Dietary Phytochemicals Targeting Cancer Stem Cells


 , , ,
, , ,  and
and 
Abstract
:1. Introduction
1.1. Aim of the Study
1.2. Source of Data
2. CSCs (Cancer Stem Cells)
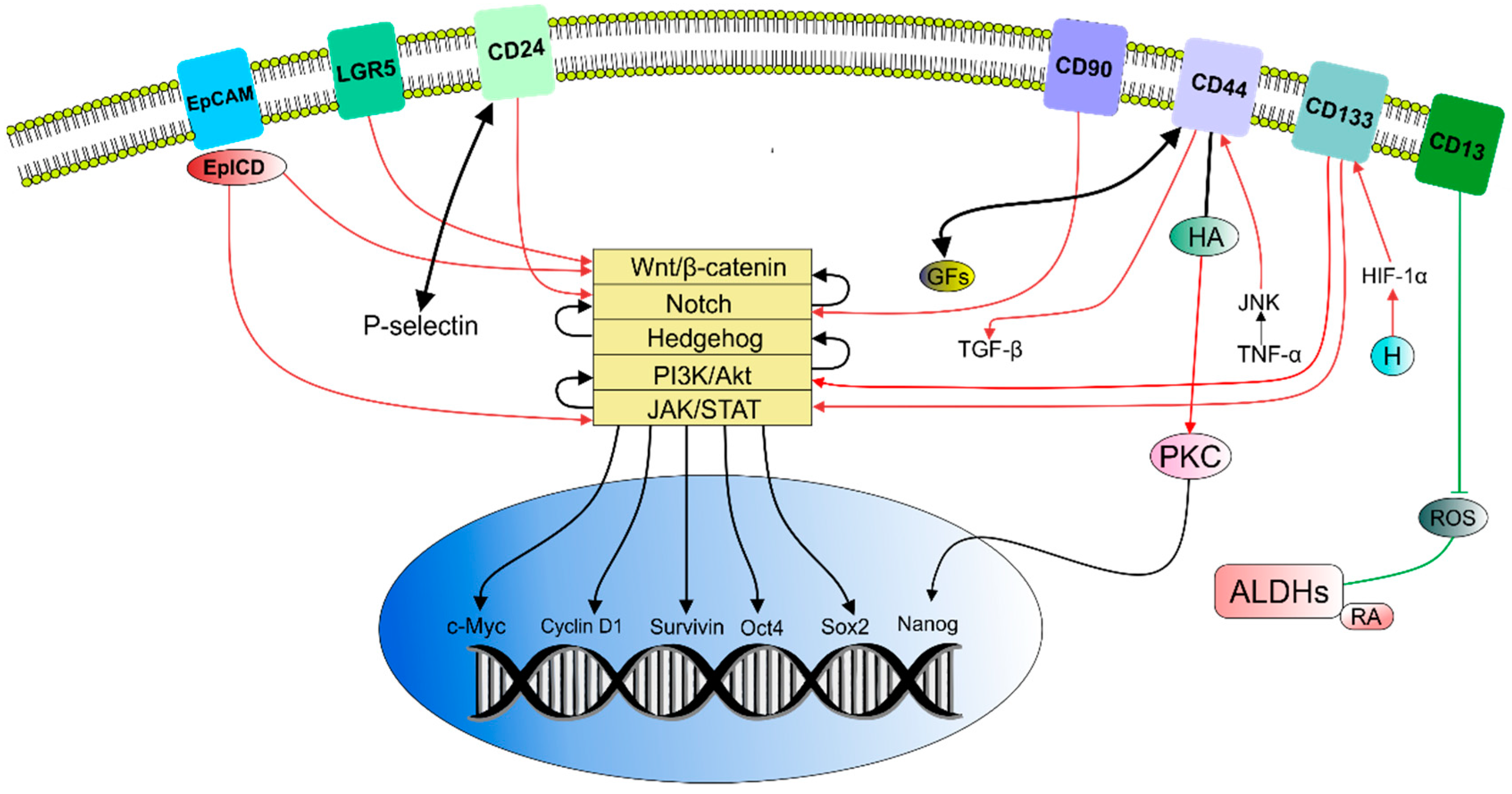
2.1. Cancer Stem Cells Markers
2.2. EMT (Epithelial–Mesenchymal Transition)
2.3. Molecular or Cellular Signaling of CSCs
2.3.1. Signaling Pathways in CSCs
2.3.2. Apoptotic and Death Resistance Signaling of CSCs
3. Natural Compounds Targeting CSCs in Cancer Research
3.1. Preclinical Research
3.1.1. Isolated Phytochemicals
3.1.2. Plant-Derived Functional Foods
3.2. Clinical research
4. Conclusion and Future Perspectives
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Konrad, C.V.; Murali, R.; Varghese, B.A.; Nair, R. The role of cancer stem cells in tumor heterogeneity and resistance to therapy. Can. J. Physiol. Pharmacol. 2017, 95, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Surh, Y.J. Chemopreventive and Therapeutic Potential of Phytochemicals Targeting Cancer Stem Cells. Curr. Pharmacol. Rep. 2015, 1, 302–311. [Google Scholar] [CrossRef]
- Kozovska, Z.; Gabrisova, V.; Kucerova, L. Colon cancer: Cancer stem cells markers, drug resistance and treatment. Biomed. Pharmacother. 2014, 68, 911–916. [Google Scholar] [CrossRef] [PubMed]
- Oo, A.K.K.; Calle, A.S.; Nair, N.; Mahmud, H.; Vaidyanath, A.; Yamauchi, J.; Khayrani, A.C.; Du, J.; Alam, M.J.; Seno, A.; et al. Up-Regulation of PI 3-Kinases and the Activation of PI3K-Akt Signaling Pathway in Cancer Stem-Like Cells Through DNA Hypomethylation Mediated by the Cancer Microenvironment. Transl. Oncol. 2018, 11, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Alison, M.R.; Islam, S.; Wright, N.A. Stem cells in cancer: Instigators and propagators? J. Cell Sci. 2010, 123, 2357–2368. [Google Scholar] [CrossRef] [PubMed]
- Salem, M.L.; El-Badawy, A.S.; Li, Z. Immunobiology and signaling pathways of cancer stem cells: Implication for cancer therapy. Cytotechnology 2015, 67, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Varghese, E.; Samuel, S.M.; Varghese, S.; Cheema, S.; Mamtani, R.; Büsselberg, D. Triptolide Decreases Cell Proliferation and Induces Cell Death in Triple Negative MDA-MB-231 Breast Cancer Cells. Biomolecules 2018, 8, 163. [Google Scholar] [CrossRef] [PubMed]
- Varghese, E.; Samuel, S.M.; Abotaleb, M.; Cheema, S.; Mamtani, R.; Büsselberg, D. The “Yin and Yang” of Natural Compounds in Anticancer Therapy of Triple-Negative Breast Cancers. Cancers 2018, 10, 346. [Google Scholar] [CrossRef] [PubMed]
- Abotaleb, M.; Samuel, S.M.; Varghese, E.; Varghese, S.; Kubatka, P.; Liskova, A.; Büsselberg, D. Flavonoids in Cancer and Apoptosis. Cancers 2019, 11, E28. [Google Scholar] [CrossRef] [PubMed]
- Lee, CH.; Chen, C.Y. Natural product-based therapeutics for the treatment of cancer stem cells: A patent review (2010–2013). Expert Opin. Ther. Pat. 2015. [Google Scholar] [CrossRef] [PubMed]
- Patel, P.; Chen, E.I. Cancer stem cells, tumor dormancy, and metastasis. Front. Endocrinol. 2012, 3, 125. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Ma, S.; Cao, K.; Zhou, S.; Zhao, A.; Li, M.; Qian, F.; Zhu, C. Therapeutic approaches targeting cancer stem cells. J. Cancer Res. Ther. 2018, 14, 1469–1475. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Shigdar, S.; Gantier, M.P.; Hou, Y.; Wang, L.; Li, Y.; Shamaileh, H.A.; Yin, W.; Zhou, S.F.; Zhao, X.; et al. Cancer stem cell targeted therapy: Progress amid controversies. Oncotarget 2015, 6, 44191–44206. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.Y.; Hu, D.P.; Xie, L.; Li, Y. Majumdar S1, Nonn L2, Hu H3, Shioda T4, Prins GS5. Isolation and functional interrogation of adult human prostate epithelial stem cells at single cell resolution. Stem Cell Res. 2017, 23, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Moharil, R.B.; Dive, A.; Khandekar, S.; Bodhade, A. Cancer stem cells: An insight. J. Oral Maxillofac. Pathol. 2017, 21, 463. [Google Scholar] [CrossRef] [PubMed]
- Palermo, R.; Ghirga, F.; Piccioni, M.G.; Bernardi, F.; Zhdanovskaya, N.; Infante, P.; Mori, M. Natural products inspired modulators of cancer stem cells-specific signaling pathways Notch and Hedgehog. Curr. Pharm. Des. 2019. [Google Scholar] [CrossRef] [PubMed]
