1. Introduction
Panicum miliaceum L., also known as proso millet, belongs to the
Graminacea family and is an ancient crop that has been cultivated for more than 7000 years [
1]. It is also considered an economically important crop that is cultivated mainly in China, Eastern Europe, India, Russia, and Northern America [
2]. This plant is mainly used as functional food owing to its high protein content [
3,
4]. In addition, it is also used as a raw material for liquor and beer, animal fodder, and as a sizing agent in the textile industry in Africa and Asia [
5,
6]. It is a gluten-free food whose mild flavor is used in food industries of America and Europe [
7]. Moreover, millet requires relatively low amounts of water and nutrients for growth and can be cultivated in a wide range of altitudes [
6]. It is considered a rich source of micronutrients such as iron (Fe), zinc (Zn), copper (Cu), manganese (Mn), vitamins, and trace elements [
8]. Therefore, this plant is receiving increasing attention from food industries. Moreover, previous studies on this plant reported about its anticancer and antidiabetic properties and its ability to prevent coronary heart disease and liver diseases [
9,
10,
11].
The rapid loss of genetic diversity of plant species due to changing environment creates an urgent need to conserve existing important alleles and valuable phenotypic traits of genetic resources and to gather information about the genetic variation of different accessions. Studies on genetic diversity not only increase our knowledge but are also vital for improving yield, disease resistance, and nutritional value [
12], as they facilitate the genetic integration of new useful genes [
13], and are important for future breeding programs [
14]. Moreover, characterizing genetic variation of plant species helps select superior germplasm for the rational use of genetic resources [
15].
Characterizing morphological traits is very important when assessing the agronomic value and taxonomically classifying of plant species [
16]. Morphological traits are also useful tools in studying geographical patterns in large gene bank collections of plant species [
17,
18]. However, morphological characterization has its own disadvantages, as it is time-consuming and varies according to the environmental changes and low level of polymorphism, low heritability, late expression, limited discriminative power, and lower potential to measure relatedness and genetic similarity [
19]. DNA fingerprinting markers play a major role in revealing polymorphisms. The selection of accessions is more accurate when genetic markers are used rather than morphological traits, as genetic markers can be obtained from all tissues of organisms, allow early detection, and can produce highly specific pattern of bands for each individual. Genetic markers, also called DNA markers, show variation among individuals based on variation in DNA sequences [
20,
21,
22]. There are many kinds of molecular markers available, including amplified fragment length polymorphism (AFLP), restriction fragment length polymorphism (RFLP), random amplified polymorphic DNA (RAPD), simple sequence repeats (SSR), single nucleotide polymorphisms (SNP), inter-simple sequence repeats (ISSR), and diversity arrays technology (DArT) markers, among which ISSR markers are most reliable as they are simple, cost-effective, and highly polymorphic, and can augment plant breeding programs [
23].
ISSR markers have become reliable tools in the field of DNA fingerprinting and analyzing genetic diversity of plants. Since these markers do not require sequence information, variations may be found at several loci simultaneously, are microsatellite-sequence specific, are reliable for DNA profiling, especially for closely related species [
24], and are highly reproducible and polymorphic, ISSRs are used extensively in taxonomic studies [
25] to determine phylogenetic relationships and plant breeding programs [
26,
27,
28]. In contrast, the use of RAPD represents a very cost-effective, rapid, and efficient approach in detect higher levels of polymorphisms and unique alleles in plants [
20]. The usefulness of these molecular markers has been extensively demonstrated for detecting genetic variations in several plant species [
29,
30,
31,
32]. Applying two marker systems gives more reliable and correct information about genetic diversity. Moreover, the omissions recorded by certain markers can be solved or minimized by using more than one marker [
33].
Previous studies on the genetic diversity in
P. miliaceum were employed by using genetic markers [
34,
35,
36,
37]. Even though genetic diversity was assessed using different types of DNA markers, the genetic diversity of
P. miliaceum has been inadequately assessed. Variation in the chlorophyll content in the plant can be used as an indicator of plant vigor [
38]. These pigments are also associated with the antioxidant properties of plants [
38]. A number of studies have reported that phytochemicals, such as phenolic compounds, flavonoids, and isoflavones not only act as antioxidant compounds [
39], but are also safe for consumption as dietary supplements. Other studies have reported natural antioxidants extracted from plants that effectively inhibited or reduced the formation or scavenging of free radicals and carcinogens [
40].
