
Polyphenols-Rich Fruit (Euterpe edulis Mart.) Prevents Peripheral Inflammatory Pathway Activation by the Short-Term High-Fat Diet

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
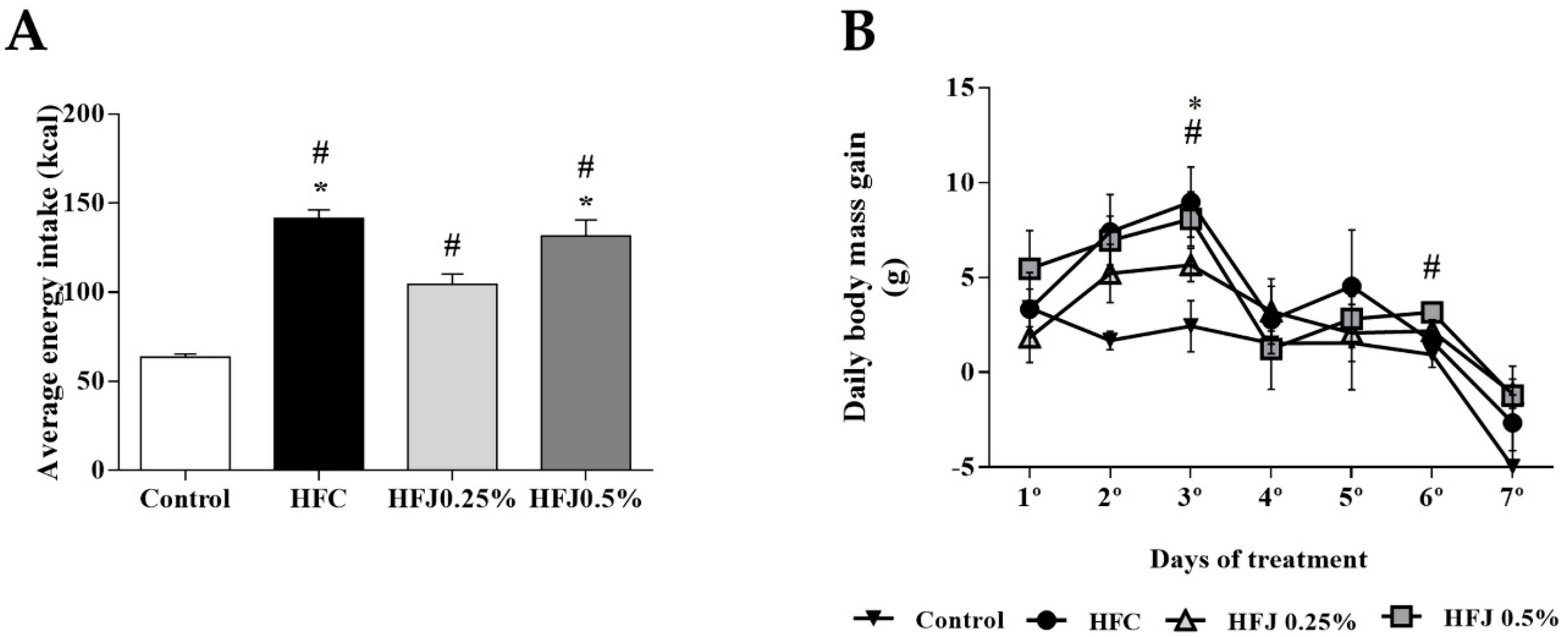
2.1. Diet Intake, Body Weight, and Tissue Weight
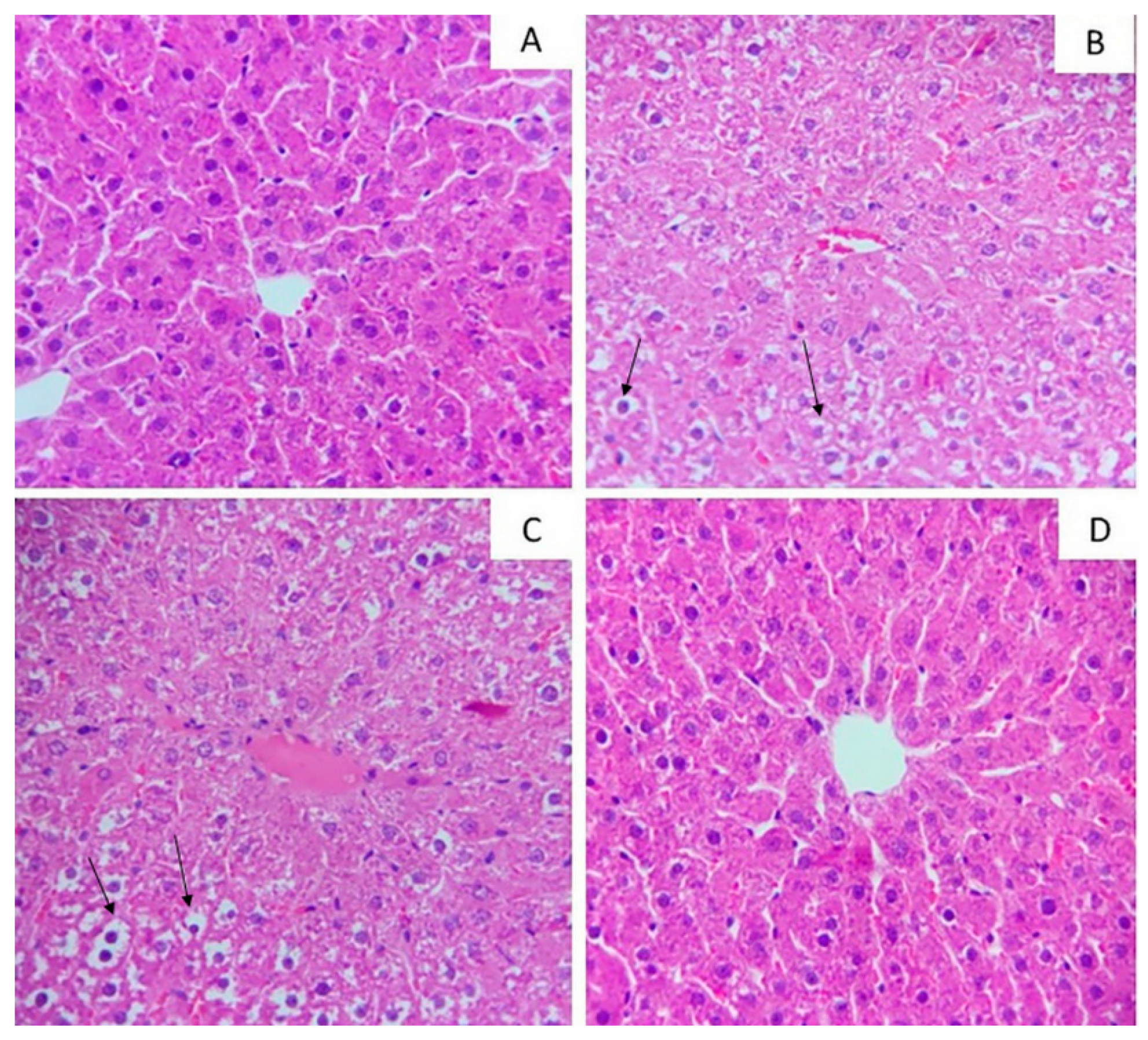
2.2. Hepatic Ectopic Fat Accumulation
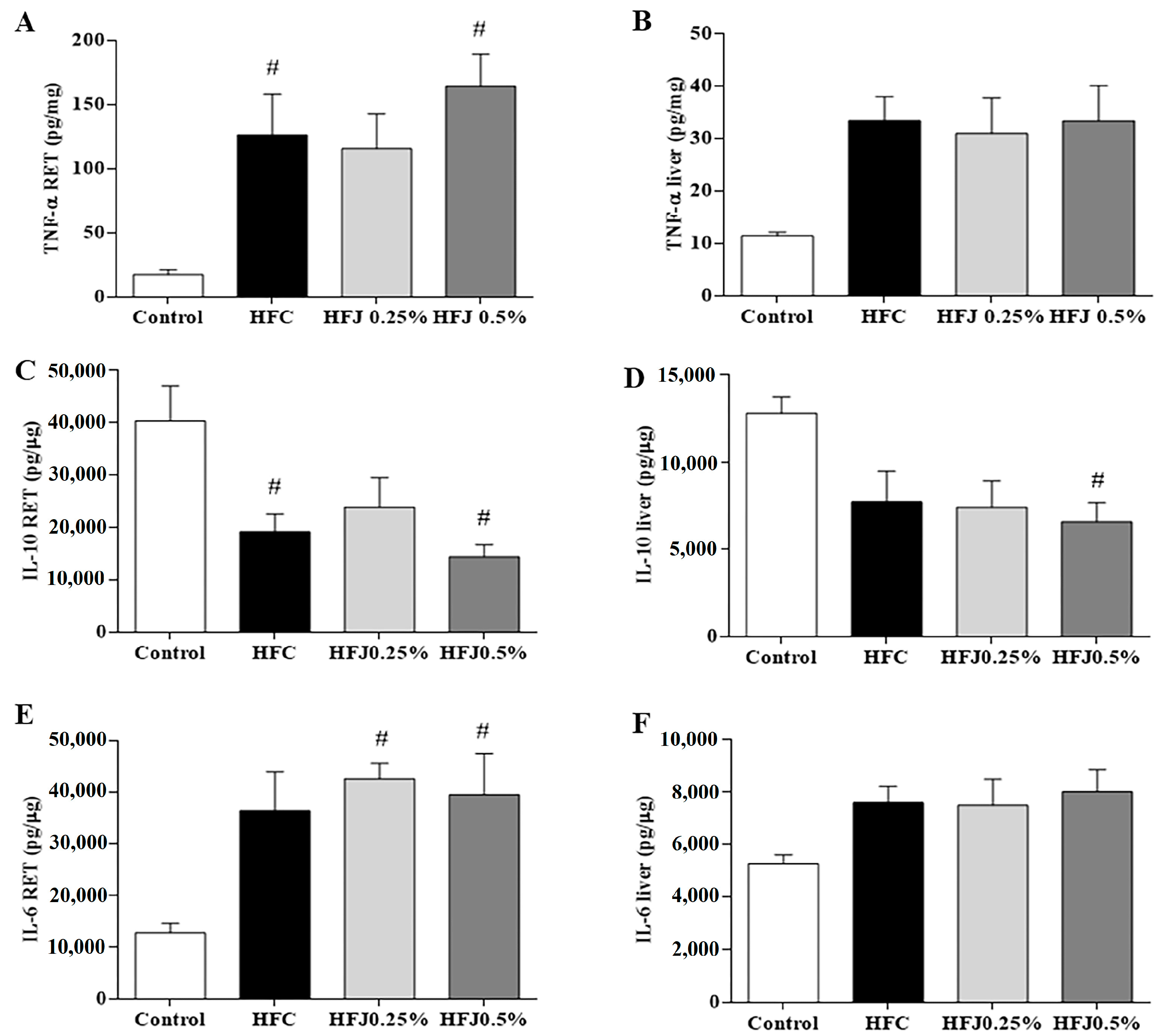
2.3. Cytokines Concentration
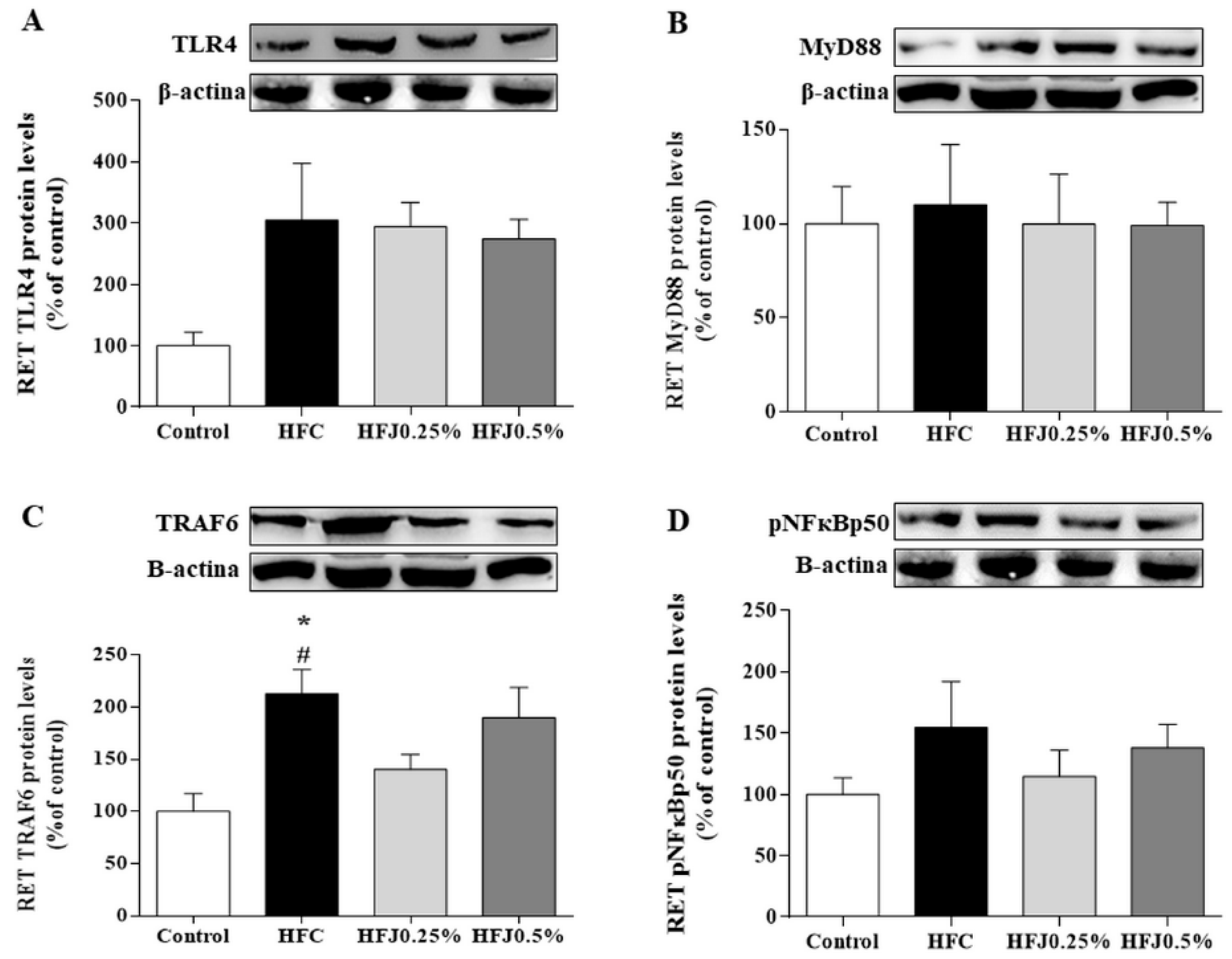
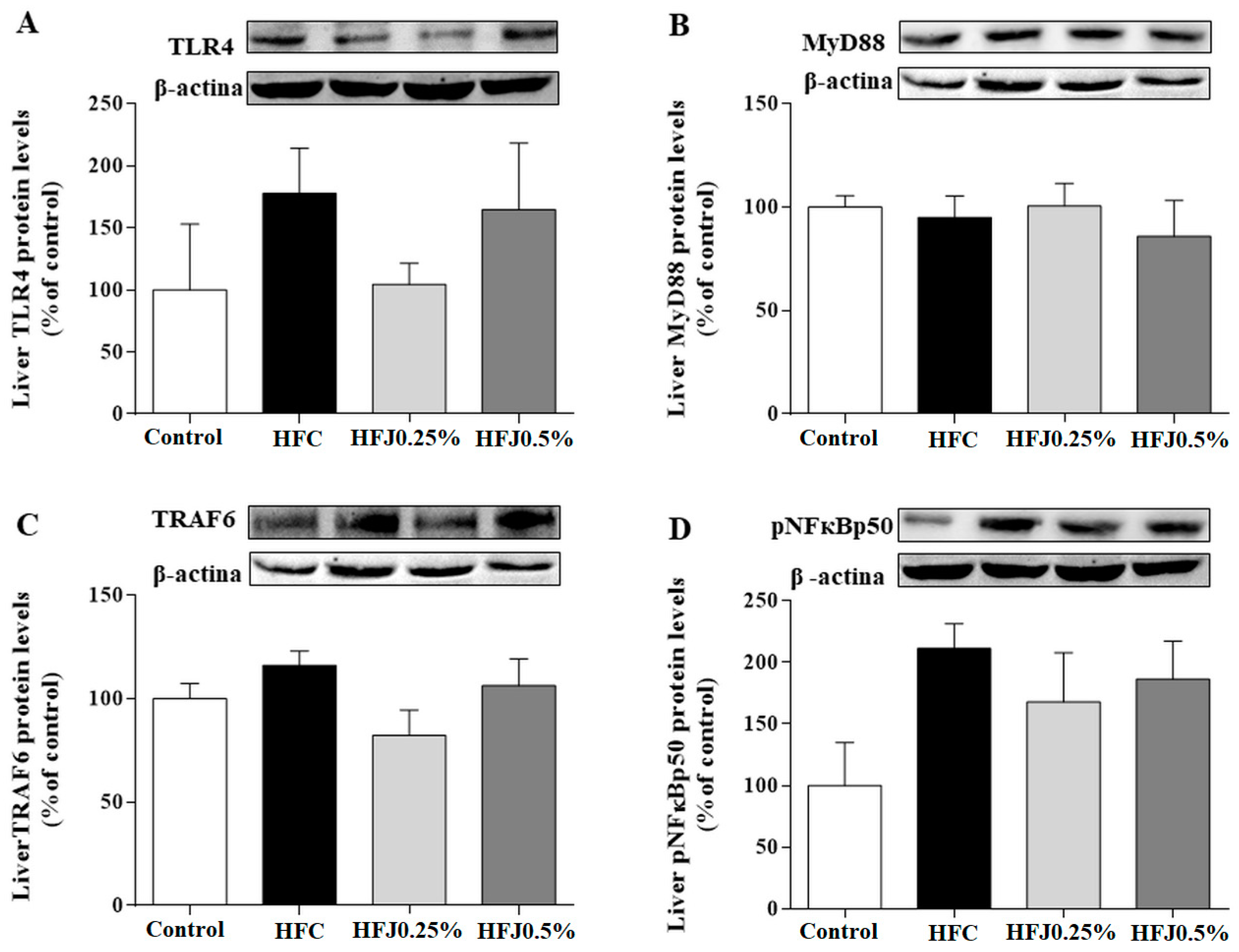
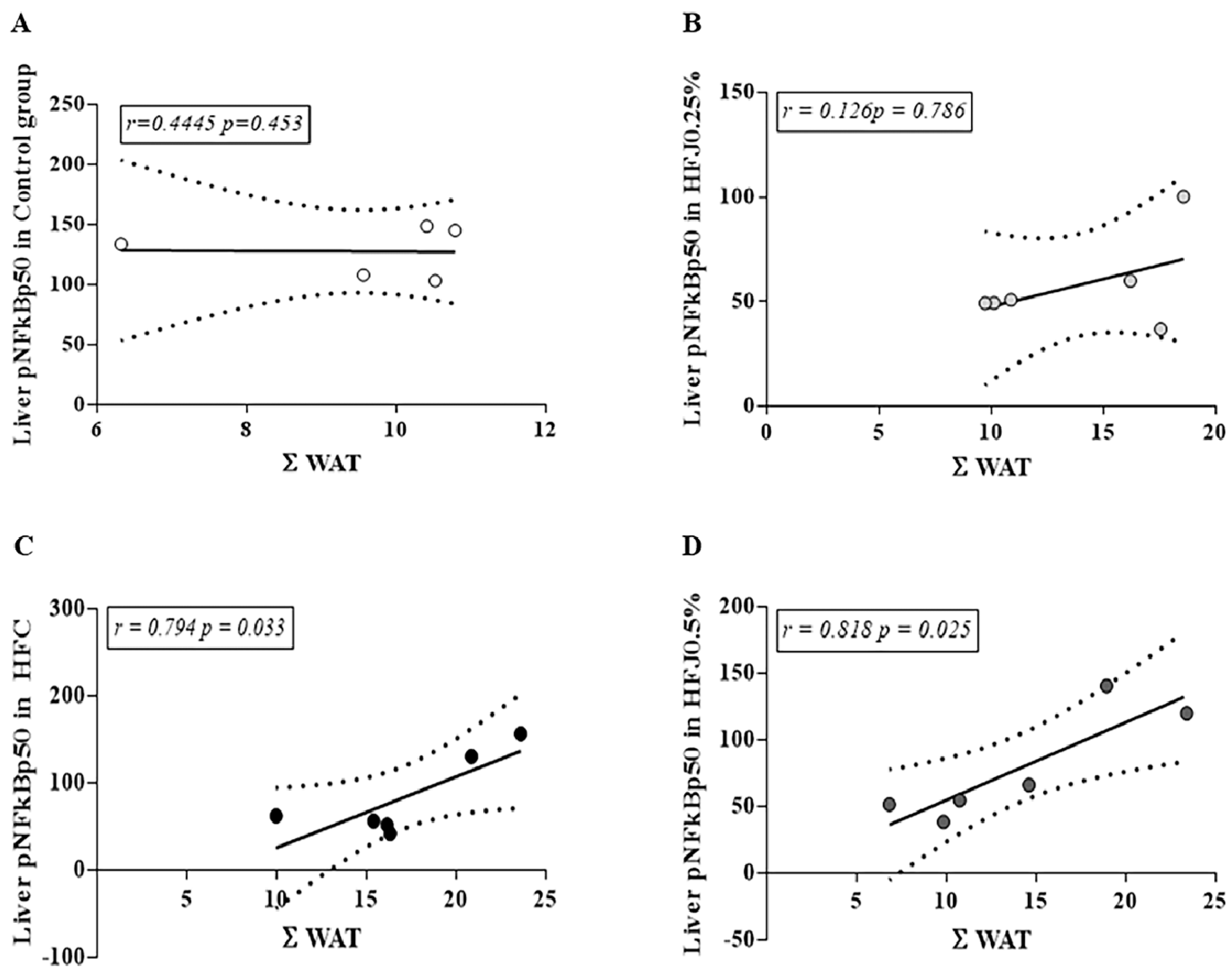
2.4. NFκB Pathway Protein Expression
3. Discussion
4. Materials and Methods
4.1. Short-Term High-Fat Diet
4.2. Freeze-Dried Juçara Pulp Powder
4.3. Animal Procedures
4.4. Histopathological Analysis
4.5. Hepatic Triacylglycerol Analysis.
4.6. Hepatic Enzymes in Serum
4.7. Tissue TNF-α, IL-6, and IL-10 Concentrations
4.8. Western Blotting Analyses
4.9. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Myles, I.A. Fast food fever: reviewing the impacts of the Western diet on immunity. Nutr. J. 2014, 13, 61. [Google Scholar] [CrossRef]
- Figueiredo, P.S.; Inada, A.C.; Marcelino, G.; Cardozo, C.M.L.; de Cássia Freitas, K.; de Cássia Avellaneda Guimarães, R.; de Castro, A.P.; do Nascimento, V.A.; Hiane, P.A. Fatty acids consumption: The role metabolic aspects involved in obesity and its associated disorders. Nutrients 2017, 9, 1158. [Google Scholar] [CrossRef]
- Shefer, G.; Marcus, Y.; Stern, N. Is obesity a brain disease? Neurosci. Biobehav. Rev. 2013, 37, 2489–2503. [Google Scholar] [CrossRef]
- Turner, N.; Kowalski, G.M.; Leslie, S.J.; Risis, S.; Yang, C.; Lee-Young, R.S.; Babb, J.R.; Meikle, P.J.; Lancaster, G.I.; Henstridge, D.C.; et al. Distinct patterns of tissue-specific lipid accumulation during the induction of insulin resistance in mice by high-fat feeding. Diabetologia 2013, 56, 1638–1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogero, M.M.; Calder, P.C. Obesity, inflammation, toll-like receptor 4 and fatty acids. Nutrients 2018, 10, 432. [Google Scholar] [CrossRef]
