Terrestrial Microorganisms: Cell Factories of Bioactive Molecules with Skin Protecting Applications

 , ,
, ,  and
and
Abstract
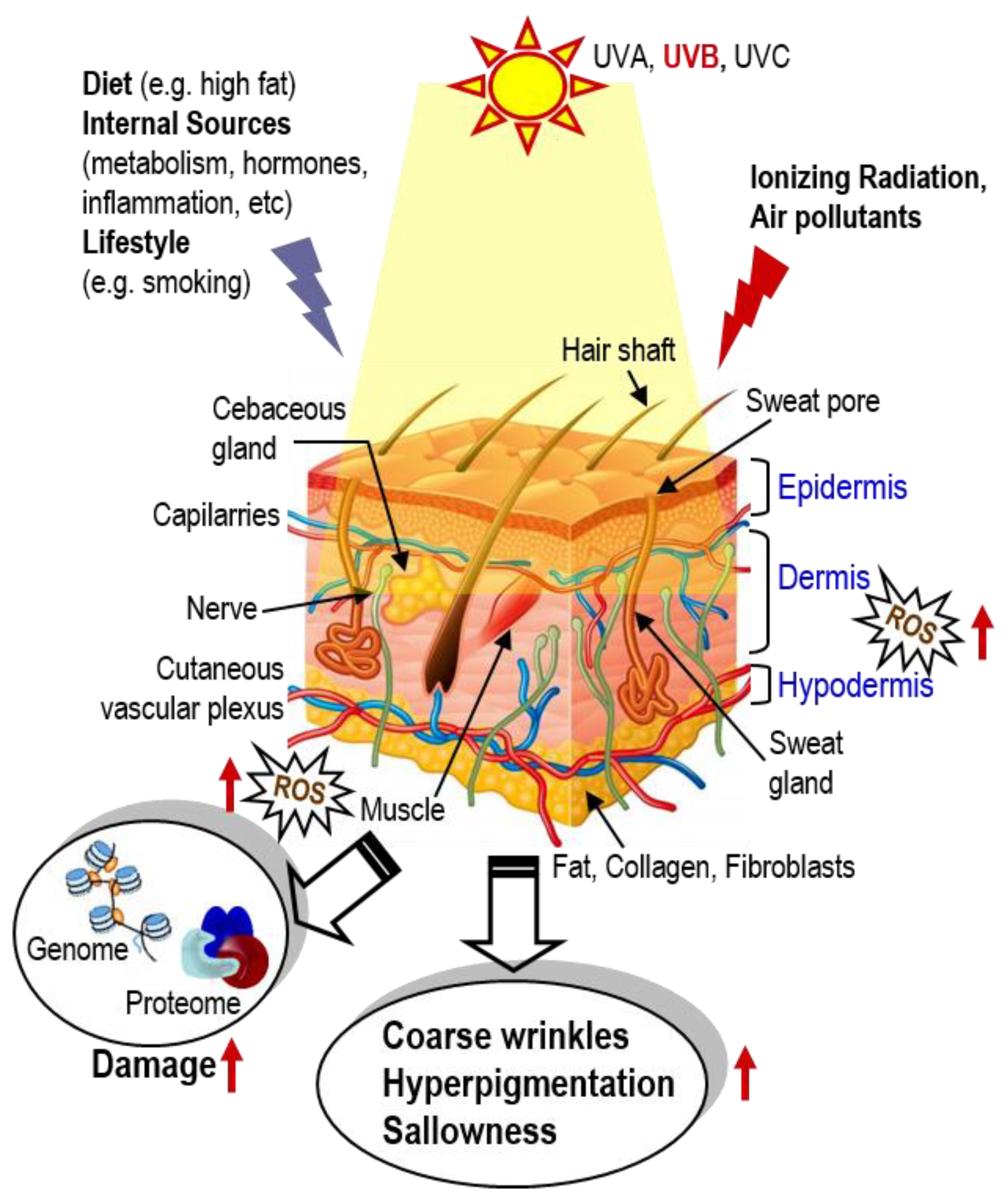
:1. Introduction
2. Antioxidants
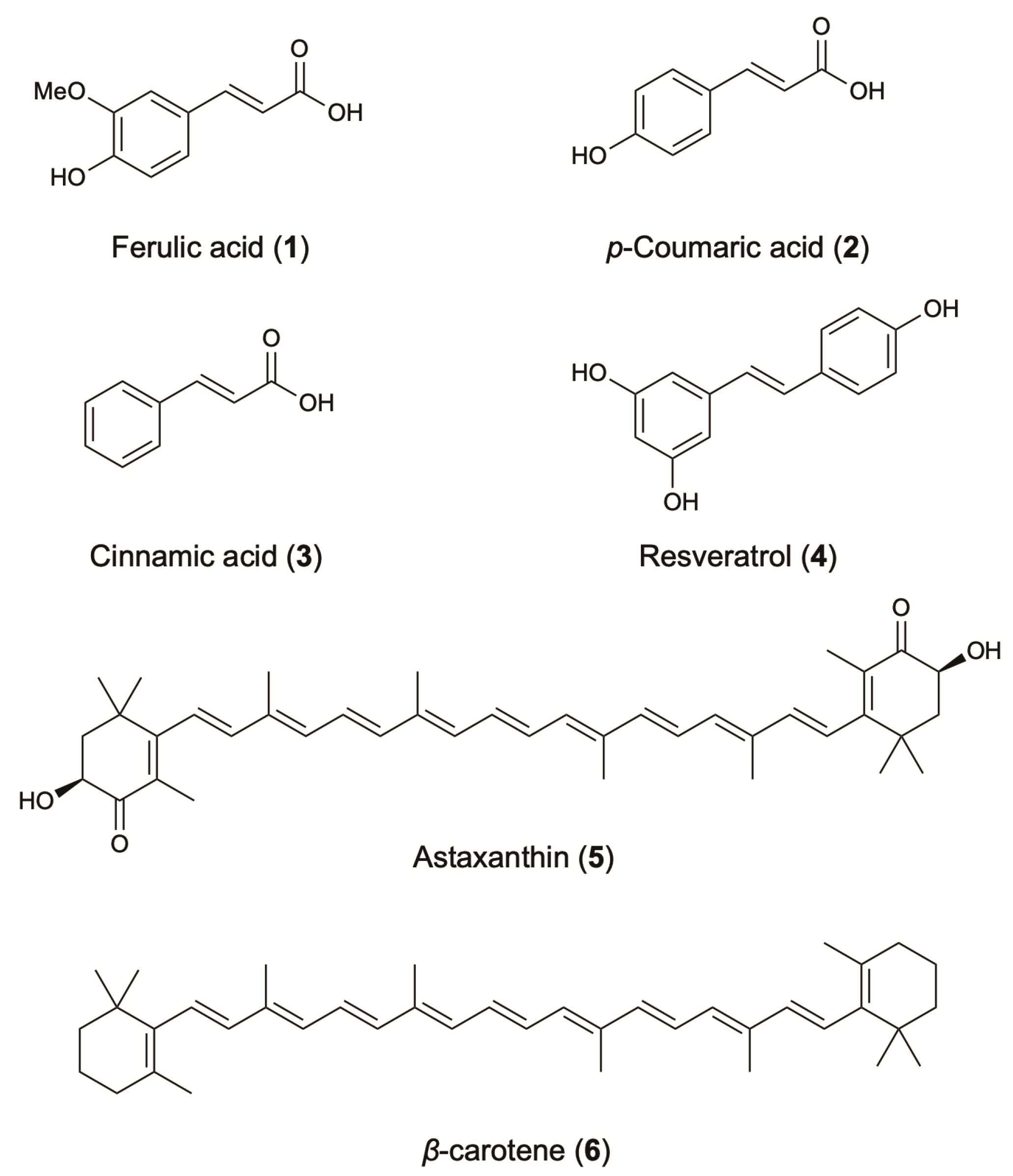
2.1. Phenolic Compounds
2.2. Carotenoids
2.3. Exopolysaccharides (EPSs)
2.4. Enzymes
3. Photo–Protective Agents
3.1. Melanins
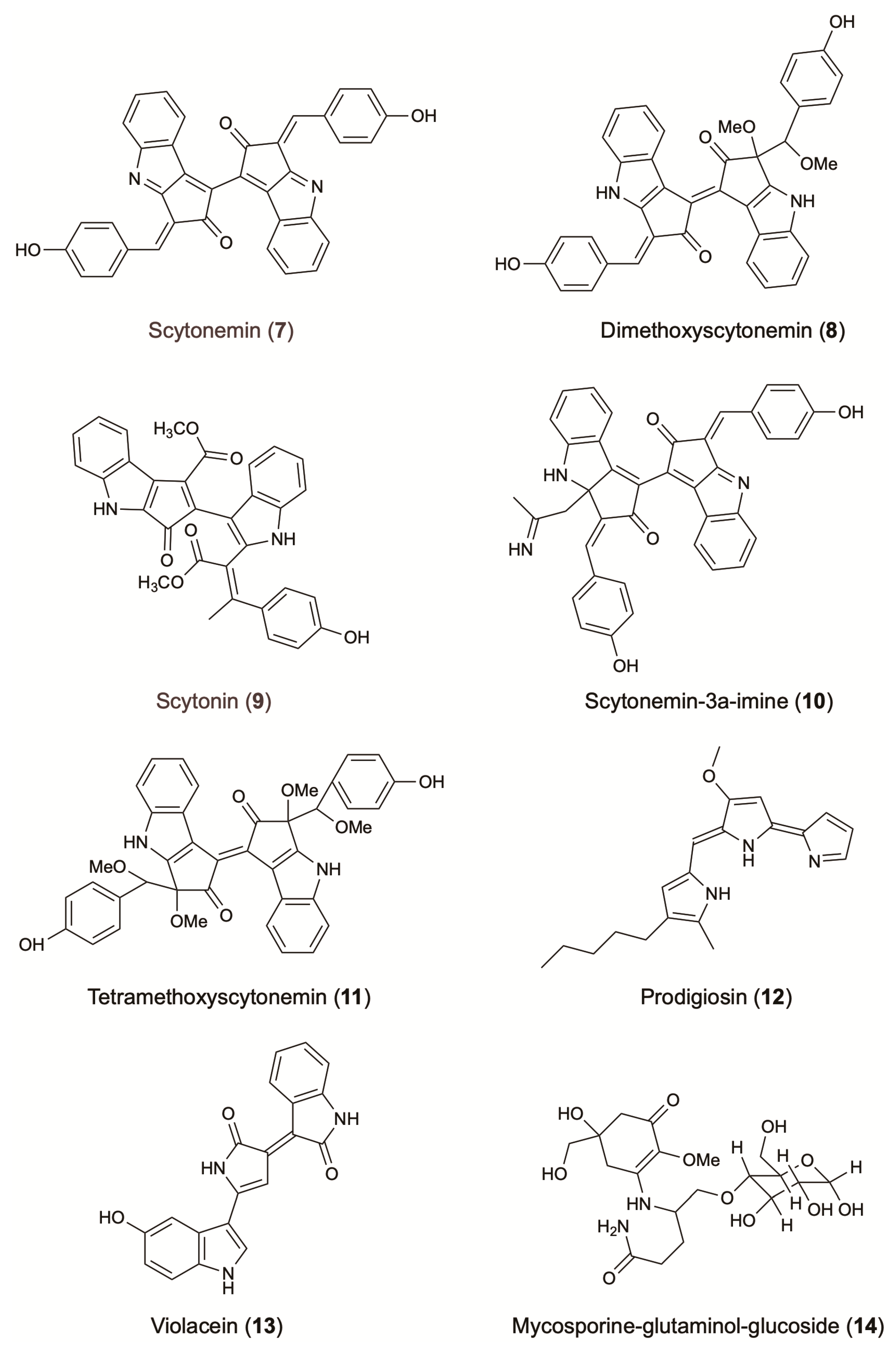
3.2. Indole and Pyrrole Derivatives
3.3. Mycosporines and Mycosporine-Like Amino Acids (MAAs)
4. Skin-Whitening Agents
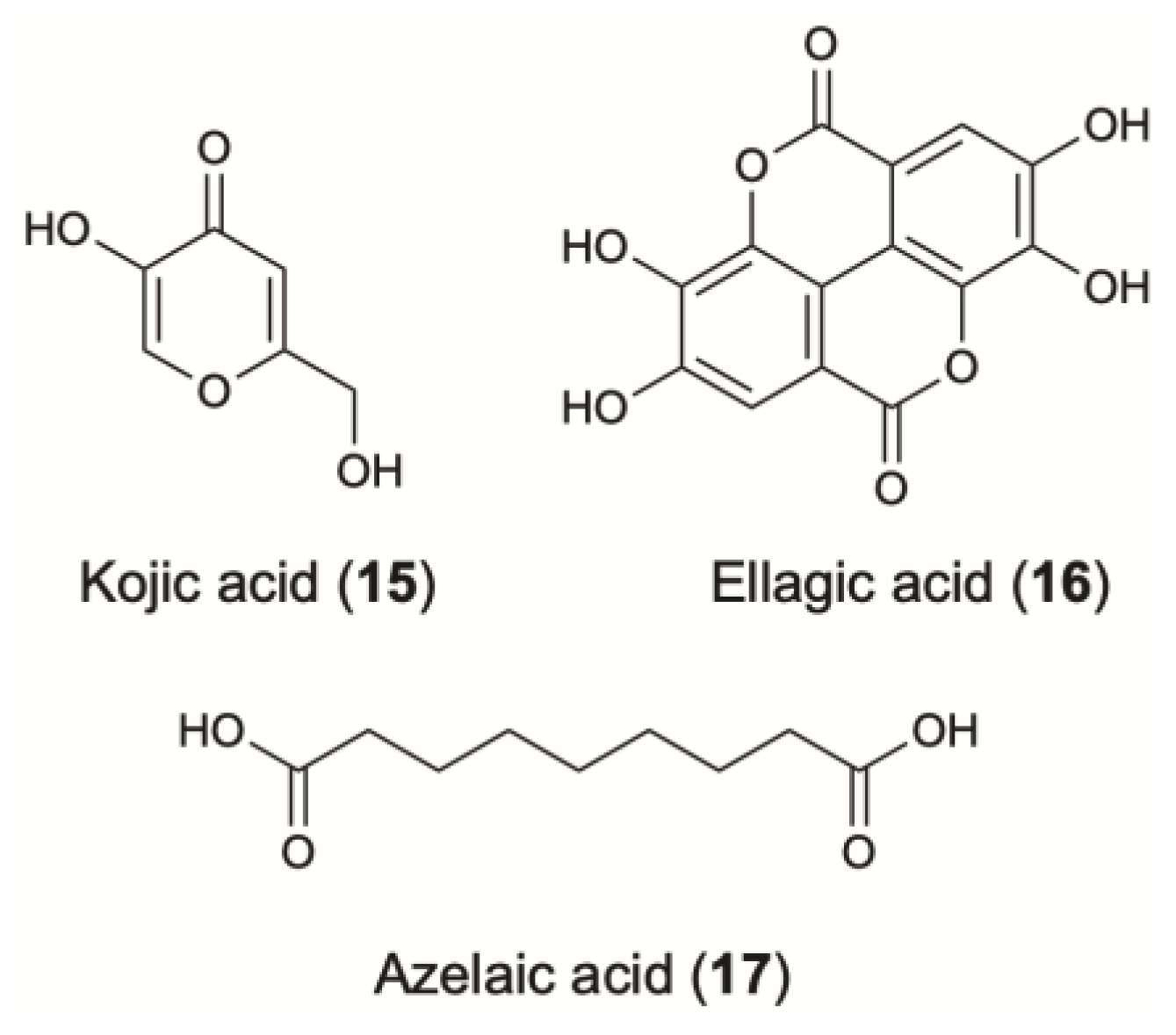
4.1. Pyrones
4.2. Phenolic Lactones
4.3. Carboxylic Acids
4.4. Enzymes and Derived Products
5. Additives and Other Active Ingredients
5.1. Antimicrobial Agents
5.2. Moisturizers and Biosurfactants
5.3. Pigments
5.4. Flavoring and Fragrances
6. Other Targets of Skin Protecting Interest
7. Targets for Future Developments
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABTS | 2,2′-Azino-bis-3-ethylbenzthiazolin-6-sulphonic acid |
| CALB | Candida antarctica lipase B |
| CAGR: | compound annual growth rate |
| CDW | cell dry weight |
