Efficient Conversion of Cane Molasses Towards High-Purity Isomaltulose and Cellular Lipid Using an Engineered Yarrowia lipolytica Strain in Fed-Batch Fermentation

Abstract
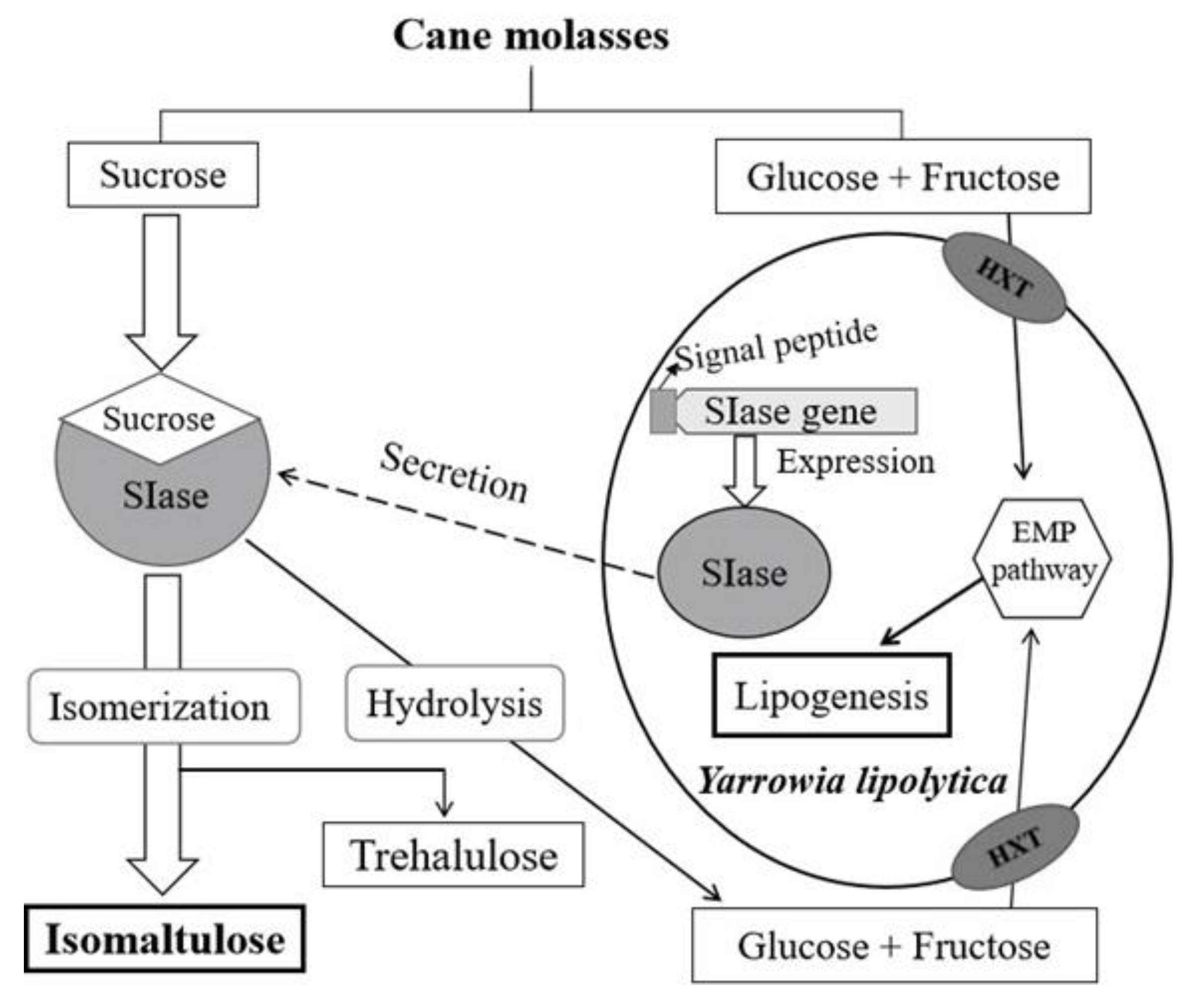
:1. Introduction
2. Results and Discussion
2.1. Isomaltulose Production Using PCM as Sole Carbon Source
2.2. Effect of CSL on Enhancement of Isomaltulose and Lipid Production
2.3. Fed-Batch Fermentation for Isomaltulose and Lipid
2.4. Fatty Acids Composition of Intracellular Lipids and Biodiesel Production
3. Materials and Methods
3.1. Strain and Cane Molasses Fermentation
3.2. CSL Optimization for Enhancement of Isomaltulose and Lipid Co-Production
3.3. Fed-Batch Fermentation in a 10-L Fermentor
3.4. Enzyme Assay and Determination for Isomaltulose and Lipid Contents, Residual Sugar
3.5. Determination of Lipid Composition and Preparation for Biodiesel
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Qiang, X.F.; Luo, J.Q.; Guo, S.W.; Cao, W.F.; Hang, X.F.; Liu, J.S.; Wan, Y.H. A novel process for molasses utilization by membrane filtration and resin adsorption. J. Clean. Prod. 2019, 207, 432–443. [Google Scholar] [CrossRef]
- Wu, L.T.; Wu, S.S.; Qiu, J.J.; Xu, C.X.; Li, S.; Xu, H. Green synthesis of isomaltulose from cane molasses by Bacillus subtilis WB800-pHA01-palI in a biologic membrane reactor. Food Chem. 2017, 229, 761–768. [Google Scholar] [CrossRef]
- Yin, F.W.; Zhu, S.Y.; Guo, D.S.; Ren, L.J.; Ji, X.J.; Huang, H.; Gao, Z. Development of a strategy for the production of docosahexaenoic acid by Schizochytrium sp. from cane molasses and algae-residue. Bioresour. Technol. 2019, 271, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Khan, E.U.; Pasha, T.N.; Aleem, M.; Rashid, M.A. Replacing corn grains with varying levels of cane molasses, post weaning intake, growth performance and structural development of Nili Ravi buffalo (Bubalus bubalis) female calves. J. Anim. Plant Sci. 2018, 28, 1002–1006. [Google Scholar]
- Senthilkumar, S.; Suganya, T.; Deepa, K.; Muralidharan, J.; Sasikala, K. Supplementation of molasses in livestock feed. Int. J. Environ. Sci. Technol. 2016, 5, 1243–1250. [Google Scholar]
- Liu, J.; Sun, Z.; Zhong, Y.J.; Gerken, H.; Huang, J.C.; Chen, F. Utilization of cane molasses towards cost-saving astaxanthin production by a Chlorella zofingiensis mutant. J. Appl. Phycol. 2013, 25, 1447–1456. [Google Scholar] [CrossRef]
- Howard, J.; Rackemann, D.W.; Bartley, J.P.; Samori, C.; Doherty, W.O.S. Conversion of sugar cane molasses to 5-hydroxymethylfurfural using molasses and bagasse-derived catalysts. ACS Sustain. Chem. Eng. 2018, 6, 4531–4538. [Google Scholar] [CrossRef]
