
Medium-Chain Fatty Acids from Eugenia winzerlingii Leaves Causing Insect Settling Deterrent, Nematicidal, and Phytotoxic Effects


 ,
, 
Abstract
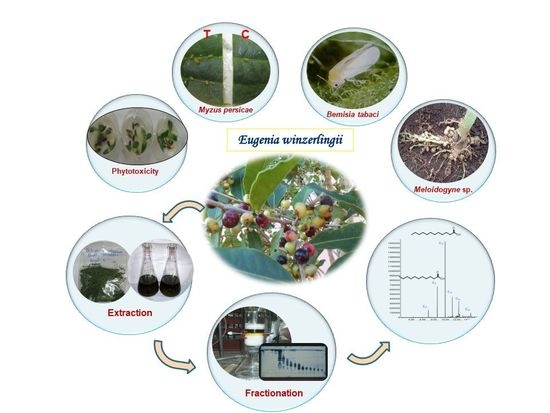
:
1. Introduction
2. Results
2.1. Effect of Extracts and their Fractions on Insect Settling Deterrence and Nematode Mortality
2.2. Major Constituents of Active Fractions
2.3. Biological Activity of Fatty Acids and Derivatives
2.4. Phytotoxicity of Fatty Acids
3. Discussion
4. Materials and Methods
4.1. Plant Material and Extraction
4.2. Chromatographic Fractionation of Organic Extracts
4.3. Preparation of Methyl and Ethyl Esters
4.4. Analytical Method
4.5. Bioassays
4.5.1. Evaluation of settling inhibition on Myzus persicae
4.5.2. Evaluation of oviposition inhibition on Bemisia tabaci
4.5.3. Nematicidal Assay
4.5.4. Phytotoxic Assay
4.6. Data Analysis
5. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bebber, D.P.; Holmes, T.; Gurr, S.J. The global spread of crop pests and pathogens. Glob. Ecol. Biogeogr. 2014, 23, 1398–1407. [Google Scholar] [CrossRef] [Green Version]
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Blackman, R.L.; Eastop, V.F. Aphids on the World’s Crops: An Identification and Information Guide, 2nd ed.; Wiley and Sons: Chichester, UK, 2000; ISBN 0471851914. [Google Scholar]
- Oliveira, M.R.V.; Henneberry, T.J.; Anderson, P. History, current status, and collaborative research projects for Bemisia tabaci. Crop Prot. 2001, 20, 709–723. [Google Scholar] [CrossRef]
- Sasser, J.N.; Eisenback, J.D.; Carter, C.C.; Triantaphyllou, A.C. The international Meloidogyne project-Its goals and accomplishments. Annu. Rev. Phytopathol. 1983, 21, 271–288. [Google Scholar] [CrossRef]
- Aktar, M.W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdisc. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef]
- Bass, C.; Denholm, I.; Williamson, M.S.; Nauen, R. The global status of insect resistance to neonicotinoid insecticides. Pest Biochem. Physiol. 2015, 121, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, A.R.; Ellsworth, P.C.; Ishaaya, I. Biorational pest control—An overview. In Biorational Control of Arthropod Pests. Application and Resistance Management, 1st ed.; Ishaaya, I., Horowitz, A.R., Eds.; Springer: Dordrecht, The Netherlands, 2009; Volume 1, pp. 1–20. [Google Scholar]
- Copping, L.G.; Duke, S.O. Natural products that have been used commercially as crop protection agents. Pest Manag. Sci. 2007, 63, 524–554. [Google Scholar] [CrossRef]
- Dubey, N.K. Natural Products in Plant Pest Management, 1st ed.; CABI: Oxfordshire, UK, 2011; pp. 1–312. ISBN 978-1-84593-671-6. [Google Scholar]
- Ntalli, N.G.; Caboni, P. Botanical nematicides: A review. J. Agric. Food Chem. 2012, 60, 9929–9940. [Google Scholar] [CrossRef] [PubMed]
- González-Coloma, A.; Reina, M.; Díaz, C.E.; Fraga, B.M. Natural product-based biopesticides for insect control. In Comprehensive Natural Products Chemistry II: Chemistry and Biology; Mander, L., Liu, H.W., Eds.; Elsevier Ltd.: Oxfor, UK, 2010; Volume 3, pp. 237–268. ISBN 978-0-08-045382-8. [Google Scholar]