P. miliaceum has a wide geographical range, which results in variations in nutritional value, biological activity, and agro-morphological traits. To our knowledge, there is no report on the chlorophyll content, morphological variation, and antioxidant and genetic diversity assessed using a combined molecular marker technique in
P. miliaceum accessions. Moreover, the antioxidant profiles of
P. miliaceum accessions that originate from different parts of the Korean peninsula have not been fully examined in earlier studies. Previous research on
P. miliaceum accessions were limited and incomplete in the absence of these parameters. Therefore, the objectives of the present study were to compare the bio-morphological and genetic traits in the accessions and to screen biological activities and their correlations with total phenolic content (TPC) and total flavonoid content (TFC). By screening the accessions, we can have diverse adapted germplasm for safe and productive
P. miliaceum production. Furthermore, the present comparative study might identify accessions with useful genes for breeding programs, and may be able to introduce a commercial variety to farmers.
4. Material and Methods
4.1. Chemicals, Standard Compounds and Solvents
All commercial standard compounds with at least 99% purity used for analyzing individual phenolic compounds were purchased from Sigma Aldrich Chemical Co. (St. Louis, MO, USA) and Extrasynthese (Genay Cedex, France). HPLC-grade methanol hexane, ethyl acetate, and butanol were supplied from Avantor–J. T. Baker (Phillipsburg, NJ, USA). Water used in this study was of analytical grade purified by a Milli-Q Water Purification System (Millipore, Bedford, MA, USA). Other chemical compounds such as DPPH and ABTS, which were used for assessing biological activity, were purchased from Avantor–J. T. Baker (Phillipsburg, NJ, USA).
4.2. Plant Materials
In this study, 15 accessions of
P. miliaceum collected from different eco-geographical regions of Korea were grown from 2014–2017 at the Agriculture Research Field of Kangwon National University, South Korea (
Table 10). All the accessions used in this study were grown naturally under ideal, similar plant growth conditions using the same field management practices at 99 m elevation.
4.3. Evaluation of Morphological Traits in P. miliaceum Accessions
Prior to cultivating the accessions, experimental units were prepared in a completely randomized block design, each unit consisting of 10 replicates. The length of each row was maintained at 70 m with 1 m between adjacent rows with approximately 80-cm gaps between seedlings. The rainfall recorded during the cultivated period was 200 nm with the average minimum and maximum temperature of 20 °C and 35 °C, respectively. The soil of the experimental field was maintained at pH of 6.1 and irrigated regularly (once a week) using a drip-irrigation system. To increase the nutrient contents of soil, the recommended dose of fertilizers (N:P:K = 15%:15%:15%) at a rate of 125 kg ha−1 were supplied when the land was prepared. To control weeds, landscape fabric was used to cover the spaces between the rows. The weeds that appeared within the rows were hand-picked at regular intervals. The recommended dose of pesticides was used to control pests. Quantitative and qualitative characteristics of field-grown accessions were recorded during the second week of October in 2014–2017. The morphological traits of each accession were recorded from five randomly selected plants of each accession. The different quantitative parameters considered for the study included: fresh weight, dry weight, plant height, leaf length, leaf width, number of leaves, leaf color, leaf orientation, ratio of leaf length to leaf width, culm length, number of nodes, tiller number, weight of 1000 seeds, full bloom time, bloom beginning, spike width, and spike length. Qualitative traits such as seed shape and seed color of each accession were rated (visually) and analyzed. The average height of each accession was measured using a ruler from the base of the plant to the apex of the stem. The emergence of flower (bloom beginning) was measured as the time duration (days) between the sowing and the emergence of the first flower. Full bloom was the time duration (days) between planting and emergence of approximately 50% of the flowers from each accession. The average weight of 1000 seeds was measured from 10 randomly selected samples from each P. miliaceum accession.
