Smoke-Isolated Karrikins Stimulated Tanshinones Biosynthesis in Salvia miltiorrhiza through Endogenous Nitric Oxide and Jasmonic Acid


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
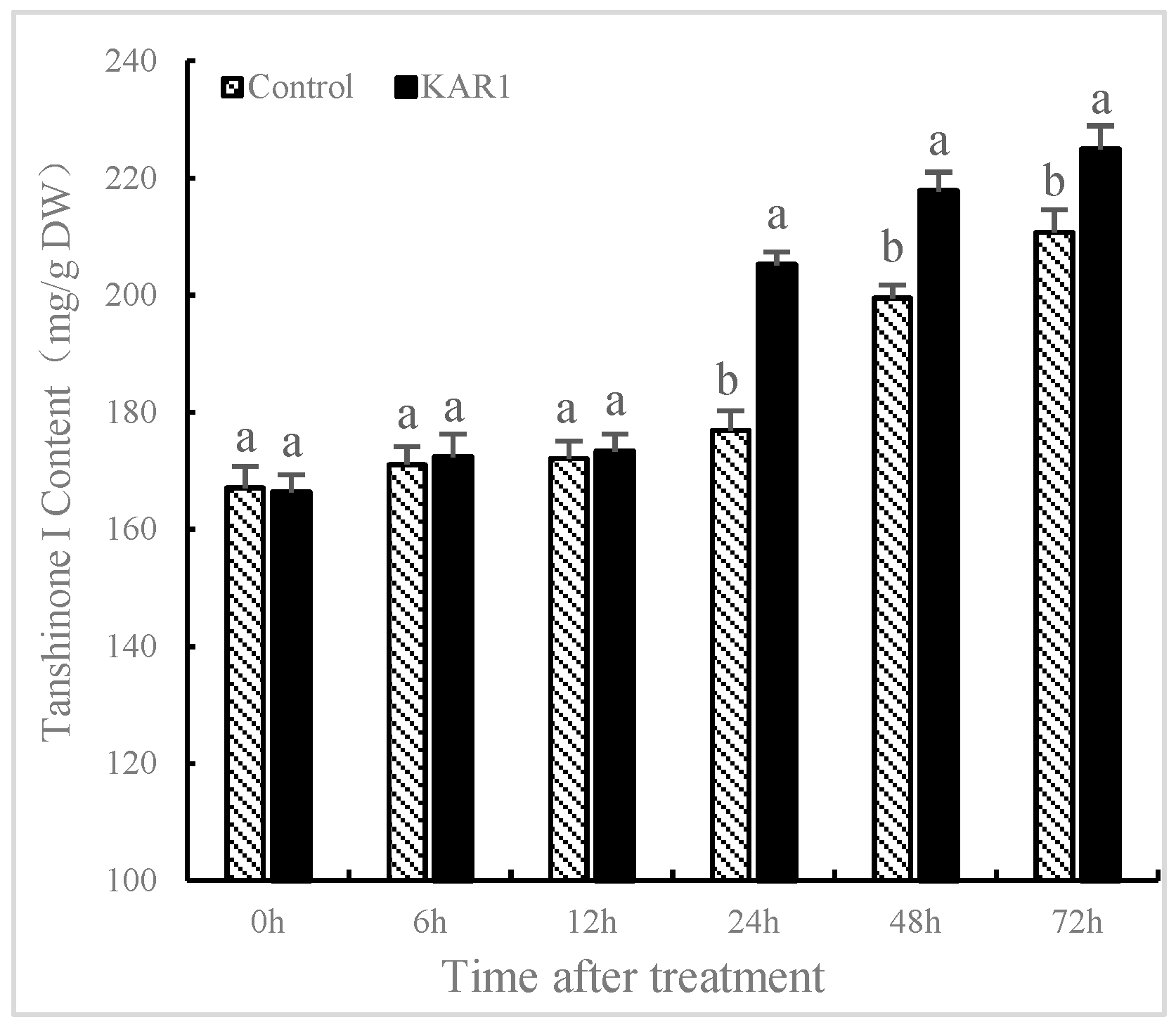
2.1. KAR1-Induced Increasing of T-I in S. miltiorrhiza Hairy Root
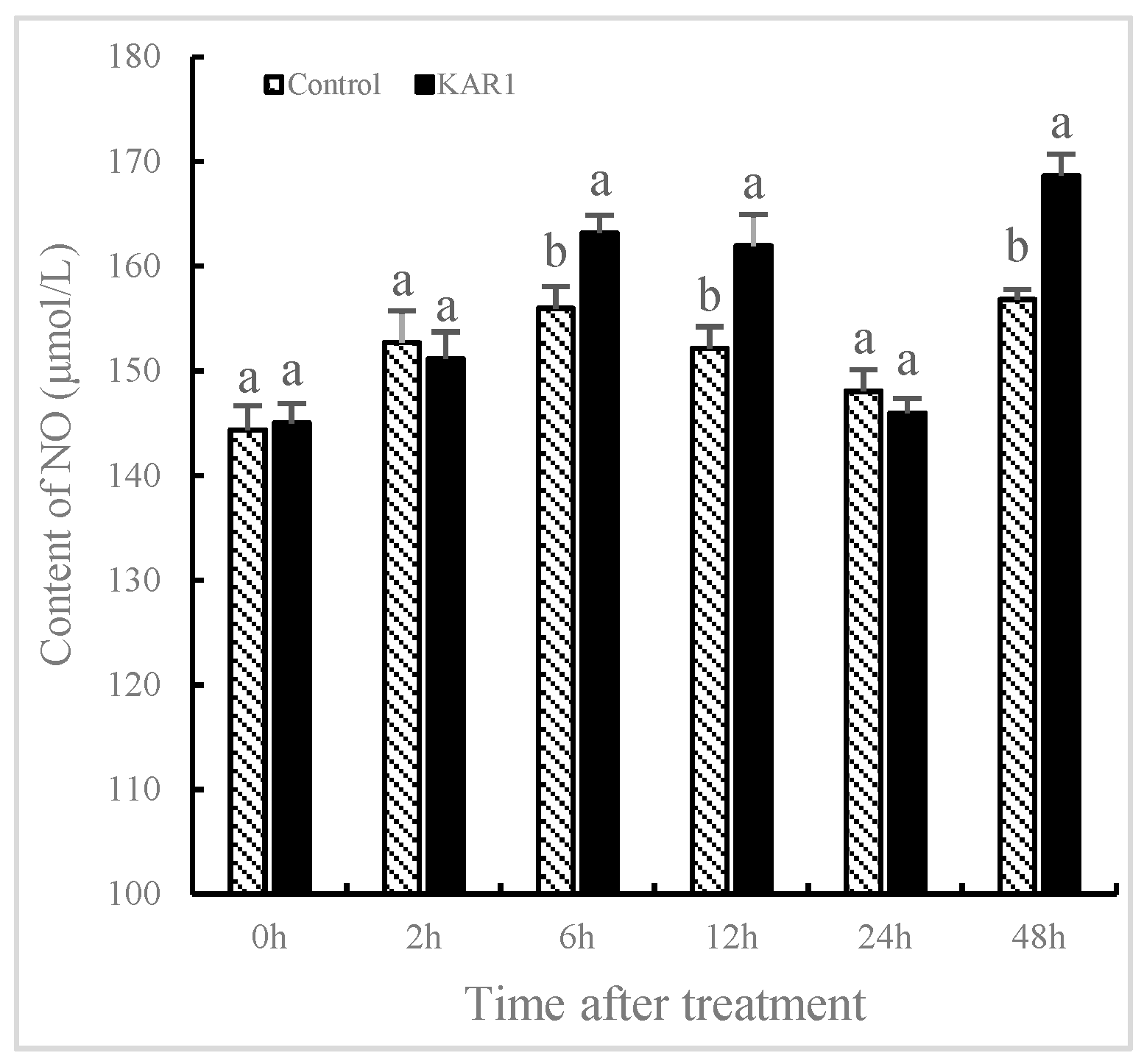
2.2. Burst of NO and JA Induced by KAR1
2.3. JA Acted as a Downstream Signal of NO Pathway Induced by KAR1
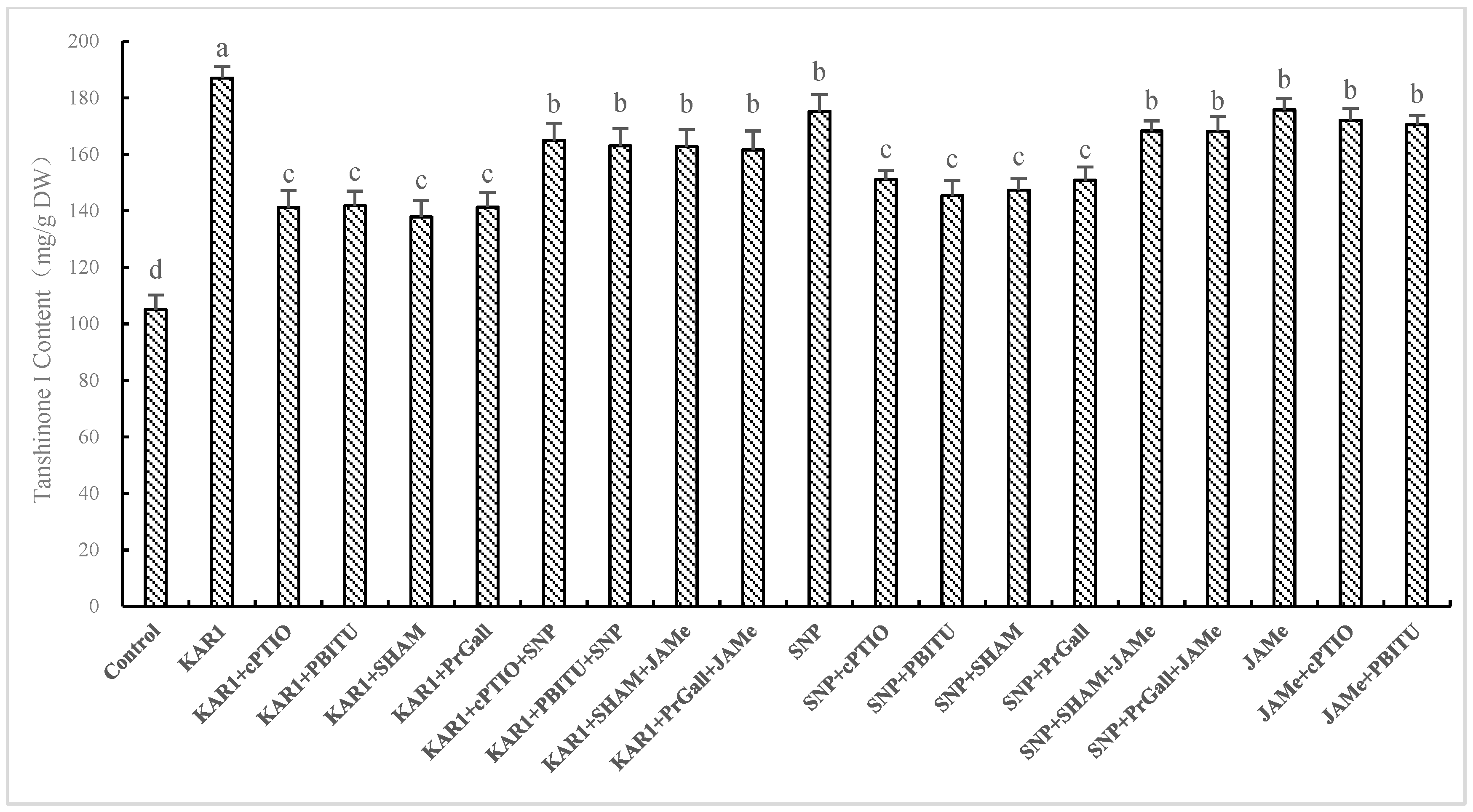
2.4. Dependence of KAR1-Stimulated T-I Production on NO Accumulation as well as JA production
3. Materials and Methods
3.1. Hairy Root Culture and Experimental Design
3.2. Preparation of KAR1 Solution
3.3. HPLC Analysis of T-I
3.4. Determination of NO
3.5. Measurement of JA
3.6. Statistical analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zhou, J.; Van Staden, J.; Guo, L.P.; Huang, L.Q. Smoke-water improves shoot growth and indigo accumulation in shoots of Isatis indigotica seedlings. S. Afr. J. Bot. 2011, 77, 787–789. [Google Scholar] [CrossRef]
- Flematti, G.R.; Ghisalberti, E.L.; Dixon, K.W.; Trengove, R.D. A compound from smoke that promotes seed germination. Science 2004, 305, 977. [Google Scholar] [CrossRef]
- Van Staden, J.; Jäger, A.K.; Light, M.E.; Burger, B.V.; Brown, N.A.C.; Thomas, T.H. Isolation of the major germination cue from plant-derived smoke. S. Afr. J. Bot. 2004, 70, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Krawczyk, E.; Koprowski, M.; Cembrowska-lech, D.; Wójcik, A.; Kępczyński, J. Synthesis of tricyclic butenolides and comparison their effects with known smoke-butenolide, KAR1. J. Plant Physiol. 2017, 215, 91–99. [Google Scholar] [CrossRef]