- Nawrot, J.; Harmatha, J. Phytochemical feeding deterrents for stored product insect pests. Phytochem. Rev. 2012, 11, 543–566. [Google Scholar] [CrossRef]
- Ntalli, N.G.; Menkissoglu-Spiroudi, U. Pesticides of botanical origin: A promising tool in plant protection. In Pesticides—Formulations, Effects, Fate; Stoytcheva, M., Ed.; InTech Europe: Rijeka, Croatia, 2011; Volume 1, pp. 3–24. ISBN 978-953-307-532-7. [Google Scholar]
- Asolkar, R.N.; Cordova-Kreylos, A.N.; Himmel, P.; Marrone, P.G. Discovery and development of natural products for pest management. In Pest Management with Natural Products; Beck, J.J., Coats, J.R., Duke, S.O., Koivunen, M.E., Eds.; American Chemical Society: Washington, DC, USA, 2013; pp. 17–30. [Google Scholar] [CrossRef]
- De Souza, A.M.; de Oliveira, C.F.; de Oliveira, V.B.; Betim, C.M.; Miguel, O.G.; Miguel, M.D. Traditional uses, phytochemistry, and antimicrobial activities of Eugenia species—A review. Planta Med. 2018, 84, 1232–1248. [Google Scholar] [CrossRef]
- De Souza, M.A.D.; Scudeller, V.V.; de Medonca, M.S. Three new species of Eugenia (Myrtaceae) from Brazilian Amazonia. Phytotaxa 2015, 212, 087–094. [Google Scholar] [CrossRef]
- Villaseñor, J.L. Checklist of the native vascular plants of Mexico. Rev. Mex. Biodivers. 2016, 87, 550–902. [Google Scholar] [CrossRef]
- Cruz-Estrada, A.; Ruiz-Sánchez, E.; Medina-Baizabal, I.L.; Balam-Uc, E.; Gamboa-Angulo, M. Effect of Eugenia winzerlingii extracts on Bemisia tabaci and evaluation of its nursery propagation. Phyton. J. Exp. Bot. 2019, in press. [Google Scholar]
- Cristóbal-Alejo, J.; Tun-Suárez, J.M.; Moguel-Catzin, S.; Marbán-Mendoza, N.; Medina-Baizabal, L.; Simá-Polanco, P.; Peraza-Sánchez, E.R.; Gamboa-Angulo, M.M. In vitro sensitivity of Meloidogyne incognita to extracts from native Yucatecan plants. Nematropica 2006, 36, 89–97. [Google Scholar]
- Carnevali Fernández-Concha, G.; Tapia-Muñoz, J.L.; Duno de Stefano, R.; Ramírez Morillo, I.M. Flora ilustrada de la Península de Yucatán: Listado florístico, 1st ed.; Carnevali Fernández-Concha, G., Tapia-Muñoz, J.L., Duno de Stefano, R., Ramírez Morillo, I.M., Eds.; CICY: Yucatán, Mexico, 2010; p. 326. ISBN 9686077823070. [Google Scholar]
- Gupta, A.; Koolwal, N.; Dobhal, M.P.; Sharma, M.C. Biological importance of phytochemical constituents isolated from Eugenia. J. Indian Chem. Soc. 2014, 91, 1539–1553. [Google Scholar]
- Magina, M.D.A.; Dalmarco, E.M.; Wisniewski, A., Jr.; Simionatto, E.L.; Dalmarco, J.B.; Pizzolattti, M.G.; Brighente, I.M.C. Chemical composition and antibacterial activity of essential oils of Eugenia species. J. Nat. Med. 2009, 63, 345–350. [Google Scholar] [CrossRef]
- Gayoso, C.W.; Lima, E.O.; Oliveira, V.T.; Pereira, F.O.; Souza, E.L.; Lima, I.O. Navarro Sensitivity of fungi isolated from onychomycosis to Eugenia cariophyllata essential oil and D.F. Fitoterapia 2005, 76, 247–249. [Google Scholar] [CrossRef] [PubMed]