4.4. Screening of Chlorophyll Contents
To extract chlorophyll a, b, and carotenoid, 0.5 g of leaf sample was taken, and homogenized with 10 mL of 80% acetone. The homogenized sample was centrifuged for 4000 rpm for 10 min at 4 °C. The supernatants were collected in the cuvette. The chlorophyll a, b, and carotenoid content was spectrophotometrically analysed using a Shimadju UV-1800 UV-VS spectrophotometer (Shimadzu, Kyoto, Japan) by following method described previously [
99]. The concentration of chlorophyll-a, chlorophyll-b, and carotenoids was calculated using following equation:
4.5. Screening of Total Phenolic Concentration
The TPC of different extracts was determined using Folin–Ciocalteu’s phenol reagent by following the method described previously [
100]. Briefly, 100 µL of plant extracts (1000 ppm) or standard compounds (gallic acid) were mixed with 200 µL of Folin–Ciocalteu’s phenol reagent and allowed stand for 5 min at room temperature (25 °C). After five minutes, 300 µL of Na
2CO
3 (20%
w/
v) solution was added to the mixture to stop the reaction. After incubating for 40 min at room temperature, the absorbance value was recorded at 765 nm against a blank solution (200 µL of 80% methanol). Gallic acid at various concentrations (1, 5, 10, 100 mg/L) was used for obtaining a calibration curve. Total phenolic contents of samples were expressed as mg gallic acid equivalent (GAE)/g of sample. All experiments were performed in triplicate.
4.6. Screening of Total Flavonoids Concentration
The TFC of different extracts was determined using a spectrophotometric method [
101]. Briefly, 500 mL of plant extract (1000 ppm) or standard (quercetin) were added to 300 µL of AlCl
3 and 200 µL of potassium cyanide. The mixtures were incubated for 30 min and absorbance value of the solution was measured immediately at 510 nm against a blank. The blank consisted of 500 mL of 80% methanol. A calibration curve was drawn using various concentrations of quercetin (1, 3, 5, 10 mg/L) and the TFCs of tested samples were expressed as mg quercetin equivalent (Qu)/G sample. All experiments were performed in triplicate.
4.7. Screening of Antioxidant Activity of P. miliaceum Accessions
4.7.1. Evaluation of DPPH Assay
Antioxidant activity of fifteen
P. miliaceum accessions was determined using a 1,1-diphenyl-2-picryl-hydrazyl radical (DPPH) assay by following the methods described previously [
102]. Briefly, 200 µL of plant crude extracts at different concentrations (ranging from 10,000 ppm to 100 ppm) or standard compounds mixed with 300 µL of 0.004% freshly prepared methanol solution of DPPH in a 96-well plate. The mixture was incubated at room temperature (25 °C) for 30 min in the dark. The absorbance value of each test solution was measured at 515 nm using a V530 UV-VIS spectrophotometer (Jasco, Tokyo, Japan) connected with an automated plate reader. α-tocopherol was used as a positive control. The DPPH scavenging potential of each sample was measured using the following equation:
where Ab
control is the absorbance value of the control reaction (only DPPH radical solution) and Ab
sample is the absorbance value of the plant sample (plant extract mixed with DPPH radical solution).
4.7.2. Evaluation of ABTS Assay
An ABTS assay was performed using a previously described previously [
103] with some modifications. Initially, a working solution of ABTS (7.4 mM) potassium persulfate solution (2.6 mM) was mixed in an equal ratio (1:1,
v/
v) and allowed to react for 12 h at room temperature (25 °C) in the dark. The mixture was then diluted in methanol and the absorbance at 734 nm of the solution was adjusted to 0.600 ± 0.01. Then, 2.5-mL aliquots of extracts were added to 2.5 mL of ABTS solution. After gentle agitation, the solution was allowed to stand for 2 h in the dark. The absorbance of the mixture was measured at 734 nm. A standard curve was generated using a Trolox standard solution at various concentrations (500–1000 µM). The radical scavenging activity of each sample was expressed as mmol Trolox equivalent/100 g. All the experiments were performed in triplicate. The ABTS capacity of each sample was calculated as follows:
where Ab
control is the absorbance of the control reaction (ABTS radical solution with no sample and positive control) and Ab
sample is the absorbance of the test compound (ABTS radical solution with a sample or positive control).