- Jialal, I.; Kaur, H.; Devaraj, S. Toll-like receptor status in obesity and metabolic syndrome: A translational perspective. J. Clin. Endocrinol. Metab. 2014, 99, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Schreckinger, M.E.; Lotton, J.; Lila, M.A.; de Mejia, E.G. Berries from South America: a comprehensive review on chemistry, health potential, and commercialization. J. Med. Food 2010, 13, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Silva, N.A.D.; Rodrigues, E.; Mercadante, A.Z.; De Rosso, V.V. Phenolic compounds and carotenoids from four fruits native from the Brazilian Atlantic forest. J. Agric. Food Chem. 2014, 62, 5072–5084. [Google Scholar] [CrossRef] [PubMed]
- Rufino, M.D.S.M.; Pérez-Jiménez, J.; Arranz, S.; Alves, R.E.; de Brito, E.S.; Oliveira, M.S.P.; Saura-Calixto, F. Açaí (Euterpe oleraceae) “BRS Pará”: A tropical fruit source of antioxidant dietary fiber and high antioxidant capacity oil. Food Res. Int. 2011, 44, 2100–2106. [Google Scholar] [CrossRef]
- Kang, J.; Thakali, K.M.; Xie, C.; Kondo, M.; Tong, Y.; Ou, B.; Jensen, G.; Medina, M.B.; Schauss, A.G.; Wu, X. Bioactivities of açaí (Euterpe precatoria Mart.) fruit pulp, superior antioxidant and anti-inflammatory properties to Euterpe oleracea Mart. Food Chem. 2012, 133, 671–677. [Google Scholar] [CrossRef]
- Andrich, D.E.; Melbouci, L.; Ou, Y.; Leduc-Gaudet, J.P.; Chabot, F.; Lalonde, F.; Lira, F.S.; Gaylinn, B.D.; Gouspillou, G.; Danialou, G.; Comtois, A.S.; St-Pierre, D.H. Altered Feeding Behaviors and Adiposity Precede Observable Weight Gain in Young Rats Submitted to a Short-Term High-Fat Diet. J. Nutr. Metab. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Jamar, G.; Santamarina, A.B.; Mennitti, L.V.; de Cássia Cesar, H.; Oyama, L.M.; de Rosso, V.V.; Pisani, L.P. Bifidobacterium spp. reshaping in the gut microbiota by low dose of juçara supplementation and hypothalamic insulin resistance in Wistar rats. J. Funct. Foods 2018, 46, 212–219. [Google Scholar] [CrossRef]