- Frausin, G.; Hidalgo, A.F.; Souza-Lima, R.B. An ethnobotanical study of anti-malarial plants among indigenous people on the upper Negro River in the Brazilian Amazon. J. Ethnopharmacol. 2015, 174, 238–252. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Bowling, J.J.; Smithson, D.; Clark, J.; Jacob, M.R.; Khan, S.I.; Tekwani, B.L.; Connelly, M.; Samoylenko, V.; Ibrahim, M.A.; et al. Diversity-oriented natural product platform identifies plant constituents targeting Plasmodium falciparum. Malar. J. 2016, 15, 270–285. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, L.; Tacoronte, J.E.; Navarro, A.; Leyva, M.; Bello, A.; Cabrera, M.T.; Marquetti, M.C. Composición química y actividad biológica del aceite esencial de Eugenia melanadenia (Myrtales: Myrtaceae) sobre Blattella germanica (Dictyoptera: Blattellidae). Rev. CENIC Ciencias Quím. 2004, 35, 131–134. [Google Scholar]
- Chaieb, K.; Hajlaoui, H.; Zmantar, T.; Kahla-Nakbi, A.B.; Rouabhia, M.; Mahdouani, K.; Bakhrouf, A. The chemical composition and biological activity of clove essential oil, Eugenia caryophyllata (Syzigium aromaticum L. Myrtaceae): A Short Review. Phytother. Res. 2007, 21, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.C.; Lee, S.H.; Lee, W.J.; Choi, D.H.; Ahn, Y.J. Ovicidal and adulticidal effects of Eugenia caryophyllata bud and leaf oil compounds on Pediculus capitis. J. Agric. Food Chem. 2003, 51, 4884–4888. [Google Scholar] [CrossRef]
- Trongtokit, Y.; Rongsriyam, Y.; Komalamisra, N.; Apiwathnasorn, C. Comparative repellency of 38 essential oils against mosquito bites. Phytother. Res. 2005, 19, 303–309. [Google Scholar] [CrossRef]
- Ho, S.H.; Cheng, L.P.L.; Sim, K.Y.; Tan, H.T.W. Potential of cloves (Syzygium aromaticum L.) Merr. and Perry as a grain protectant against Tribolium castaneum (Herbst) and Sitophilus zeamais Motsch. Postharv. Biol. Technol. 1994, 4, 179–183. [Google Scholar] [CrossRef]
- Jairoce, C.F.; Teixeira, C.M.; Nunes, C.F.P.; Nunes, A.M.; Pereira, C.M.P.; Garcia, F.R.M. Insecticide activity of clove essential oil on bean weevil and maize weevil. Rev. Bras. Eng. Agric. Ambient. 2016, 20, 72–77. [Google Scholar] [CrossRef] [Green Version]
- Choi, W.-I.; Lee, E.-H.; Choi, B.-R.; Park, H.-M.; Ahn, Y.-J. Toxicity of plant essential oils to Trialeurodes vaporariorum (Homoptera: Aleyrodidae). J. Econ. Entomol. 2003, 96, 1479–1484. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Lee, S.G.; Lee, H.S. Active component isolated from Eugenia caryophyllata leaves and its structural analogues show insecticidal properties against Pochazia shantungensis. Appl. Biol. Chem. 2016, 59, 609–614. [Google Scholar] [CrossRef]
- Ibrahim, R.A.; Alahmadi, S.S. Effect of Syzygium aromaticum cloves on larvae of the rhinoceros beetle, Oryctes agamemnon (Coleoptera: Scarabaeidae). Afr. Entomol. 2015, 23, 458–466. [Google Scholar] [CrossRef]