- Toledo-Guzmán, M.E.; Bigoni-Ordóñez, G.D.; Ibáñez Hernández, M.; Ortiz-Sánchez, E. Cancer stem cell impact on clinical oncology. World J. Stem Cells 2018, 10, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.; Hlatky, L.; Jeong, Y.S.; Kim, D. Therapeutic Effectiveness of Anticancer Phytochemicals on Cancer Stem Cells. Toxins 2016, 8, 199. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Huang, Y.H.; Chen, J.L. Understanding and targeting cancer stem cells: Therapeutic implications and challenges. Acta Pharmacol. Sin. 2013, 34, 732–740. [Google Scholar] [CrossRef] [PubMed]
- Taylor, W.F.; Jabbarzadeh, E. The use of natural products to target cancer stem cells. Am. J. Cancer Res. 2017, 7, 1588–1605. [Google Scholar] [PubMed]
- Hu, W.Y.; Shi, G.B.; Lam, H.M.; Hu, D.P.; Ho, S.M.; Madueke, I.C.; Kajdacsy-Balla, A.; Prins, G.S. Estrogen-initiated transformation of prostate epithelium derived from normal human prostate stem-progenitor cells. Endocrinology 2011, 152, 2150–2163. [Google Scholar] [CrossRef] [PubMed]
- Prins, G.S.; Hu, W.Y.; Xie, L.; Shi, G.B.; Hu, D.P.; Birch, L.; Bosland, M.C. Evaluation of Bisphenol A (BPA) Exposures on Prostate Stem Cell Homeostasis and Prostate Cancer Risk in the NCTR-Sprague-Dawley Rat: An NIEHS/FDA CLARITY-BPA Consortium Study. Environ. Health Perspect. 2018, 126, 117001. [Google Scholar] [CrossRef] [PubMed]
- Morath, I.; Hartmann, T.N.; Orian-Rousseau, V. CD44: More than a mere stem cell marker. Int. J. Biochem. Cell Biol. 2016, 81, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.; Cheng, C.; Shao, Q.; Lin, Z.; Lu, S.; Chen, Y. CD24 promotes HCC progression via triggering Notch-related EMT and modulation of tumor microenvironment. Tumour Biol. 2016, 37, 6073–6084. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Bhanja, A.; Bhattacharyya, J.; Jaganathan, B.G. Multiple roles of CD90 in cancer. Tumour Biol. 2016, 37, 11611–11622. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.M.; Haraguchi, N.; Ishii, H.; Ohkuma, M.; Okano, M.; Mimori, K.; Eguchi, H.; Yamamoto, H.; Nagano, H.; Sekimoto, M.; et al. Increased CD13 expression reduces reactive oxygen species, promoting survival of liver cancer stem cells via an epithelial-mesenchymal transition-like phenomenon. Ann. Surg. Oncol. 2012, S539–S548. [Google Scholar] [CrossRef] [PubMed]
- Nakahata, K.; Uehara, S.; Nishikawa, S.; Kawatsu, M.; Zenitani, M.; Oue, T.; Okuyama, H. Aldehyde Dehydrogenase 1 (ALDH1) Is a Potential Marker for Cancer Stem Cells in Embryonal Rhabdomyosarcoma. PLoS ONE 2015, 10, e0125454. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Wang, S.; Li, M.Y.; Hu, B.G.; Liu, L.P.; Yang, S.L.; Yang, S.; Gong, Z.; Lai, P.B.S.; Chen, G.G. Cancer stem cells in hepatocellular carcinoma: An overview and promising therapeutic strategies. Ther. Adv. Med. Oncol. 2018, 10, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Zuo, X.; Wei, D. Concise Review: Emerging Role of CD44 in Cancer Stem Cells: A Promising Biomarker and Therapeutic Target. Stem Cells Transl. Med. 2015, 4, 1033–1043. [Google Scholar] [CrossRef] [PubMed]
- Jaggupilli, A.; Elkord, E. Significance of CD44 and CD24 as cancer stem cell markers: An enduring ambiguity. Clin. Dev. Immunol. 2012, 708036. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Ma, H.; Zhang, J.; Zhu, L.; Wang, C.; Yang, Y. Unraveling the roles of CD44/CD24 and ALDH1 as cancer stem cell markers in tumorigenesis and metastasis. Sci. Rep. 2017, 7, 13856. [Google Scholar] [CrossRef] [PubMed]
- Munz, M.; Baeuerle, P.A.; Gires, O. The emerging role of EpCAM in cancer and stem cell signaling. Cancer Res. 2009, 69, 5627–5629. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.W.; Song, Y.; Kim, S.H.; Kim, J.; Seo, H.R. Potential mechanisms of CD133 in cancer stem cells. Life Sci. 2017, 184, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Glumac, P.M.; LeBeau, A.M. The role of CD133 in cancer: A concise review. Clin. Transl. Med. 2018, 7, 18. [Google Scholar] [CrossRef] [PubMed]
- Haraguchi, N.; Ishii, H.; Mimori, K.; Tanaka, F.; Ohkuma, M.; Kim, H.M.; Akita, H.; Takiuchi, D.; Hatano, H.; Nagano, H.; et al. CD13 is a therapeutic target in human liver cancer stem cells. J. Clin. Investig. 2010, 120, 3326–3339. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Wang, P.; Wang, R.; Wang, J.; Liu, M.; Xiong, S.; Li, Y.; Cheng, B. The Notch pathway promotes the cancer stem cell characteristics of CD90+ cells in hepatocellular carcinoma. Oncotarget 2016, 7, 9525–9537. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, M.V.; Kala, M.; Nivsarkar, M. CD90 a potential cancer stem cell marker and a therapeutic target. Cancer Biomark. 2016, 16, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Tomita, H.; Tanaka, K.; Tanaka, T.; Hara, A. Aldehyde dehydrogenase 1A1 in stem cells and cancer. Oncotarget 2016, 7, 11018–11032. [Google Scholar] [CrossRef] [PubMed]