- Santamarina, A.B.; Jamar, G.; Mennitti, L.V.; de Rosso, V.V.; Cesar, H.C.; Oyama, L.M.; Pisani, L.P. The Use of Juçara (Euterpe edulis Mart.) Supplementation for Suppression of NF-κB Pathway in the Hypothalamus after High-Fat Diet in Wistar Rats. Mol. (Baselswitz.) 2018, 23, 1814. [Google Scholar] [CrossRef]
- Wiedemann, M.S.F.; Wueest, S.; Item, F.; Schoenle, E.J.; Konrad, D. Adipose tissue inflammation contributes to short-term high-fat diet-induced hepatic insulin resistance. Am. J. Physiol. -Endocrinol. Metab. 2013, 305, E388–E395. [Google Scholar] [CrossRef]
- Estadella, D.; da Penha Oller do Nascimento, C.M.; Oyama, L.M.; Ribeiro, E.B.; Dâmaso, A.R.; de Piano, A. Lipotoxicity: Effects of Dietary Saturated and Transfatty Acids. Mediat. Inflamm. 2013, 2013, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, N.; Sikder, K.; Bhattacharjee, S.; Majumdar, S.B.; Ghosh, S.; Majumdar, S.; Dey, S. Quercetin alleviates inflammation after short-term treatment in high-fat-fed mice. Food Funct. 2013, 4, 889. [Google Scholar] [CrossRef] [PubMed]
- Magri-Tomaz, L.; Melbouci, L.; Mercier, J.; Ou, Y.; Auclair, N.; Lira, F.S.; Lavoie, J.M.; St-Pierre, D.H. Two weeks of high-fat feeding disturb lipid and cholesterol molecular markers. Cell Biochem. Funct. 2018, 36, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Most, J.; Goossens, G.H.; Jocken, J.W.E.; Blaak, E.E. Short-term supplementation with a specific combination of dietary polyphenols increases energy expenditure and alters substrate metabolism in overweight subjects. Int. J. Obes. 2014, 38, 698–706. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, L.D.P.; Nogueira Neto, J.F.; Klein, M.R.S.T.; Sanjuliani, A.F. Short-term Effects of Green Tea on Blood Pressure, Endothelial Function, and Metabolic Profile in Obese Prehypertensive Women: A Crossover Randomized Clinical Trial. J. Am. Coll. Nutr. 2017, 36, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Lyall, K.A.; Hurst, S.M.; Cooney, J.; Jensen, D.; Lo, K.; Hurst, R.D.; Stevenson, L.M. Short-term blackcurrant extract consumption modulates exercise-induced oxidative stress and lipopolysaccharide-stimulated inflammatory responses. Am. J. Physiol. -Regul. Integr. Comp. Physiol. 2009, 297, R70–R81. [Google Scholar] [CrossRef] [PubMed]
- Mullan, A.; Delles, C.; Ferrell, W.; Mullen, W.; Edwards, C.A.; McColl, J.H.; Roberts, S.A.; Lean, M.E.; Sattar, N. Effects of a beverage rich in (poly)phenols on established and novel risk markers for vascular disease in medically uncomplicated overweight or obese subjects: A four week randomized trial. Atherosclerosis 2016, 246, 169–176. [Google Scholar] [CrossRef]
- Oyama, L.M.; Silva, F.P.; Carnier, J.; De Miranda, D.A.; Santamarina, A.B.; Ribeiro, E.B.; Oller Do Nascimento, C.M.; De Rosso, V.V. Jucąra pulp supplementation improves glucose tolerance in mice. Diabetol. Metab. Syndr. 2016, 8, 1–8. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, D.; Zhang, D.; Lv, Y.; Wei, Y.; Wu, W.; Zhou, F.; Tang, M.; Mao, T.; Li, M.; Ji, B. Inhibitory Effect of Blueberry Polyphenolic Compounds on Oleic Acid-Induced Hepatic Steatosis in Vitro. J. Agric. Food Chem. 2011, 59, 12254–12263. [Google Scholar] [CrossRef] [PubMed]
- Valenti, L.; Riso, P.; Mazzocchi, A.; Porrini, M.; Fargion, S.; Agostoni, C. Dietary anthocyanins as nutritional therapy for nonalcoholic fatty liver disease. Oxidative Med. Cell. Longev. 2013, 2013. [Google Scholar] [CrossRef]
- Lecci, R.; Logrieco, A.; Leone, A. Pro-Oxidative Action of Polyphenols as Action Mechanism for their Pro-Apoptotic Activity. Anti-Cancer Agents Med. Chem. 2014, 14, 1363–1375. [Google Scholar] [CrossRef]