- Andrés, M.F.; González-Coloma, A.; Sanz, J.; Burillo, J.; Sainz, P. Nematicidal activity of essential oils: A review. Phytochem. Rev. 2012, 11, 371–390. [Google Scholar] [CrossRef]
- Lobo, A.P.; da Camara, C.A.G.; de Melo, J.P.R.; de Moraes, M.M. Chemical composition and repellent activity of essential oils from the leaves of Cinnamomum zeylanicum and Eugenia uniflora against Diaphania hyalinata L. (Lepidoptera: Crambidae). J. Plant Dis. Protect. 2019, 126, 79–87. [Google Scholar] [CrossRef]
- Santana, O.; Reina, M.; Fraga, B.M.; Sanz, J.; González-Coloma, A. Antifeedant activity of fatty acid esters and phytosterols from Echium wildpretii. Chem. Biodiver. 2012, 9, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Auricchio, M.T.; Bacchi, E.M. Eugenia uniflora L. “brazilian cherry” leaves: Pharmacobotanical, chemical and pharmacological properties. Rev. Inst. Adolfo Lutz 2003, 62, 55–61. [Google Scholar]
- Mesquita, P.R.R.; Nunes, E.C.; dos Santos, F.N.; Bastos, L.P.; Costa, M.A.P.C.; Rodrigues, F.M.; de Andrade, J.B. Discrimination of Eugenia uniflora L. biotypes based on volatile compounds in leaves using HS-SPME/GC–MS and chemometric analysis. Microchem. J. 2017, 130, 79–87. [Google Scholar] [CrossRef]
- Zatelli, G.A.; Zimath, P.; Tenfen, A.; de Cordova, C.M.M.; Scharf, D.R.; Simionatto, E.L.; Alberton, M.D.; Falkenberg, M. Antimycoplasmic activity and seasonal variation of essential oil of Eugenia hiemalis Cambess. (Myrtaceae). Nat. Prod. Res. 2016, 30, 1961–1964. [Google Scholar] [CrossRef]
- Beaudoin, F.; Sayanova, O.; Haslam, R.P.; Bancroft, I.; Napier, J.A. Oleaginous crops as integrated production platforms for food, feed, fuel and renewable industrial feedstack-Manipulation of plant lipid composition via metabolic engineering and new opportunities from association genetics for crop improvement and valorization of co-products. Ocl 2014, 21, D606. [Google Scholar] [CrossRef]
- Müller, C.; Riederer, M. Plant surface properties in chemical ecology. J. Chem. Ecol. 2005, 31, 2621–2651. [Google Scholar] [CrossRef] [PubMed]
- Castillo, L.; Díaz, M.; González-Coloma, A.; González, A.; Alonso-Paz, E.; Bassagoda, M.J.; Rossini, C. Clytostoma callistegioides (Bignoniaceae) wax extract with activity on aphid settling. Phytochemistry 2010, 71, 2052–2057. [Google Scholar] [CrossRef] [PubMed]
- Phelan, P.L.; Miller, J.R. Post-landing behaviour of alate Myzus persicae as altered by (E)-β-farnesene and three carboxylic acids. Ent. Exp. Appl. 1982, 32, 46–53. [Google Scholar] [CrossRef]
- Ali, A.; Cantrell, C.L.; Bernier, U.R.; Duke, O.S.; Schneider, J.C.; Agramonte, N.M.; Khan, I. Aedes aegypti (Diptera: Culicidae) biting deterrence: Structure-activity relationship of saturated and unsaturated fatty acids. J. Med. Entomol. 2012, 49, 1370–1378. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.M.P.; Klun, J.A.; Cantrell, C.L.; Ragone, D.; Chauhan, K.R.; Brown, P.N.; Murch, S.J. Isolation and identification of mosquito (Aedes aegypti) biting deterrent fatty acids from male inflorescences of breadfruit (Artocarpus altilis (Parkinson) Fosberg). J. Agric. Food Chem. 2012, 60, 3867–3873. [Google Scholar] [CrossRef]