- Fadel, M.; Keera, A.A.; Mouafi, F.E.; Kahil, T. High level ethanol from sugar cane molasses by a new thermotolerant Saccharomyces cerevisiae strain in industrial scale. Biotechnol. Res. Int. 2013, 2013, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Muñiz, B.; Carvajal-Zarrabal, O.; Torrestiana-Sanchez, B.; Aguilar-Uscanga, M.G. Kinetic study on ethanol production using Saccharomyces cerevisiae ITV-01 yeast isolated from sugar cane molasses. J. Chem. Technol. Biotechnol. 2010, 85, 1361–1367. [Google Scholar] [CrossRef]
- Feng, X.H.; Tang, B.; Jiang, Y.X.; Xu, Z.Q.; Lei, P.; Liang, J.F.; Xu, H. Efficient production of poly-γ-glutamic acid from cane molasses by Bacillus subtilis NX-2 immobilized on chemically modified sugarcane bagasse. J. Chem. Technol. Biotechnol. 2016, 91, 2085–2093. [Google Scholar] [CrossRef]
- Mladenović, D.D.; Djukić-Vuković, A.P.; Kocić-Tanackov, S.D.; Pejin, J.D.; Mojović, L.V. Lactic acid production on a combined distillery stillage and sugar beet molasses substrate. J. Chem. Technol. Biotechnol. 2016, 91, 2474–2479. [Google Scholar] [CrossRef]
- Wang, Z.; Li, H.; Feng, J.; Zhang, A.; Ying, H.; He, X.; Jiang, M.; Chen, K.Q.; Ouyang, P.K. Enhanced succinic acid production from polyacrylamide-pretreated cane molasses in microbial electrolysis cells. J. Chem. Technol. Biotechnol. 2018, 93, 855–860. [Google Scholar] [CrossRef]
- Rocky-Salimi, K.; Hashemi, M.; Safari, M.; Mousivand, M. Valorisation of untreated cane molasses for enhanced phytase production by Bacillus subtilis K46b and its potential role in dephytinisation. J. Sci. Food Agric. 2017, 97, 222–229. [Google Scholar] [CrossRef]
- Sawale, P.D.; Shendurse, A.M.; Mohan, M.S.; Patil, G.R. Isomaltulose (palatinose)—An emerging carbohydrate. Food Biosci. 2017, 18, 46–52. [Google Scholar] [CrossRef]
- Goulter, K.C.; Hashimi, S.M.; Birch, R.G. Microbial sucrose isomerases, producing organisms, genes and enzymes. Enzyme Microb. Tech. 2012, 50, 57–64. [Google Scholar] [CrossRef]
- Wu, L.G.; Birch, R.G. Characterization of the highly efficient sucrose isomerase from Pantoea dispersa UQ68J and cloning of the sucrose isomerase gene. Appl. Environ. Microb. 2005, 71, 1581–1590. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Cai, H.; Qing, Y.J.; Ren, B.; Xu, H.; Zhu, H.Y.; Yao, J. Cloning and characterization of a sucrose isomerase from Erwinia rhapontici NX-5 for isomaltulose hyperproduction. Appl. Biochem. Biotech. 2011, 163, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Jung, J.H.; Seo, D.H.; Ha, S.J.; Yoon, J.W.; Kim, Y.C.; Shim, J.H.; Park, C.S. Microbial production of palatinose through extracellular expression of a sucrose isomerase from Enterobacter sp. FMB-1 in Lactococcus lactis MG1363. Bioresour. Technol. 2010, 101, 8828–8833. [Google Scholar] [CrossRef] [PubMed]