- Hatia, S.; Septembre-Malaterre, A.; Le Sage, F.; Badiou-Bénéteau, A.; Baret, P.; Payet, B.; Lefebvre D’hellencourt, C.; Gonthier, M.P. Evaluation of antioxidant properties of major dietary polyphenols and their protective effect on 3T3-L1 preadipocytes and red blood cells exposed to oxidative stress. Free Radic. Res. 2014, 48, 387–401. [Google Scholar] [CrossRef] [PubMed]
- Blachnio-Zabielska, A.; Baranowski, M.; Zabielski, P.; Gorski, J. Effect of high fat diet enriched with unsaturated and diet rich in saturated fatty acids on sphingolipid metabolism in rat skeletal muscle. J. Cell. Physiol. 2010, 225, 786–791. [Google Scholar] [CrossRef] [PubMed]
- Buettner, R.; Parhofer, K.G.; Woenckhaus, M.; Wrede, C.E.; Kunz-Schughart, L.A.; Schölmerich, J.; Bollheimer, L.C. Defining high-fat-diet rat models: Metabolic and molecular effects of different fat types. J. Mol. Endocrinol. 2006, 36, 485–501. [Google Scholar] [CrossRef] [PubMed]
- Gerner, R.R.; Wieser, V.; Moschen, A.R.; Tilg, H. Metabolic inflammation: role of cytokines in the crosstalk between adipose tissue and liver. Can. J. Physiol. Pharm. 2013, 91, 867–872. [Google Scholar] [CrossRef]
- Lira, F.S.; Rosa, J.C.; Dos Santos, R.V.; Venancio, D.P.; Carnier, J.; Sanches, P.D.L.; Do Nascimento, C.M.O.; De Piano, A.; Tock, L.; Tufik, S.; et al. Visceral fat decreased by long-term interdisciplinary lifestyle therapy correlated positively with interleukin-6 and tumor necrosis factor-α and negatively with adiponectin levels in obese adolescents. Metab. Clin. Exp. 2011, 60, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Sánchez, A.; Madrigal-Santillán, E.; Bautista, M.; Esquivel-Soto, J.; Morales-González, Á.; Esquivel-Chirino, C.; Durante-Montiel, I.; Sánchez-Rivera, G.; Valadez-Vega, C.; Morales-González, J.A. Inflammation, oxidative stress, and obesity. Int. J. Mol. Sci. 2011, 12, 3117–3132. [Google Scholar] [CrossRef] [PubMed]
