cancer Theranostic Agents. Molecules, 24(4), 752. https://doi.org/10.3390/molecules24040752