- Kawaguti, H.Y.; Sato, H.H. Isomaltulose production by free cells of Serratia plymuthica in a batch process. Food Chem. 2010, 120, 789–793. [Google Scholar] [CrossRef]
- Orsi, D.C.; Sato, H.H. Isomaltulose production using free and immobilized Serratia plymuthica cells. Afr. J. Biotechnol. 2016, 15, 835–842. [Google Scholar]
- Lee, G.Y.; Jung, J.H.; Seo, D.H.; Hansin, J.; Ha, S.J.; Cha, J.; Kim, Y.S.; Park, C.S. Isomaltulose production via yeast surface display of sucrose isomerase from Enterobacter sp. FMB-1 on Saccharomyces cerevisiae. Bioresour. Technol. 2011, 102, 9179–9184. [Google Scholar] [CrossRef]
- Li, L.J.; Wang, H.W.; Cheng, H.R.; Deng, Z.X. Isomaltulose production by yeast surface display of sucrose isomerase from Pantoea dispersa on Yarrowia lipolytica. J. Funct. Foods 2017, 32, 208–217. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, Z.P.; Sheng, J.; Zheng, Y.; Ji, X.F.; Zhou, H.X.; Liu, X.Y.; Chi, Z.M. High and efficient isomaltulose production using an engineered Yarrowia lipolytica strain. Bioresour. Technol. 2018, 265, 577–580. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wang, Z.P.; Liu, S.; Wang, Y.L.; Zhang, Z.F.; Liu, X.M.; Du, Y.M.; Yuan, X.L. Overexpression of secreted sucrose isomerase in Yarrowia lipolytica and its application in isomaltulose production after immobilization. Int. J. Biol. Macromol. 2019, 121, 97–103. [Google Scholar] [CrossRef]
- Qiao, K.J.; Abidi, S.H.I.; Liu, H.J.; Zhang, H.R.; Chakraborty, S.; Watson, N.; Ajikumar, P.K.; Stephanopoulos, G. Engineering lipid overproduction in the oleaginous yeast Yarrowia lipolytica. Metab. Eng. 2015, 29, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.R.; Chi, Z.; Jiang, H.; Liu, G.L.; Xue, S.J.; Hu, Z.; Chi, Z.M. Overexpression of a pyruvate carboxylase gene enhances extracellular liamocin and intracellular lipid biosynthesis by Aureobasidium melanogenum M39. Process Biochem. 2018, 69, 64–74. [Google Scholar] [CrossRef]
- Liu, X.Y.; Wang, X.F.; Xu, J.X.; Xia, J.; Lv, J.S.; Zhang, T.; Wu, Z.; Deng, Y.F.; He, J.L. Citric acid production by Yarrowia lipolytica SWJ-1b using corn steep liquor as a source of organic nitrogen and vitamins. Ind. Crop Prod. 2015, 78, 154–160. [Google Scholar] [CrossRef]
- Amado, I.R.; Vázquez, J.A.; Pastrana, L.; Teixeira, J.A. Microbial production of hyaluronic acid from agro-industrial by-products, molasses and corn steep liquor. Biochem. Eng. J. 2017, 117, 181–187. [Google Scholar] [CrossRef]