- Jayarathne, S.; Koboziev, I.; Park, O.H.; Oldewage-Theron, W.; Shen, C.L.; Moustaid-Moussa, N. Anti-Inflammatory and Anti-Obesity Properties of Food Bioactive Components: Effects on Adipose Tissue. Prev. Nutr. Food Sci. 2017, 22, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Dornellas, A.P.S.; Watanabe, R.L.H.; Pimentel, G.D.; Boldarine, V.T.; Nascimento, C.M.O.; Oyama, L.M.; Ghebremeskel, K.; Wang, Y.; Bueno, A.A.; Ribeiro, E.B. Deleterious effects of lard-enriched diet on tissues fatty acids composition and hypothalamic insulin actions. Prostaglandins Leukot. Essent. Fat. Acids 2015, 102–103, 21–29. [Google Scholar] [CrossRef]
- Karlsen, A.; Retterstøl, L.; Laake, P.; Paur, I.; Kjølsrud-Bøhn, S.; Sandvik, L.; Blomhoff, R. Anthocyanins Inhibit Nuclear Factor-κB Activation in Monocytes and Reduce Plasma Concentrations of Pro-Inflammatory Mediators in Healthy Adults. J. Nutr. 2007, 137, 1951–1954. [Google Scholar] [CrossRef] [Green Version]
- Graf, D.; Seifert, S.; Jaudszus, A.; Bub, A.; Watzl, B. Anthocyanin-Rich Juice Lowers Serum Cholesterol, Leptin, and Resistin and Improves Plasma Fatty Acid Composition in Fischer Rats. PLoS ONE 2013, 8, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Center for Drug Evaluation and Research Guidance for Industry: Estimating the Maximum Safe Starting Dose in Initial Clinical Trials for Therapeutics in Adult Healthy Volunteers. Us Dep. Health Hum. Serv. 2005, 1–27. [CrossRef]
- Da Silva, V.H.P.; de Moura, C.F.G.; Ribeiro, F.A.P.; Cesar, A.; Pereira, C.D.S.; Silva, M.J.D.; Vilegas, W.; Ribeiro, D.A. Genotoxicity and cytotoxicity induced by municipal effluent in multiple organs of Wistar rats. Environ. Sci. Pollut. Res. 2014, 21, 13069–13080. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1953, 226, 497–509. [Google Scholar] [CrossRef]
Sample Availability: Not available. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Absolute tissue weight (g) | |||||
|---|---|---|---|---|---|