- Katoh, M. Canonical and non-canonical WNT signaling in cancer stem cells and their niches: Cellular heterogeneity, omics reprogramming, targeted therapy and tumor plasticity. Int. J. Oncol. 2017, 51, 1357–1369. [Google Scholar] [CrossRef] [PubMed]
- Matsui, W.H. Cancer stem cell signaling pathways. Medicine 2016, 95, S8–S19. [Google Scholar] [CrossRef] [PubMed]
- Koury, J.; Zhong, L.; Hao, J. Targeting Signaling Pathways in Cancer Stem Cells for Cancer Treatment. Stem Cells Int. 2017, 2925869. [Google Scholar] [CrossRef] [PubMed]
- Kuşoğlu, A.; Avcı, Ç.B. Cancer stem cells: A brief review of the current status. Gene 2019, 681, 80–85. [Google Scholar] [CrossRef]
- Lee, C.M.; Su, Y.H.; Huynh, T.T.; Lee, W.H.; Chiou, J.F.; Lin, Y.K.; Hsiao, M.; Wu, C.H.; Lin, Y.F.; Wu, A.T.; et al. BlueBerry Isolate, Pterostilbene, Functions as a Potential Anticancer Stem Cell Agent in Suppressing Irradiation-Mediated Enrichment of Hepatoma Stem Cells. Evid. Based Complement. Alternat. Med. 2013, 258425. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Settleman, J. EMT, cancer stem cells and drug resistance: An emerging axis of evil in the war on cancer. Oncogene 2010, 29, 4741–4751. [Google Scholar] [CrossRef] [PubMed]
- Dandawate, P.; Padhye, S.; Ahmad, A.; Sarkar, F.H. Novel strategies targeting cancer stem cells through phytochemicals and their analogs. Drug Deliv. Transl. Res. 2013, 3, 165–182. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, V.; Nataraj, R.; Thangaraj, G.S.; Karthikeyan, M.; Gnanasekaran, A.; Kaginelli, S.B.; Kuppanna, G.; Kallappa, C.G.; Basalingappa, K.M. Targeting Notch signalling pathway of cancer stem cells. Stem Cell Investig. 2018, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Samadani, A.A.; Norollahi, S.E.; Rashidy-Pour, A.; Mansour-Ghanaei, F.; Nemati, S.; Joukar, F.; Afshar, A.M.; Ghazanfari, S.; Safizadeh, M.; Rostami, P.; et al. Cancer signaling pathways with a therapeutic approach: An overview in epigenetic regulations of cancer stem cells. Biomed. Pharmacother. 2018, 108, 590–599. [Google Scholar] [CrossRef] [PubMed]
- Dandawate, P.R.; Subramaniam, D.; Jensen, R.A.; Anant, S. Targeting cancer stem cells and signaling pathways by phytochemicals: Novel approach for breast cancer therapy. Semin. Cancer Biol. 2016, 40, 192–208. [Google Scholar] [CrossRef] [PubMed]
- Du, W.Z.; Feng, Y.; Wang, X.F.; Piao, X.Y.; Cui, Y.Q.; Chen, L.C.; Lei, X.H.; Sun, X.; Liu, X.; Wang, H.B.; et al. Curcumin suppresses malignant glioma cells growth and induces apoptosis by inhibition of SHH/GLI1 signaling pathway in vitro and vivo. CNS Neurosci. Ther. 2013, 19, 926–936. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Graham, P.H.; Ni, J.; Hao, J.; Bucci, J.; Cozzi, P.J.; Li, Y. Targeting PI3K/Akt/mTOR signaling pathway in the treatment of prostate cancer radioresistance. Crit. Rev. Oncol. Hematol. 2015, 96, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Suvarna, V.; Murahari, M.; Khan, T.; Chaubey, P.; Sangave, P. Phytochemicals and PI3K Inhibitors in Cancer-An Insight. Front. Pharmacol. 2017, 8, 916. [Google Scholar] [CrossRef] [PubMed]
- Ning, Y.; Feng, W.; Cao, X.; Ren, K.; Quan, M.; Chen, A.; Xu, C.; Qiu, Y.; Cao, J.; Li, X.; et al. Genistein inhibits stemness of SKOV3 cells induced by macrophages co-cultured with ovarian cancer stem-like cells through IL-8/STAT3 axis. J. Exp. Clin. Cancer Res. 2019, 38, 19. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, K.; Resat, H. Constitutive activation of STAT3 in breast cancer cells: A review. Int. J. Cancer 2016, 138, 2570–2578. [Google Scholar] [CrossRef] [PubMed]
- Cafferkey, C.; Chau, I. Novel STAT 3 inhibitors for treating gastric cancer. Expert Opin. Investig. Drugs 2016, 25, 1023–1031. [Google Scholar] [CrossRef] [PubMed]
- He, Y.C.; Zhou, F.L.; Shen, Y.; Liao, D.F.; Cao, D. Apoptotic death of cancer stem cells for cancer therapy. Int. J. Mol. Sci. 2014, 15, 8335–8351. [Google Scholar] [CrossRef] [PubMed]