4.8. DNA Extraction
Young leaves each from fifteen
P. miliaceum accessions were collected for DNA extraction. DNA of individual accessions was extracted by following the standard CTAB method [
104]. Approximately one gram of fresh leaf from 15 accessions was crushed individually in liquid nitrogen and suspended with extraction buffer (100 mM Tris-HCl, pH 8.5, 1.4 mM NaCl, 20 mM EDTA, 2% CTAB, and 0.2% β-mercaptoethanol) and incubated in a water bath for 45 min at 65 °C. After incubation, the supernatant was transferred to another 1.5 mL Eppendorf tube and an equal volume of ice-cold chloroform–isoamyl alcohol was added and the mixture were inverted and then spun at 11,952×
g for 10 min. The supernatant was placed into another Eppendorf tube and an equal volume of ice-cold isopropanol was added to the mixture and incubated at −20 °C for 40 min. Then, the tube was centrifuged at 17,226×
g for 10 min. After discarding the supernatant, 70% ice-cold ethanol was added to the DNA pellet. The DNA pellet was air dried and dissolved in 30 µL of nuclease-free water. The quality and concentration of DNA genomic DNA of all accessions was determined using a UV-VIS spectrophotometer (Jasco V530 UV-VIS) and 0.8% agarose gel electrophoresis. Extracted DNA was diluted to 5 ng/µL using 1 mmol/L TE buffer.
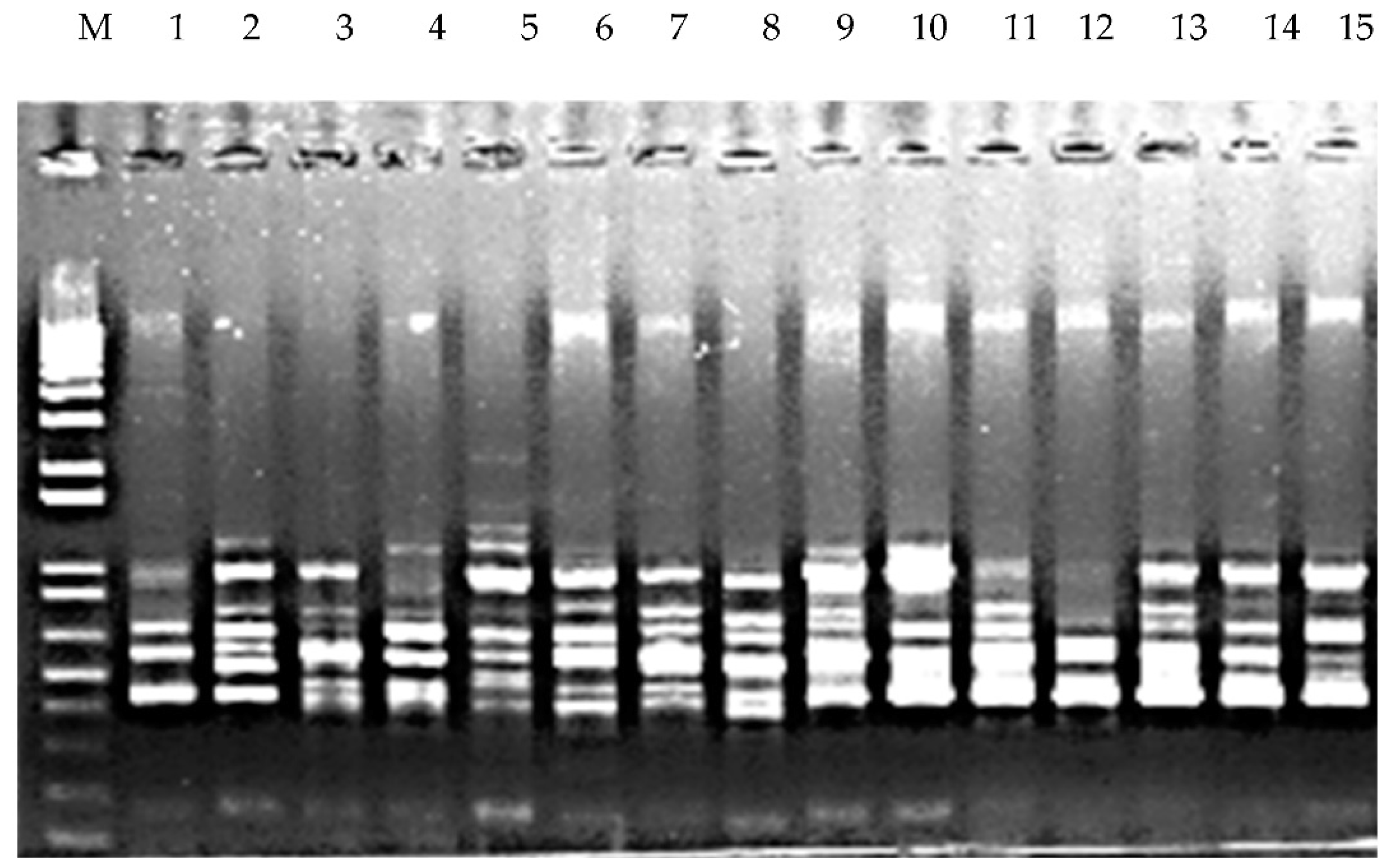
PCR Amplification and Electrophoresis
A total of 29 ISSR primers used to amplify the DNA of 15 accessions of P. miliaceum were obtained from the Bioherb Research Institute, Kangwon National University, South Korea. A total of 15 highly polymorphic ISSR markers were selected for this study. PCR amplification for the genomic DNA of each accession was performed in a 20-µL reaction volume containing 50 ng genomic DNA, 2 µL 1X PCR buffer, 1 U Taq DNA polymerase; 1 µM of primer, and 300 µM of dNTPs. PCR conditions for DNA amplification were as follows: initial denaturation step for 4 min at 95 °C, followed by 45 cycles of denaturation for 30 s at 94 °C, primer annealing for 45 s at 48–52 °C, and extension for 2 min at 72 °C, followed by a final extension for 10 min at 72 °C. The amplified products were loaded onto the 1% gel in 0.5X TBE buffer. The amplified PCR products were electrophoretically separated on 0.8% agarose in 0.5X TBE buffer for 20 min at 25 V and observed under UV light.