| Control (n = 6) | HFC (n = 7) | HFJ0.25% (n = 7) | HFJ0.5% (n = 7) | p-Vaule | |
| Liver | 10.57 ± 0.54 | 11.46 ± 0.66 | 10.23 ± 0.70 | 10.16 ± 0.85 | - |
| RET | 3.39 ± 0.52 | 6.99 ± 0.90 # | 4.67 ± 0.71 | 4.95 ± 0.64 | 0.029 |
| EPI | 3.74 ± 0.38 | 5.67 ± 0.69 # | 4.20 ± 0.61 | 3.94 ± 0.70 | 0.044 |
| MES | 3.57 ± 0.64 | 4.18 ± 0.44 | 3.44 ± 0.26 | 3.99 ± 0.50 | - |
| ΣWAT | 10.70 ± 1.36 | 16.84 ± 1.64 # | 12.32 ± 1.37 | 12.88 ± 1.76 | 0.020 |
| 0 | 1 | 2 | 3 | |
|---|---|---|---|---|
| Control (n = 5) | 5 | 0 | 0 | 0 |
| HFC (n = 7) | 4 | 3 | 0 | 0 |
| HFJ0.5% (n = 7) | 3 | 4 | 0 | 0 |
| HFJ0.25% (n = 7) | 5 | 2 | 0 | 0 |
| Control (n = 6) | HFC (n = 7) | HFJ0.25% (n = 7) | HFJ0.5% (n = 7) | |
|---|---|---|---|---|
| AST (U/L) | 36.18 ± 2.77 | 49.43 ± 4.99 & | 34.11 ± 7.25 | 49.79 ± 6.31 & |
| ALT (U/L) | 16.61 ± 1.37 | 23.06 ± 9.89 | 15.41 ± 3.21 | 17.17 ± 4.58 |
| TAG (mg/100mg) | 129.91 ± 4.41 | 157.33 ± 4.92 # | 142.59 ± 5.21 | 151.71 ± 2.14 # |
| Components | Diet (g/100g) | |||
|---|---|---|---|---|
| Control | HFC | HFJ0.25% | HFJ0.5% | |
| Standard chow * | 100 | 50 | 50 | 50 |
| Sucrose | - | 10 | 10 | 10 |
| Casein | - | 20 | 20 | 20 |
| Soybean oil | - | 2 | 2 | 2 |
| Lard | 18 | 18 | 18 | |
| Butyl hydroquinone | - | 0.004 | 0.004 | 0.004 |
| Juçara pulp powder | - | - | 0.25 | 0.5 |
| Mineral mix® § | 0.5 | 0.5 | 0.5 | |
| Vitamins mix® & | 1.75 | 1.75 | 1.75 | |
| Energy (Kcal/100g) | 270 | 410 | 420 | 430 |
| Bioactive Compound | Concentration (in 100 g Dry Basis) |
| Cyanidin 3-rutinoside (mg) | 1790.0 ± 57.5 |
| Cyanidin 3-glucoside (mg) | 740.9 ± 22.1 |
| Total anthocyanins (mg) | 2663.7 ± 76.2 |
| Apigenin deoxyhexosyl-hexoside (mg) | 224.7 ± 13.2 |
| Luteolin deoxyhexosyl-hexoside (mg) | 332.7 ± 16.8 |
| Dihydrokaempferol-hexoside (mg) | 587.6 ± 23.0 |
| Total phenolic compounds (mg) | 3976.1 ± 197.34 |
| Total fiber (g) | 28.3 ± 0.3 |
| Fatty Acids | (% Fatty Acids Total) |