- Safa, A.R. Resistance to Cell Death and Its Modulation in Cancer Stem Cells. Crit. Rev. Oncog. 2016, 21, 203–219. [Google Scholar] [CrossRef] [PubMed]
- Rinkenbaugh, A.L.; Baldwin, A.S. The NF-κB Pathway and Cancer Stem Cells. Cells 2016, 5, 16. [Google Scholar] [CrossRef] [PubMed]
- Pramanik, K.C.; Makena, M.R.; Bhowmick, K.; Pandey, M.K. Advancement of NF-κB Signaling Pathway: A Novel Target in Pancreatic Cancer. Int. J. Mol. Sci. 2018, 19, 3890. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.M.; Chen, R.2.; Fong, D. Targeting cancer stem cells with dietary phytochemical—Repositioned drug combinations. Cancer Lett. 2018, 433, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Mayne, S.T.; Playdon, M.C.; Rock, C.L. Diet, nutrition, and cancer: Past, present and future. Nat. Rev. Clin. Oncol. 2016, 13, 504–515. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Saha, S.; Manna, A.; Mazumdar, M.; Chakraborty, S.; Paul, S.; Das, T. Targeting Cancer Stem Cells by Phytochemicals: A Multimodal Approach to Colorectal Cancer. Curr. Colorectal. Cancer Rep. 2014, 10, 431–441. [Google Scholar] [CrossRef]
- Jaiswal, A.S.; Marlow, B.P.; Gupta, N.; Narayan, S. Beta-catenin-mediated transactivation and cell-cell adhesion pathways are important in curcumin (diferuylmethane)-induced growth arrest and apoptosis in colon cancer cells. Oncogene 2002, 21, 8414–8427. [Google Scholar] [CrossRef] [PubMed]
- Ryu, M.J.; Cho, M.; Song, J.Y.; Yun, Y.S.; Choi, I.W.; Kim, D.E.; Park, B.S.; Oh, S. Natural derivatives of curcumin attenuate the Wnt/beta-catenin pathway through down-regulation of the transcriptional coactivator p300. Biochem. Biophys. Res. Commun. 2008, 377, 1304–1308. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, Y.; Banerjee, S.; Li, Y.; Sarkar, F.H. Notch-1 down-regulation by curcumin is associated with the inhibition of cell growth and the induction of apoptosis in pancreatic cancer cells. Cancer 2006, 106, 2503–2513. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, H.; Sueoka, E.; Rawangkan, A.; Suganuma, M. Human cancer stem cells are a target for cancer prevention using (-)-epigallocatechin gallate. J. Cancer Res. Clin. Oncol. 2017, 143, 2401–2412. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Jiang, Y.; Yang, X.; Wang, S.; Xie, CH.; Li, X.; Li, Y.; Chen, Y.; Wang, X.; Meng, Y.; et al. Wnt/β-catenin pathway mediates (−)-Epigallocatechin-3-gallate (EGCG) inhibition of lung cancer stem cells. Biochem. Biophys. Res. Commun. 2017, 482, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Nam, H.J.; Kang, H.J.; Kwon, H.W.; Lim, Y.C. Epigallocatechin-3-gallate attenuates head and neck cancer stem cell traits through suppression of Notch pathway. Eur. J. Cancer 2013, 49, 3210–3218. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wen, X.; Li, M.; Li, S.; Zhao, H. Targeting cancer stem cells and signaling pathways by resveratrol and pterostilbene. Biofactors 2018, 44, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Chang, H.; Peng, X.; Bai, Q.; Yi, L.; Zhou, Y.; Zhu, J.; Mi, M. Resveratrol inhibits breast cancer stem-like cells and induces autophagy via suppressing Wnt/β-catenin signaling pathway. PLoS ONE 2014, 9, e102535. [Google Scholar] [CrossRef] [PubMed]
- Cilibrasi, C.; Riva, G.; Romano, G.; Cadamuro, M.; Bazzoni, R.; Butta, V.; Paoletta, L.; Dalprà, L.; Strazzabosco, M.; Lavitrano, M.; et al. Resveratrol Impairs Glioma Stem Cells Proliferation and Motility by Modulating the Wnt Signaling Pathway. PLoS ONE 2017, 12, e0169854. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Jiang, D. Resveratrol eliminates cancer stem cells of osteosarcoma by STAT3 pathway inhibition. PLoS ONE 2018, 13, e0205918. [Google Scholar] [CrossRef] [PubMed]