| DPPH | 2,2-Diphenyl-1-picrylhydrazyl |
| EPSs | exopolysaccharides |
| HQ | hydroquinone |
| LTA | lipoteichoic acid |
| MAAs | mycosporine-like amino acids |
| MIC | minimum inhibitory concentration |
| γ-PGA | poly- γ-glutamic acid |
| RNS | reactive nitrogen species |
| ROS | reactive oxygen species |
| SPFs | sunscreen protection factors |
| SSR | solar-simulated radiation |
| SOD | superoxide dismutases |
| UVA | ultraviolet A |
| UVB | ultraviolet B |
References
- Ma, T.; Deng, Z.; Liu, T. Microbial production strategies and applications of lycopene and other terpenoids. World J. Microbiol. Biotechnol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Corinaldesi, C.; Barone, G.; Marcellini, F.; Dell’Anno, A.; Danovaro, R. Marine microbial-derived molecules and their potential use in cosmeceutical and cosmetic products. Mar. Drugs 2017, 15, 118. [Google Scholar] [CrossRef]
- Lobanovska, M.; Pilla, G. Penicillin’s discovery and antibiotic resistance: lessons for the future? J. Biol. Med. 2017, 90, 135–145. [Google Scholar]
- Raja, A.; Prabakarana, P. Actinomycetes and Drug-An Overview. Am. J. Drug Discov. Dev. 2011, 1, 75–84. [Google Scholar] [CrossRef]
- Berdy, J. Bioactive microbial metabolites. J. Antibiot. 2005, 58, 1–26. [Google Scholar] [PubMed]
- Ferreira, A.; Vecino, X.; Ferreira, D.; Cruz, J.M.; Moldes, A.B.; Rodrigues, L.R. Novel cosmetic formulations containing a biosurfactant from Lactobacillus paracasei. Colloids Surf. B Biointerfaces 2017, 155, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Argyropoulou, A.; Aligiannis, N.; Trougakos, I.P.; Skaltsounis, A.L. Natural compounds with anti-ageing activity. Nat. Prod. Rep. 2013, 30, 1412–1437. [Google Scholar] [CrossRef]
- Cavinato, M.; Jansen-Durr, P. Molecular mechanisms of UVB-induced senescence of dermal fibroblasts and its relevance for photoaging of the human skin. Exp. Gerontol. 2017, 94, 78–82. [Google Scholar]
- Velarde, M.C.; Demaria, M. Targeting senescent cells: Possible implications for delaying skin aging: A mini-review. Gerontology 2016, 62, 513–518. [Google Scholar]