- Cheng, C.; Zhou, Y.P.; Lin, M.; Wei, P.L.; Yang, S.T. Polymalic acid fermentation by Aureobasidium pullulans for malic acid production from soybean hull and soy molasses, fermentation kinetics and economic analysis. Bioresour. Technol. 2017, 223, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; Lv, J.S.; Xu, J.X.; Xia, J.; He, A.Y.; Zhang, T.; Li, X.Q.; Xu, J.M. Effects of osmotic pressure and pH on citric acid and erythritol production from waste cooking oil by Yarrowia lipolytica. Eng. Life Sci. 2018, 18, 344–352. [Google Scholar] [CrossRef]
- Li, S.; Xu, H.; Yu, J.G.; Wang, Y.Y.; Feng, X.H.; Ouyang, P.K. Enhancing isomaltulose production by recombinant Escherichia coli producing sucrose isomerase, culture medium optimization containing agricultural wastes and cell immobilization. Bioprocess Biosyst. Eng. 2013, 36, 1395–1405. [Google Scholar] [CrossRef]
- Jiru, T.M.; Steyn, L.; Pohl, C.; Abate, D. Production of single cell oil from cane molasses by Rhodotorula kratochvilovae (syn, Rhodosporidium kratochvilovae) SY89 as a biodiesel feedstock. Chem. Cent. J. 2018, 12, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Martínez, P.; Buey, R.M.; Ledesma-Amaro, R.; Jiménez, A.; Revuelta, J.L. Engineering Ashbya gossypii strains for de novo lipid production using industrial by-products. Microb. Biotechnol. 2017, 10, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.P.; Fu, W.J.; Xu, H.M.; Chi, Z.M. Direct conversion of inulin into cell lipid by an inulinase-producing yeast Rhodosporidium toruloides 2F5. Bioresour. Technol. 2014, 161, 131–136. [Google Scholar] [CrossRef]
- Wang, S.K.; Wang, X.; Tao, H.H.; Sun, X.S.; Tian, Y.T. Heterotrophic culture of Chlorella pyrenoidosa using sucrose as the sole carbon source by co-culture with immobilized yeast. Bioresour. Technol. 2018, 249, 425–430. [Google Scholar] [CrossRef]
- Taskin, M.; Ortucu, S.; Aydogan, M.N.; Arslan, N.P. Lipid production from sugar beet molasses under non-aseptic culture conditions using the oleaginous yeast Rhodotorula glutinis TR29. Renew. Energy 2016, 99, 198–204. [Google Scholar] [CrossRef]
- Katre, G.; Joshi, C.; Khot, M.; Zinjarde, S.; Ravikumar, A. Evaluation of single cell oil (SCO) from a tropical marine yeast Yarrowia lipolytica NCIM 3589 as a potential feedstock for biodiesel. AMB Express 2012, 2, 36–50. [Google Scholar] [CrossRef]
- Wang, Q.; Guo, F.J.; Rong, Y.J.; Chi, Z.M. Lipid production from hydrolysate of cassava starch by Rhodosporidium toruloides 21167 for biodiesel making. Renew. Energy 2012, 46, 164–168. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |



{kind=link}
{kind=link}
| PCM (g L–1) | Isomaltulose (g L−1) | SIase (U mL−1) | Lipid Content (g L−1) | Biomass (g L−1) | Residual Sugar (g L–1) |
|---|---|---|---|---|---|
| 200 | 58.6 ± 2.3 | 3.3 ± 0.2 | 3.8 ± 0.2 | 9.9 ± 0.7 | 1.3 ± 0.1 |
| 250 | 73.3 ± 7.2 | 3.3 ± 0.1 | 4.6 ± 0.3 | 11.8 ± 0.6 | 1.6 ± 0.1 |