- López-Ráez, J.A.; Shirasu, K.; Foo, E. Strigolactones in Plant Interactions with Beneficial and Detrimental Organisms: The Yin and Yang. Trends Plant Sci. 2017, 22, 527–537. [Google Scholar] [CrossRef]
- Salomon, M.V.; Piccoli, P.; Funes, I.P.; Stirk, W.A.; Kulkarni, M.; Van staden, J.; Bottini, R. Bacteria and smoke-water extract improve growth and induce the synthesis of volatile defense mechanisms in Vitis vinifera L. Plant Physiol. Biochem. 2017, 120, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Baniela, M.; Carlon, L.; Diaz, T.E.; Bueno, A.; Fernandez-Pascual, E. Plant-derived smoke and temperature effects on seed germination of five Helianthemum (Cistaceae). Flora 2016, 223, 56–61. [Google Scholar] [CrossRef]
- Morffy, N.; Faure, L.; Nelson, D.C. Smoke and Hormone Mirrors: Action and Evolution of Karrikin and Strigolactone Signaling. Trends Genet. 2016, 32, 176–188. [Google Scholar] [CrossRef] [PubMed]
- China Pharmacopoeia Committee. Pharmacopoeia of Peoples Republic of China; China Medical Science and Technology Press: Beijing, China, 2015; p. 76. [Google Scholar]
- Yu, Y.; Zhang, W.B.; Li, X.Y.; Piao, X.C.; Jiang, J.; Lian, M.L. Pathogenic fungal elicitors enhance ginsenoside biosynthesis of adventitious roots in Panax quinquefolius during bioreactor culture. Ind. Crop Prod. 2016, 94, 729–735. [Google Scholar] [CrossRef]
- Wang, Y.J.; Shen, Y.; Shen, Z.; Zhao, L.; Ning, D.L.; Jiang, C.; Zhao, R.; Huang, L.Q. Comparative proteomic analysis of the response to silver ions and yeast extract in salvia miltiorrhiza hairy root cultures. Plant Physiol. Biochem. 2016, 107, 364–373. [Google Scholar] [CrossRef]