- Mak, K.K.; Wu, A.T.; Lee, W.H.; Chang, T.C.; Chiou, J.F.; Wang, L.S.; Wu, CH.; Huang, C.Y.; Shieh, Y.S.; Chao, T.Y.; et al. Pterostilbene, a bioactive component of blueberries, suppresses the generation of breast cancer stem cells within tumor microenvironment and metastasis via modulating NF-κB/microRNA 448 circuit. Mol. Nutr. Food Res. 2013, 57, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.; Farooqui, M.; Rao, C.V. Role of Dietary Cancer-Preventive Phytochemicals in Pancreatic Cancer Stem Cells. Curr. Pharmacol. Rep. 2018, 4, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Fan, P.; Fan, S.; Wang, H.; Mao, J.; Shi, Y.; Ibrahim, M.M.; Ma, W.; Yu, X.; Hou, Z.; Wang, B.; et al. Genistein decreases the breast cancer stem-like cell population through Hedgehog pathway. Stem Cell Res. Ther. 2013, 4, 146. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Ren, K.; Song, Z.; Li, D.; Quan, M.; Zheng, Y.; Cao, J.; Zeng, W.; Zou, H. Difluoromethoxyl-5,4’-di-n-octyl genistein inhibits the stem-like characteristics of gastric cancer stem-like cells and reverses the phenotype of epithelial-mesenchymal transition in gastric cancer cells. Oncol. Rep. 2016, 36, 1157–1165. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, T. Targeting cancer stem cells by curcumin and clinical applications. Cancer Lett. 2014, 346, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, Y.; Wang, X.Q.; Meng, Y.; Zhang, Q.; Zhu, J.Y.; Chen, J.Q.; Cao, W.S.; Wang, X.Q.; Xie, C.F.; et al. Phenethyl isothiocyanate inhibits colorectal cancer stem cells by suppressing Wnt/β-catenin pathway. Phytother Res. 2018, 32, 2447–2455. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Domina, A.; Lim, G.; Chang, T.; Zhang, T. Evaluation of curcumin, a natural product in turmeric, on Burkitt lymphoma and acute myeloid leukemia cancer stem cell markers. Future Oncol. 2018, 14, 2353–2360. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Ye, M.; Lu, Y.; Zhang, H.; Chen, Q.; Huang, S.; Su, S. Curcumin Improves the Tumoricidal Effect of Mitomycin C by Suppressing ABCG2 Expression in Stem Cell-Like Breast Cancer Cells. PLoS ONE 2015, 10, e0136694. [Google Scholar] [CrossRef] [PubMed]
- Sha, J.; Li, J.; Wang, W.; Pan, L.; Cheng, J.; Li, L.; Zhao, H.; Lin, W. Curcumin induces G0/G1 arrest and apoptosis in hormone independent prostate cancer DU-145 cells by down regulating Notch signaling. Biomed. Pharmacother. 2016, 84, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Pistollato, F.; Giampieri, F.; Battino, M. The use of plant-derived bioactive compounds to target cancer stem cells and modulate tumor microenvironment. Food Chem. Toxicol. 2015, 75, 58–70. [Google Scholar] [CrossRef] [PubMed]
- Cheung, K.L.; Kong, A.N. Molecular targets of dietary phenethyl isothiocyanate and sulforaphane for cancer chemoprevention. AAPS J. 2010, 12, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Li, S.H.; Fu, J.; Watkins, D.N.; Srivastava, R.K.; Shankar, S. Sulforaphane regulates self-renewal of pancreatic cancer stem cells through the modulation of Sonic hedgehog-GLI pathway. Mol. Cell Biochem. 2013, 373, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.Q.; Xie, Y.K.; Wu, Y.; Li, L.L.; Liu, Y.; Miao, X.B.; Liu, Q.Z.; Yao, K.T.; Xiao, G.H. Sulforaphane inhibits cancer stem-like cell properties and cisplatin resistance through miR-214-mediated downregulation of c-MYC in non-small cell lung cancer. Oncotarget 2017, 8, 12067–12080. [Google Scholar] [CrossRef] [PubMed]
- Castro, N.P.; Rangel, M.C.; Merchant, A.S.; MacKinnon, G.M.; Cuttitta, F.; Salomon, D.S.; Kim, Y.S. Sulforaphane suppresses the growth of triple-negative breast cancer stem-like cells in vitro and in vivo. Cancer Prev. Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.H.; Kim, K.A.; Yoo, G.; Kim, S.Y.; Shin, J.M.; Kim, J.H.; Jung, S.H.; Kim, J.; Nho, C.W. Phenethyl isothiocyanate suppresses cancer stem cell properties in vitro and in a xenograft model. Phytomedicine 2017, 30, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Li, X.T.; Chen, Y.; Chen, J.Q.; Zhu, J.Y.; Meng, Y.; Wang, X.Q.; Li, Y.; Geng, S.S.; Xie, C.F.; et al. Wnt/β-catenin signaling mediates the suppressive effects of diallyl trisulfide on colorectal cancer stem cells. Cancer Chemother. Pharmacol. 2018, 81, 969–977. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Meng, Y.; Xie, C.; Zhu, J.; Wang, X.; Li, Y.; Geng, S.; Wu, J.; Zhong, C.; Li, M. Diallyl Trisulfide inhibits breast cancer stem cells via suppression of Wnt/β-catenin pathway. J. Cell Biochem. 2018, 119, 4134–4141. [Google Scholar] [CrossRef] [PubMed]
- Kapinova, A.; Stefanicka, P.; Kubatka, P.; Zubor, P.; Uramova, S.; Kello, M.; Mojzis, J.; Blahutova, D.; Qaradakhi, T.; Zulli, A.; et al. Are plant-based functional foods better choice against cancer than single phytochemicals? A critical review of current breast cancer research. Biomed. Pharmacother. 2017, 96, 1465–1477. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.M.; Kim, I.H.; Nam, T.J. Capsosiphon fulvescens glycoprotein inhibits AGS gastric cancer cell proliferation by downregulating Wnt-1 signaling. Int. J. Oncol. 2013, 43, 1395–1401. [Google Scholar] [CrossRef] [PubMed]