| 300 | 88.1 ± 6.6 | 3.6 ± 0.1 | 5.2 ± 0.2 | 12.6 ± 1.2 | 1.9 ± 0.2 |
| 350 | 96.7 ± 4.9 | 3.6 ± 0.2 | 5.3 ± 0.3 | 12.3 ± 0.5 | 8.6 ± 0.4 |
| 400 | 71.9 ± 6.3 | 3.5 ± 0.2 | 4.8 ± 0.2 | 11.8 ± 0.9 | 42.8 ± 2.7 |
| CSL (g L−1) | Isomaltulose (g L−1) | SIase (U mL−1) | Lipid Content (g L−1) | Biomass (g L−1) | Residual Sugar (g L−1) |
|---|---|---|---|---|---|
| Control | 96.7 ± 7.2 | 3.6 ± 0.2 | 5.3 ± 0.3 | 12.3 ± 0.4 | 8.6 ± 0.5 |
| 0.5 | 102.6 ± 5.4 | 3.9 ± 0.1 | 6.7 ± 0.4 | 14.4 ± 0.8 | 2.3 ± 0.1 |
| 1.0 | 102.6 ± 4.9 | 4.7 ± 0.1 | 7.2 ± 0.3 | 16.1 ± 0.6 | 2.3 ± 0.1 |
| 1.5 | 102.6 ± 6.7 | 4.9 ± 0.2 | 6.2 ± 0.4 | 19.7 ± 0.7 | 2.3 ± 0.1 |
| Strains | Substrate | Isomaltulose Production (g L−1) | Yield (g g−1) | Isomaltulose Proportion (%) | Other Products (g L−1) | References |
|---|---|---|---|---|---|---|
| L. lactis | Sucrose | 36 | 0.72 | <90 | - | [18] |
| S. cerevisiae | Sucrose | <4 | 0.074 | <10 | - | [21] |
| Y. lipolytica | Sucrose | 572.1 | 0.96 | 97.8 | Lipid, 8.1 | [23] |
| Y. lipolytica | Sucrose | 620.7 | 0.96 | - | - | [24] |
| S. plymuthica | Molasses | <33.5 | 0.84 | 80.4 | - | [20] |
| B. subtilis | Molasses | 212.6 | 0.92 | <92.4 | - | [2] |
| Y. lipolytica | Molasses | 161.2 | 0.96 | 97.4 | Lipid, 12.2 | This study |
| Fatty Acids | C14:0 | C16:0 | C16:1 | C18:0 | C18:1 | C18:2 |
|---|---|---|---|---|---|---|
| Percentage (%) | 5.71 | 16.55 | 12.67 | 6.82 | 45.14 | 13.11 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.-P.; Wang, Q.-Q.; Liu, S.; Liu, X.-F.; Yu, X.-J.; Jiang, Y.-L. Efficient Conversion of Cane Molasses Towards High-Purity Isomaltulose and Cellular Lipid Using an Engineered Yarrowia lipolytica Strain in Fed-Batch Fermentation. Molecules 2019, 24, 1228. https://doi.org/10.3390/molecules24071228
Wang Z-P, Wang Q-Q, Liu S, Liu X-F, Yu X-J, Jiang Y-L. Efficient Conversion of Cane Molasses Towards High-Purity Isomaltulose and Cellular Lipid Using an Engineered Yarrowia lipolytica Strain in Fed-Batch Fermentation. Molecules. 2019; 24(7):1228. https://doi.org/10.3390/molecules24071228
Chicago/Turabian StyleWang, Zhi-Peng, Qin-Qing Wang, Song Liu, Xiao-Fang Liu, Xin-Jun Yu, and Yun-Lin Jiang. 2019. "Efficient Conversion of Cane Molasses Towards High-Purity Isomaltulose and Cellular Lipid Using an Engineered Yarrowia lipolytica Strain in Fed-Batch Fermentation" Molecules 24, no. 7: 1228. https://doi.org/10.3390/molecules24071228
APA StyleWang, Z. -P., Wang, Q. -Q., Liu, S., Liu, X. -F., Yu, X. -J., & Jiang, Y. -L. (2019). Efficient Conversion of Cane Molasses Towards High-Purity Isomaltulose and Cellular Lipid Using an Engineered Yarrowia lipolytica Strain in Fed-Batch Fermentation. Molecules, 24(7), 1228. https://doi.org/10.3390/molecules24071228