- Wu, C.F.; Bohnert, S.; Thines, E.; Efferth, T. Cytotoxicity of salvia miltiorrhiza against multidrug-resistant cancer cells. Am. J. Chin. Me. 2016, 44, 871–894. [Google Scholar] [CrossRef]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef]
- Lu, X.J.; Zhang, X.L.; Mei, M.; Liu, G.L.; Ma, B.B. Proteomic analysis of Magnolia sieboldii K. Koch seed germination. J. Proteomics. 2016, 133, 76–85. [Google Scholar] [CrossRef]
- Cembrowska-lech, D.; Koprowski, M.; Kępczyński, J. Germination induction of dormant Avena fatua caryopses by KAR1 and GA3 involving the control of reactive oxygen species (H2O2 and O2•−) and enzymatic antioxidants (superoxide dismutase and catalase) both in the embryo and the aleurone layers. J. Plant Physiol. 2015, 176, 169–179. [Google Scholar] [CrossRef]
- Ren, C.G.; Dai, C.C. Nitric oxide and brassinosteroids mediated fungal endophyte-lnduced volatile oil production through protein phosphorylation pathways in Atractylodes lancea plantlets. J. Integr. Plant Biol. 2013, 55, 1136–1146. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.Z.; Li, M.Y.; Jin, H.H.; Sun, L.N.; Zhu, Y.; Xu, M.J.; Dong, J.F. UV-B irradiation alleviates the deterioration of cold-stored mangoes by enhancing endogenous nitric oxide levels. Food Chem. 2015, 169, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Dong, L.X.; Jiang, Z.F.; Yang, X.L.; Sun, Z.H.; Li, J.X.; Wu, Y.H.; Xu, M.J. Salicylic acid-induced flavonoid accumulation in Ginkgo biloba leaves is dependent on red and far-red light. Ind. Crop Prod. 2018, 118, 102–110. [Google Scholar] [CrossRef]
- Xu, M.J.; Dong, J.F.; Ming, Z.; Xu, X.B.; Sun, L.N. Cold-induced endogenous nitric oxide generation plays a role in chilling tolerance of loquat fruit during postharvest storage. Postharvest Biol. Tec. 2012, 65, 5–12. [Google Scholar] [CrossRef]