- Kubatka, P.; Kello, M.; Kajo, K.; Kruzliak, P.; Výbohová, D.; Mojžiš, J.; Adamkov, M.; Fialová, S.; Veizerová, L.; Zulli, A.; et al. Oregano demonstrates distinct tumour-suppressive effects in the breast carcinoma model. Eur. J. Nutr. 2017, 56, 1303–1316. [Google Scholar] [CrossRef] [PubMed]
- Kubatka, P.; Uramova, S.; Kello, M.; Kajo, K.; Kruzliak, P.; Mojzis, J.; Vybohova, D.; Adamkov, M.; Jasek, K.; Lasabova, Z.; et al. Antineoplastic effects of clove buds (Syzygium aromaticum L.) in the model of breast carcinoma. J. Cell Mol. Med. 2017, 21, 2837–2851. [Google Scholar] [CrossRef] [PubMed]
- Mandal, A.; Bhatia, D.; Bishayee, A. Anti-Inflammatory Mechanism Involved in Pomegranate-Mediated Prevention of Breast Cancer: The Role of NF-κB and Nrf2 Signaling Pathways. Nutrients 2017, 9, E436. [Google Scholar] [CrossRef] [PubMed]
- Mandal, A.; Bishayee, A. Trianthema portulacastrum Linn. displays anti-inflammatory responses during chemically induced rat mammary tumorigenesis through simultaneous and differential regulation of NF-κB and Nrf2 signaling pathways. Int J. Mol. Sci. 2015, 16, 2426–2445. [Google Scholar] [CrossRef] [PubMed]
- Dong, R.; Chen, P.; Chen, Q. Extract of the Medicinal Plant Pao Pereira Inhibits Pancreatic Cancer Stem-Like Cell In Vitro and In Vivo. Integr. Cancer Ther. 2018, 17, 1204–1215. [Google Scholar] [CrossRef] [PubMed]
- Dong, R.; Chen, P.; Chen, Q. Inhibition of pancreatic cancer stem cells by Rauwolfia vomitoria extract. Oncol. Rep. 2018, 40, 3144–3154. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, S.; Wei, C.; Rankin, G.O.; Ye, X.; Chen, Y.C. Dietary compound proanthocyanidins from Chinese bayberry (Myrica rubra Sieb. et Zucc.) leaves attenuate chemotherapy-resistant ovarian cancer stem cell traits via targeting the Wnt/β-catenin signaling pathway and inducing G1 cell cycle arrest. Food Funct. 2018, 9, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Aravindan, S.; Ramraj, S.K.; Somasundaram, S.T.; Herman, T.S.; Aravindan, N. Polyphenols from marine brown algae target radiotherapy-coordinated EMT and stemness-maintenance in residual pancreatic cancer. Stem Cell Res. Ther. 2015, 6, 182. [Google Scholar] [CrossRef] [PubMed]
- Yen, CH.; Lai, C.C.; Shia, T.H.; Chen, M.; Yu, H.C.; Liu, Y.P.; Chang, F.R. Gynura divaricata attenuates tumor growth and tumor relapse after cisplatin therapy in HCC xenograft model through suppression of cancer stem cell growth and Wnt/β-catenin signalling. J. Ethnopharmacol. 2018, 213, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Reddivari, L.; Charepalli, V.; Radhakrishnan, S.; Vadde, R.; Elias, R.J.; Lambert, J.D.; Vanamala, J.K. Grape compounds suppress colon cancer stem cells in vitro and in a rodent model of colon carcinogenesis. BMC Complement. Altern. Med. 2016, 16, 278. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, A.V.; Martinez, M.; Stamos, M.J.; Moyer, M.P.; Planutis, K.; Hope, C.; Holcombe, R.F. Results of a phase I pilot clinical trial examining the effect of plant-derived resveratrol and grape powder on Wnt pathway target gene expression in colonic mucosa and colon cancer. Cancer Manag. Res. 2009, 1, 25–37, PMC3004662. [Google Scholar] [PubMed]
- Messing, E.; Gee, J.R.; Saltzstein, D.R.; Kim, K.; diSant‘Agnese, A.; Kolesar, J.; Harris, L.; Faerber, A.; Havighurst, T.; Young, J.M.; et al. A phase 2 cancer chemoprevention biomarker trial of isoflavone G-2535 (genistein) in presurgical bladder cancer patients. Cancer Prev. Res. 2012, 5, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Nuñez-Sánchez, M.A.; González-Sarrías, A.; García-Villalba, R.; Monedero-Saiz, T.; García-Talavera, N.V.; Gómez-Sánchez, M.B.; Sánchez-Álvarez, C.; García-Albert, A.M.; Rodríguez-Gil, F.J.; Ruiz-Marín, M.; et al. Gene expression changes in colon tissues from colorectal cancer patients following the intake of an ellagitannin-containing pomegranate extract: A randomized clinical trial. J. Nutr. Biochem. 2017, 42, 126–133. [Google Scholar] [CrossRef] [PubMed]
- ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ (accessed on 28 January 2019).