- Trougakos, I.P.; Sesti, F.; Tsakiri, E.; Gorgoulis, V.G. Non-enzymatic post-translational protein modifications and proteostasis network deregulation in carcinogenesis. J. Proteomics 2013, 92, 274–298. [Google Scholar]
- Sklirou, A.; Papanagnou, E.D.; Fokialakis, N.; Trougakos, I.P. Cancer chemoprevention via activation of proteostatic modules. Cancer Lett. 2018, 413, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Market Research Reports. Available online: https://www.reuters.com/brandfeatures/venture-capital/article?id=30351 (accessed on 22 August 2017).
- Hyde, K.D.; Bahkali, A.H.; Moslem, M.A. Fungi—An unusual source for cosmetics. Fungal Divers. 2010, 43, 1–9. [Google Scholar] [CrossRef]
- CosIng Inventory. Available online: http://ec.europa.eu/growth/sectors/cosmetics/cosing_en (accessed on 22 March 2018).
- Chai, T.T.; Law, Y.C.; Wong, F.C.; Kim, S.K. Enzyme-assisted discovery of antioxidant peptides from edible marine invertebrates: A Review. Mar. Drugs 2017, 15, 42. [Google Scholar] [CrossRef] [PubMed]
- Kauppila, T.E.S.; Kauppila, J.H.K.; Larsson, N.G. Mammalian mitochondria and aging: an update. Cell. Metab. 2017, 25, 57–71. [Google Scholar] [CrossRef]