- Seenivasagan, T.; Guha, L.; Iqbal, S.T. Behavioral and electrophysiological responses of Culex quinquefasciatus to certain fatty acid esters. Acta Trop. 2013, 128, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Li, G.Q. Olfactory perception of oviposition-deterring fatty acids and their methyl esters by the asian corn borer, Ostrinia furnacalis. J. Insect Sci. 2009, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Davis, E.L.; Meyers, D.M.; Dullum, C.J.; Feitelson, J.S. Nematicidal activity of fatty acid esters on soybean cyst and root-knot nematode. J. Nematol. 1997, 29, 677–684. [Google Scholar] [PubMed]
- Zhang, W.P.; Ruan, W.B.; Deng, Y.Y.; Gao, Y.B. Potential antagonistic effects of nine natural fatty acids against Meloidogyne incognita. J. Agric. Food Chem. 2012, 60, 11631–11637. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Li, X.; Huang, L.; Gao, Y.; Zhong, L.; Zheng, Y.; Zuo, Y. Lauric acid in crown daisy root exudate potently regulates root-knot nematode chemotaxis and disrupts MI-flp-18 expression to block infection. J Exp. Bot. 2014, 65, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Wafula, G.O.; Muthomi, J.W.; Nderitu, J.H.; Chemining’wa, G.N. Efficacy of potassium salts of fatty acids in the management of thrips and whitefly on snap beans. Sustain. Agric. Res. 2017, 6, 45–54. [Google Scholar] [CrossRef]
- Mohamad, S.F.S.; Mohamad, S.; Aziz, A.A. The susceptibility of aphids, Aphis gossypii Glover to lauric acid based natural pesticide. Procedia Eng. 2013, 53, 20–28. [Google Scholar] [CrossRef]
- Gallardo, M.; de Rueda, P.M.; Matilla, A.J.; Sanches-Calle, I.M. Effect of short-chain fatty acids on the ethylene pathway in embryonic axes of Cicer arietinum during germination. Physiol. Plant 1994, 92, 629–635. [Google Scholar] [CrossRef]
- Himanen, M.; Prochazka, P.; Hänninen, K.; Oikari, A. Phytotoxicity of low-weight carboxylic acids. Chemosphere 2012, 88, 426–431. [Google Scholar] [CrossRef]
- Babiano, J.; Nicolas, G.; Matilla, A. Changes in the cellular permeability of the embryonic axis in Cicer arietinum L. seeds during germination. Biol. Plant. 1989, 31, 92–99. [Google Scholar] [CrossRef]
- Ferrarese, M.L.L.; Baleroni, C.R.S.; Ferrarese-Filho, O. Effects of fatty acids on carbohydrates and lipids of canola seeds during germination. Braz. Arch. Biol. Technol. 1998, 41, 315–319. [Google Scholar] [CrossRef]
- Coll, J.C.; Bowden, B.F. The application of vacuum liquid chromatography to the separation of the terpene mixtures. J. Nat. Prod. 1986, 49, 934–936. [Google Scholar] [CrossRef]
- Abdelillah, A.; Houcine, B.; Halima, D.; Meriem, C.S.; Imane, Z.; Eddine, S.D.; Abdallah, M.; Daoudi, C.S. Evaluation of antifungal activity of free fatty acids methyl esters fraction isolated from Algerian Linum usitatissimum L. seeds against toxigenic Aspergillus. Asian Pac. J. Trop. Biomed. 2013, 3, 443–448. [Google Scholar] [CrossRef]