{kind=link}
{kind=link}
{kind=link}
| Cancer Type | Marker | References |
|---|---|---|
| Brain cancer | CD34+/CD38−/CD133+/CD44+ | [6,23] |
| Breast cancer | CD44+/CD24−/Lineage-/ALDH1+/ EpCAM+ | [18,20,23] |
| Colon cancer | CD133+/CD44+/CD166+/ ALDH1+/LGR5+/EpCAM+ | [6,18,19] |
| Leukemia | CD34+/CD38−/CD90− | [6,18,19] |
| Liver | CD133+/CD90+/EpCAM+ | [6] |
| Lung | CD133+/CD44+/CD90+ | [2,18] |
| Ovary | CD44+/ALDH1+/CD133+ | [6,18] |
| Pancreas | CD44+/CD24+/CD133+/EpCAM+ | [18,23] |
| Signaling Pathway | Cancer Type | Mechanism of Action | References |
|---|---|---|---|
| Notch | Brain cancer | ↑Notch1 ↑JAG1 ↑DLL1 | [6,40,44] |
| T cell acute lymphoblastic leukemia | ↑ Notch1 | ||
| Breast cancer | ↑Notch1 ↑JAG1, | ||
| Pancreatic cancer | ↑Notch1 ↑Notch3 ↑Jag1 ↑Jag2 ↑Hes1 | ||
| Non-Small Lung Cancer | ↑ Notch3 | ||
| Wnt/β-catenin | Breast cancer | ↑LEF-1 ↑TCF-4 ↑cyclin D1 ↑β-catenin ↓SFRP | [6,40,41,47] |
| Colorectal carcinoma Brain tumor Prostate cancer | Mutations in APC/β-catenin site | ||
| Hematologic cancer Skin cancer Lung cancer | ↓ WIF-1 ↓SFRP-1 ↓ DKK ↓AXIN2 | ||
| Hedgehog | Colon cancer | ↑sHH ↑GLI2 | [6,40] |
| Medulloblastoma predisposition | Mutations in PTCH1 | ||
| Myeloma | ↑SMO ↑GLI1 | ||
| Glioma | ↑GLI1 ↑SHH ↑PTCH1 | ||
| PI3K/Akt/mTOR | Gastric cancer | ↑Akt1 | [19] |
| Ovarian cancer Pancreatic cancer | ↑Akt2 | ||
| T cell acute lymphoblastic leukemia Melanoma Endometrial carcinoma Prostate cancer Glioblastoma | Mutations in PTEN | ||
| JAK/STAT | Breast cancer Gastric cancer Glioblastoma | ↑STAT3 | [53,54,40] |
| Phytochemical (Isolated or Mixture) | Cell Line/Animal Model | Mechanism | References |
|---|---|---|---|
| EGCG | A549, H1299 | ↓β-catenin ↓CD133 ↓CD44 ↓ALDH1A1 ↓Nanog ↓Oct4 | [66] |
| EGCG/ EGCG and cisplatin | HNSC CSCs BALB/c nude mice | ↓Oct4 ↓Sox2 ↓Nanog ↓CD44 ↓ABCC2 ↓ABCG2 | [67] |
| Resveratrol | MCF-7, SUM159 NOD/SCID xenografted mice | →autophagy ↓Wnt/β-catenin | [69] |
| GBM2, GBM7, G144, G179, G166, GliNS2, GBM04 | ↓β-catenin ↓c-Myc ↓Twist1 ↓Snail1 | [70] | |
| MNNG/HOS. MG-63, hFOB1.19 | ↓JAK2/STAT3 ↓CD133 | [71] | |
| Pterostilbene | MCF7, MDA-MB-231 | ↓NF-κB ↓Twist1 ↓vimentin ↑E-cadherin | [72] |
| HCC Mahlavu | ↓c-Myc ↓COX-2 ↓vimentin ↓CXCR4 ↓Twist1 | [43] | |
| Genistein | MCF-7 Nude mice | ↓SMO ↓GLI1 | [74] |
| SKOV3 Nude mice | ↓CD 163 ↓p-STAT3 ↓IL-10 ↑IL-12 ↓CD133 ↓CD44 | [52] | |
| GCSLCs | ↓ Twist1 ↓N-cadherin ↑E-cadherin ↓CD133 ↓CD44 ↓ALDH1 | [75] | |
| Curcumin | BL41-3, Ramos, DG-75, THP-1 | ↓ALDH+ cells ↓GLI1 ↓Notch1 ↓cyclin D1 | [78] |
| MCF-7, MDA-MB-231 Athymic mice | ↓ABCG2 ↓ABCC1 | [79] | |
| U87, T98G U87-implanted nude mice | ↓sHH ↓SMO ↓GLI1 ↓cyclin D1 ↓Bcl-2 ↓FoxM1 ↑Bax/Bcl-2 ratio | [49] | |
| DU-145 | ↓cyclin D1 ↓CDK2 ↓Bcl-2 ↑p21 ↑p27 ↑p53 | [80] | |
| Sulforaphane | NOD/SCID/IL2Rgamma mice | ↓SMO ↓GLI1 ↓GLI2 ↓Nanog ↓Oct-4 ↑Bcl-2 ↓Zeb-1 ↓E-cadherin ↓VEGF ↓PDGFRα | [83] |