- Afrin, S.; Huang, J.J.; Luo, Z.Y. JA-mediated transcriptional regulation of secondary metabolism in medicinal plants. Sci. Bull. 2015, 60, 1062. [Google Scholar] [CrossRef]
- Pei, T.L.; Ma, P.D.; Ding, K.; Liu, S.J.; Jia, Y.Y.; Ru, M.; Dong, J.N.; Liang, Z.S. SmJAZ8 acts as a core repressor regulating JA-induced biosynthesis of salvianolic acids and tanshinones in Salvia miltiorrhiza hairy roots. J. Exp. Bot. 2018, 69, 1663–1678. [Google Scholar] [CrossRef]
- Ming, Q.L.; Han, T.; Li, W.C.; Zhang, Q.Y.; Zhang, H.; Zheng, C.J.; Huang, F.; Rahman, K.; Qin, L.P. Tanshinone IIA and tanshinone I production by Trichoderma atroviride D16, an endophytic fungus in Salvia miltiorrhiza. Phytomedicine 2012, 19, 330–333. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.C.; Wu, J.Y. Tanshinone production and isoprenoid pathways in Salvia miltiorrhiza hairy roots induced by Ag+ and yeast elicitor. Plant Sci. 2005, 168, 487–491. [Google Scholar] [CrossRef]
- Zhou, J.; Guo, L.P.; Zhang, J.; Zhou, S.F.; Yang, G.; Zhao, M.Q.; Huang, L.Q. Effects of LaCl3 on photosynthesis and the accumulation of tanshinones and salvianolic acids in Salvia miltiorrhiza seedlings. J. Rare Earth. 2011, 29, 494–498. [Google Scholar] [CrossRef]
- Gao, W.; Sun, H.X.; Xiao, H.B.; Cui, G.H.; Hillwig, M.L.; Jackson, A.; Wang, X.; Shen, Y.; Zhou, N.; Zhang, L.X.; et al. Combining metabolomics and transcriptomics to characterize tanshinone biosynthesis in Salvia miltiorrhiza. BMC Genomics. 2014, 15, 73. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.H.; Zheng, L.P.; Tian, H.; Wang, J.W. Synergistic effects of ultraviolet-B and methyl jasmonate on tanshinone biosynthesis in Salvia miltiorrhiza hairy roots. J. Photochem. Photobiol. B 2016, 159, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Aremu, A.O.; Masondo, N.A.; Van Staden, J. Smoke–water stimulates secondary metabolites during in vitro seedling development in Tulbaghia species. S. Afr. J. Bot. 2014, 9, 49–52. [Google Scholar] [CrossRef]
- Soós, V.; Sebestyén, E.; Juhász, A.; Szalai, G.; Tandori, J.; Light, M.E.; Kohout, L.; Van Staden, J.; Balázs, E. Transcriptome analysis of germinating maize kernels exposed to smoke–water and the active compound KAR1. BMC Plant Biol. 2010, 10, 236. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.J.; Dong, J.F. Synergistic action between jasmonic acid and nitric oxide in inducing matrine accumulation of Sophora flavescens suspension cells. J. Integr. Plant Biol. 2008, 50, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.H.; Liao, Y.C.; Zhang, Z.; Liu, J.; Sun, P.W.; Gao, Z.H.; Sui, C.; Wei, J.H. Jasmonic acid is a crucial signal transducer in heat shock induced sesquiterpene formation in Aquilaria sinensis. Sci. Rep. 2016, 6, 21843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Yan, Y.; Wang, B.; Liang, Z.; Liu, Y.; Liu, F. Selective responses of enzymes in the two parallel pathways of rosmarinic acid biosynthetic pathway to elicitors in Salvia miltiorrhiza hairy root cultures. J. Biosci. Bioeng. 2014, 117, 645–651. [Google Scholar] [CrossRef]