- Hori, M. Antifeeding, settling inhibitory and toxic activities of labiate essential oils against the green peach aphid, Myzus persicae (Sulzer) (Homoptera: Aphididae). Appl. Entomol. Zoo. 1999, 34, 113–118. [Google Scholar] [CrossRef]
- Cruz-Estrada, A.; Gamboa-Angulo, M.; Borges-Argaez, R.; Ruiz-Sanchez, E. Insecticidal effects of plant extracts on immature whitefly Bemisia tabaci Genn. (Hemiptera: Aleyroideae). Electron. J. Biotech. 2013, 16, 1–6. [Google Scholar] [CrossRef]
- Baldin, E.L.L.; Crotti, A.E.M.; Wakabayashi, K.A.L.; Silva, J.P.G.F.; Aguiar, G.P.; Souza, E.S.; Veneziani, R.C.S.; Groppo, M. Plant-derived essential oils affecting settlement and oviposition of Bemisia tabaci (Genn.) biotype B on tomato. J. Pest. Sci. 2013, 86, 301–308. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |



{kind=link}
{kind=link}
{kind=link}
| Extracts/Fractions | % SI | % M | |
|---|---|---|---|
| M. persicae | M. incognita | M. javanica | |
| Hexane extract | 87.9 ± 2.1 ab | 100 ± 0.0 a | 100 ± 0.0 a |
| 1a | 60.3 ± 5.6 c | 25.0 ± 9.2 c | 5.53 ± 0.92 d |
| 1b | 97.6 ± 1.4 a | 100 ± 0 a | 100 ± 0.0 a |
| 1c | 94.4 ± 2.1 a | 100 ± 0 a | 100. ± 0.0 a |
| 1d | 90.9 ± 2.2 ab | 32.9 ± 7.8 b | 3.92 ± 1.13 d |
| 1e | 60.4 ± 7.7 c | 0 ± 0 d | 4.13 ± 0.52 d |
| 1f | 70.4 ± 6.9 bc | 0 ± 0 d | 3.92 ± 0.46 d |
| Ethyl acetate extract | 96.9 ± 1.2 a | 100 ± 0 a | 100 ± 0.0 a |
| 2a | nt | 20.7 ± 13.6 b | 11.39 ± 3.3 b |
| 2b | 97.6 ± 1.1 a | 100 ± 0 a | 100.0 ± 0.0 a |
| 2c | 91.3 ± 2.7 b | 100 ± 0 a | 100.0 ± 0.0 a |
| 2d | 89.7 ± 2.1 b | 0 ± 0 c | 13.47 ± 4.8 b |
| Compound | Retention Time (min) | Fractions (%) | |||
|---|---|---|---|---|---|
| 1b | 1c | 2b | 2c | ||
| Methyl decanoate | 7.27 | 4.75 | 6.55 | 7.08 | |
| Methyl undecanoate | 8.59 | 17.24 | 17.46 | 17.89 | |
| Methyl dodecanoate | 9.99 | 40.22 | 32.41 | 43.56 | 37.81 |
| Methyl tridecanoate | 11.25 | 10.42 | 12.29 | 13.67 | 10.58 |
| Methyl tetradecanoate | 12.41 | 9.07 | 12.51 | 12.77 | 7.51 |
| Methyl hexadecanoate | 14.52 | 2.18 | 4.08 | 2.46 | |
| 4,8,12,16-Tetramethylheptadecan-4-olide | 18.24 | 8.7 | |||
| Unknown | 22.04 | 17.02 | |||
| Unknown | 22.21 | 16.1 | 5.90 | ||
| Unknown | 27.61 | 7.50 | |||
| Compound | % SI | % OI | % M | |
|---|---|---|---|---|
| M. persicae | B. tabaci | M. incognita | M. javanica | |
| Decanoic acid | 89.7 ± 2.5 ab | 98.2 ± 1.7 a | 100 ± 0 a | 100.0 ± 0.0 a |
| Undecanoic acid | 83.3 ± 3.4 ab | 94.3 ± 3.3 a | 100 ± 0 a | 100.0 ± 0.0 a |
| Dodecanoic acid | 97.6 ± 1.4 a | 94.7 ± 2.9 a | 100 ± 0 a | 94.40 ± 0.4 b |
| Tridecanoic acid | 53.8 ± 8.9 c | Nt | 94.4 ± 6.4 a | nt |
| Tetradecanoic acid | 76.2 ± 4.7 b | 59.1 ± 9.2 b | 100 ± 0 a | 90.86 ± 2.5 b |
| Methyl decanoate | 36.3 ± 8.8 fg | 10.1 ± 6.2 de | 100 ± 0 a | 98.7 ± 0.5 a |
| Methyl undecanoate | 33.8 ± 7.1 fg | 15.7 ± 14.2 cde | nt | 95.4 ± 1.0 b |