| Oleic acid (C18:1) | 44.2 |
| Linoleic acid (C18:2) | 17.4 |
| Linolenic acid (C18:3) | 0.45 |
| Score | Necrosis Area | Alterations |
|---|---|---|
| 0 | 0% | Preserved structures |
| 1 | <30% | Discreet eosinophilia, the presence of intracytoplasmic vacuoles |
| 2 | ≥30% | Marked eosinophilia, dilation of sinusoid space, congested vessels, and intracytoplasmic vacuoles |
| 3 | ≥50% | Marked eosinophilia, congested vessels, intracytoplasmic vacuoles, karyolysis, and structural loss |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santamarina, A.B.; Jamar, G.; Mennitti, L.V.; Ribeiro, D.A.; Cardoso, C.M.; de Rosso, V.V.; Oyama, L.M.; Pisani, L.P. Polyphenols-Rich Fruit (Euterpe edulis Mart.) Prevents Peripheral Inflammatory Pathway Activation by the Short-Term High-Fat Diet. Molecules 2019, 24, 1655. https://doi.org/10.3390/molecules24091655
Santamarina AB, Jamar G, Mennitti LV, Ribeiro DA, Cardoso CM, de Rosso VV, Oyama LM, Pisani LP. Polyphenols-Rich Fruit (Euterpe edulis Mart.) Prevents Peripheral Inflammatory Pathway Activation by the Short-Term High-Fat Diet. Molecules. 2019; 24(9):1655. https://doi.org/10.3390/molecules24091655
Chicago/Turabian StyleSantamarina, Aline Boveto, Giovana Jamar, Laís Vales Mennitti, Daniel Araki Ribeiro, Caroline Margonato Cardoso, Veridiana Vera de Rosso, Lila Missae Oyama, and Luciana Pellegrini Pisani. 2019. "Polyphenols-Rich Fruit (Euterpe edulis Mart.) Prevents Peripheral Inflammatory Pathway Activation by the Short-Term High-Fat Diet" Molecules 24, no. 9: 1655. https://doi.org/10.3390/molecules24091655
APA StyleSantamarina, A. B., Jamar, G., Mennitti, L. V., Ribeiro, D. A., Cardoso, C. M., de Rosso, V. V., Oyama, L. M., & Pisani, L. P. (2019). Polyphenols-Rich Fruit (Euterpe edulis Mart.) Prevents Peripheral Inflammatory Pathway Activation by the Short-Term High-Fat Diet. Molecules, 24(9), 1655. https://doi.org/10.3390/molecules24091655