- Light, M.E.; Daws, M.I.; Van Staden, J. Smoke-derived butenolide: Towards understanding its biological effects. S. Afr. J. Bot. 2009, 75, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Liang, W.; Chen, W.; Wu, L.; Li, S.; Qi, Q.; Cui, Y.; Liang, L.; Ye, T.; Zhang, L. Quality Evaluation and Chemical Markers Screening of Salvia miltiorrhiza Bge. (Danshen) Based on HPLC Fingerprints and HPLC-MS (n) Coupled with Chemometrics. Molecules 2017, 22, 2–16. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, L.; Ahammed, G.J.; Li, Z.X.; Wei, J.P.; Shen, C.; Yan, P.; Zhang, L.P.; Han, W.Y. Nitric oxide mediates brassinosteroid-induced flavonoid biosynthesis in Camellia sinensis L. J. Plant Physiol. 2017, 214, 145–151. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds (tanshinone I) are available from the authors. |






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, J.; Xu, Z.-x.; Sun, H.; Guo, L.-p. Smoke-Isolated Karrikins Stimulated Tanshinones Biosynthesis in Salvia miltiorrhiza through Endogenous Nitric Oxide and Jasmonic Acid. Molecules 2019, 24, 1229. https://doi.org/10.3390/molecules24071229
Zhou J, Xu Z-x, Sun H, Guo L-p. Smoke-Isolated Karrikins Stimulated Tanshinones Biosynthesis in Salvia miltiorrhiza through Endogenous Nitric Oxide and Jasmonic Acid. Molecules. 2019; 24(7):1229. https://doi.org/10.3390/molecules24071229
Chicago/Turabian StyleZhou, Jie, Zi-xin Xu, Hui Sun, and Lan-ping Guo. 2019. "Smoke-Isolated Karrikins Stimulated Tanshinones Biosynthesis in Salvia miltiorrhiza through Endogenous Nitric Oxide and Jasmonic Acid" Molecules 24, no. 7: 1229. https://doi.org/10.3390/molecules24071229
APA StyleZhou, J., Xu, Z. -x., Sun, H., & Guo, L. -p. (2019). Smoke-Isolated Karrikins Stimulated Tanshinones Biosynthesis in Salvia miltiorrhiza through Endogenous Nitric Oxide and Jasmonic Acid. Molecules, 24(7), 1229. https://doi.org/10.3390/molecules24071229