| Methyl dodecanoate | 48.8 ± 7.1 efg | 0.0 ± 0 e | 100 ± 0 a | 14.53 ± 0.61 c |
| Methy tridecanoate | 57.7 ± 8.2 cdef | Nt | 74.8 ± 5.8 b | nt |
| Methyl tetradecanoate | 93.3 ± 0.2 ab | 16.4 ± 10.2 bcde | 100 ± 0 a | 10.24 ± 1.9 cd |
| Ethyl decanoate | 34.4 ± 6.9 fg | 30.7 ± 16.2 bcde | 55.9 ± 11.7 b | 5.91 ± 0.3 de |
| Ethyl undecanoate | 37.4 ± 34.1 fg | 46.6 ± 11.9 bcd | 53.9 ± 2.1 b | 16.71 ± 3.2 c |
| Ethy dodecanoate | 52.7 ± 7.6 defg | 62.9 ± 12 ab | 61.6 ± 3.9 b | 3.28 ± 0.2 e |
| Ethyl tetradecanoate | 77.4 ± 4.3 b | 42.2 ± 13.6 bcd | 63.8 ± 3.6 b | 3.64 ± 0.4 e |
| Compound | EC50 (nmol/cm2) | LC50 (nmol/mL) | ||
|---|---|---|---|---|
| M. persicae | B. tabaci | M. incognita | M. javanica | |
| Decanoic acid | * | 95 (91–99) a | 229 (218–240) a | 85 (83–87) b |
| Undecanoic acid | 21 (11–30) a | 6 (2–11) c | 192 (186–199) b | 64 (62–66) c |
| Dodecanoic acid | * | 49 (42–55) b | 231 (220–241) a | 368 (323–410) a |
| Extract | Lolium perenne a | Solanum lycopersicum a | |||
|---|---|---|---|---|---|
| Fatty Acid | Germination | Root Length | Leaf Length | Germination | Rootlet Length |
| Hexane extract | 62.16 ± 9.62 | 61.21 ± 8.05 | 81.36 ± 10.30 | 102.56 ± 0.0 | 94.49 ± 8.64 |
| Ethyl acetate extract | 72.97 ± 11.47 | 76.79 ± 8.61 | 80.83 ± 8.22 | 97.44 ± 7.52 | 67.96 ± 5.46 |
| Decanoic acid | 72.2 ± 10.1 | 62.3 ± 11.0 * | 50.5 ± 10.4 * | 125.8 ± 4.6 | 88.1 ± 12.0 |
| Undecanoic acid | 58.3 ± 7.5 * | 48.1 ± 11.0 * | 39.2 ± 13.4 * | 129.0 ± 0.0 | 87.1 ± 11.7 |
| Dodecanoic acid | 94.4 ± 4.5 | 58.7 ± 8.7 * | 46.4 ± 9.2 * | 122.6 ± 9.3 | 118.3 ± 13.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruz-Estrada, A.; Ruiz-Sánchez, E.; Cristóbal-alejo, J.; González-Coloma, A.; Andrés, M.F.; Gamboa-Angulo, M. Medium-Chain Fatty Acids from Eugenia winzerlingii Leaves Causing Insect Settling Deterrent, Nematicidal, and Phytotoxic Effects. Molecules 2019, 24, 1724. https://doi.org/10.3390/molecules24091724
Cruz-Estrada A, Ruiz-Sánchez E, Cristóbal-alejo J, González-Coloma A, Andrés MF, Gamboa-Angulo M. Medium-Chain Fatty Acids from Eugenia winzerlingii Leaves Causing Insect Settling Deterrent, Nematicidal, and Phytotoxic Effects. Molecules. 2019; 24(9):1724. https://doi.org/10.3390/molecules24091724
Chicago/Turabian StyleCruz-Estrada, Angel, Esaú Ruiz-Sánchez, Jairo Cristóbal-alejo, Azucena González-Coloma, María Fe Andrés, and Marcela Gamboa-Angulo. 2019. "Medium-Chain Fatty Acids from Eugenia winzerlingii Leaves Causing Insect Settling Deterrent, Nematicidal, and Phytotoxic Effects" Molecules 24, no. 9: 1724. https://doi.org/10.3390/molecules24091724
APA StyleCruz-Estrada, A., Ruiz-Sánchez, E., Cristóbal-alejo, J., González-Coloma, A., Andrés, M. F., & Gamboa-Angulo, M. (2019). Medium-Chain Fatty Acids from Eugenia winzerlingii Leaves Causing Insect Settling Deterrent, Nematicidal, and Phytotoxic Effects. Molecules, 24(9), 1724. https://doi.org/10.3390/molecules24091724