A total of eleven RAPD primers used to amplify the DNA of 15 accessions of P. miliaceum were obtained from Bioherb Research Institute (Kangwon National University, Chuncheon, South Korea). RAPD amplification reactions were performed in a 20-µL reaction volume containing 50 ng genomic DNA, 2 µL 1X PCR buffer, 1 U Taq DNA polymerase, 1 µM of primer, and 300 µM of dNTPs obtained from Bioneer (Daejeon, South Korea). The amplification conditions were an initial denaturation at 94 °C for 5 min; 45 cycles of 30 s at 94 °C, 1 min at 42 °C, and 2 min at 72 °C; a final extension of 5 min at 72 ° C, followed by storage at 4 °C.
4.9. Statistical Analysis
Each experiment was performed in triplicate. The data obtained from the experiments were expressed as mean ± standard deviation. Quantitative data were statistically analyzed using one-way analysis of variance (ANOVA). Significant differences between the obtained data were determined by using Duncan’s multiple range test at
p < 0.05 (SPSS ver. 20.0, SPSS Inc., Chicago, IL, USA). Correlations among the morphological traits, antioxidant activities, total phenolic content, and total flavonoid content were obtained by calculating Pearson’s correlation coefficient using SPSS software ver. 20.0. Principal component analysis (PCA) of quantitative morphological traits was performed using SPSS software ver. 20.0. Cluster analysis using the unweighted pair group method with arithmetic averages (UPGMA) was performed using SPSS software ver. 20. Shannon’s information index (I), number of effective alleles (Ne), number of observed alleles (Na), expected heterozygosity (He), and unbiased expected heterozygosity (uHe) were determined using GenAlEx software. NTSYS software v. 2.00 [
105] (Exeter Publishing, Ltd., Setauket, NY, USA) was used to generate a UPGMA dendrogram.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}