| BalbC/nude mice | ↓CR1 ↓CR3 ↓Nanog ↓ALDHH1A1 ↓Wnt3 ↓Notch4 | [85] | |
| BEAS-2B, H460, H1299, A549 | ↓c-Myc | [84] | |
| Phenethyl isothiocyanate | DLD-1 SW480 | ↓size/number of cell spheroids ↓CD133+ | [77] |
| 293T, NCCIT, HCT116 Xenograft model | ↓Oct4 ↓Sox-2 ↓Nanog | [86] | |
| Diallyl trisulfide | SW48, DLD-1 | ↓β-catenin ↓c-Myc ↓cyclin D1 | [87] |
| MCF-7, SUM159 | ↓CD44 ↓ALDH1A1 ↓Nanog ↓Oct4 | [88] | |
| Capsosiphon fulvescens | AGS | ↓Wnt-1 ↓β-catenin → G0/G1 arrest | [90] |
| Origanum vulgare | NMU-induced rat mammary carcinogenesis | ↓CD24 ↓EpCAM | [91] |
| Syzygium aromaticum | NMU-induced rat mammary carcinogenesis | ↓CD24 ↓CD44 ↑ALDH1 | [92] |
| Pomegranate extract | DMBA-induced rat mammary carcinogenesis | ↓ER-α:ER-β↓β-catenin ↓cyclin D1 ↓COX-2 ↑Nrf2 IκBα degradation and NF-κB translocation blockage | [93] |
| Trianthema portulacastrum extract | DMBA-induced rat mammary carcinogenesis | ↓COX-2 ↑Nrf2 IκBα degradation and NF-κB translocation blockage | [94] |
| Pao Pereira extract | PANC-1, MIA PaCa-2, AsPC-1, HPAF-II, BxPC-3 in immunocompromised mice | ↓Nanog ↓β-catenin | [95] |
| Rauwolfia vomitoria extract | PANC-1, AsPC-1, HPAF-II, BxPC-3 and MiA PaCa-2 in immunocompromised mice | ↓Nanog ↓β-catenin | [96] |
| Chinese bayberry leaf proanthocyanidins (BLPs) | OVCAR-3 | ↓β-catenin ↓cyclin D1 ↓c-Myc →G1 arrest | [97] |
| HT-EA, SA-EA, PT-EA | Panc-1, MiaPaCa-2, Panc-3.27, and BxPC-3 Athymic NCr-nu/nu nude mice | ↓Nanog ↓Oct-4 ↓Sox2 ↓N-cadherin | [98] |
| Water extract of Gynura divaricata (GDE) | Huh7, Hep3B | ↓β-catenin | [99] |
| Resveratrol (RSV) and grape seed extracts (GSE) | Human colon CSCs Azoxymethane-induced mice | ↓ nuclear translocation of β-catenin ↓c-Myc ↓cyclin D1 ↑p53 ↑Bax/Bcl-2 | [100] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liskova, A.; Kubatka, P.; Samec, M.; Zubor, P.; Mlyncek, M.; Bielik, T.; Samuel, S.M.; Zulli, A.; Kwon, T.K.; Büsselberg, D. Dietary Phytochemicals Targeting Cancer Stem Cells. Molecules 2019, 24, 899. https://doi.org/10.3390/molecules24050899
Liskova A, Kubatka P, Samec M, Zubor P, Mlyncek M, Bielik T, Samuel SM, Zulli A, Kwon TK, Büsselberg D. Dietary Phytochemicals Targeting Cancer Stem Cells. Molecules. 2019; 24(5):899. https://doi.org/10.3390/molecules24050899
Chicago/Turabian StyleLiskova, Alena, Peter Kubatka, Marek Samec, Pavol Zubor, Milos Mlyncek, Tibor Bielik, Samson Mathews Samuel, Anthony Zulli, Taeg Kyu Kwon, and Dietrich Büsselberg. 2019. "Dietary Phytochemicals Targeting Cancer Stem Cells" Molecules 24, no. 5: 899. https://doi.org/10.3390/molecules24050899
APA StyleLiskova, A., Kubatka, P., Samec, M., Zubor, P., Mlyncek, M., Bielik, T., Samuel, S. M., Zulli, A., Kwon, T. K., & Büsselberg, D. (2019). Dietary Phytochemicals Targeting Cancer Stem Cells. Molecules, 24(5), 899. https://doi.org/10.3390/molecules24050899