- Kusumawati, I.; Indrayanto, G. Natural antioxidants in cosmetics. Stud. Nat. Prod. Chem. 2013, 40, 485–505. [Google Scholar]
- Abramovič, H.; Grobin, B.; Ulrih, N.P.; Cigić, B. Relevance and standardization of in vitro antioxidant assays: ABTS, DPPH, and Folin–Ciocalteu. J. Chem. 2018, 2018, 1–9. [Google Scholar] [CrossRef]
- Huang, W.Y.; Cai, Y.Z.; Xing, J. A potential antioxidant resource: endophytic fungi from medicinal plants. Econ. Bot. 2007, 61, 14–30. [Google Scholar] [CrossRef]
- Danagoudar, A.; Joshi, C.G.; Kumar, R.S.; Poyya, J.; Nivya, T.; Hulikere, M.M.; Appaiah, K.A.A. Molecular profiling and antioxidant as well as anti-bacterial potential of polyphenol producing endophytic fungus-Aspergillus austroafricanus CGJ-B3. Mycology 2017, 8, 28–38. [Google Scholar] [CrossRef]
- Colla, L.M.; Furlong, E.B.; Costa, J.A.V. Antioxidant properties of Spirulina (Arthospira) platensis cultivated under different temperatures and nitrogen regimes. Braz. Arch. Biol. Technol. 2007, 50, 161–167. [Google Scholar] [CrossRef]
- Miranda, M.S.; Sato, S.; Mancini-Filho, J. Antioxidant activity of the microalga Chlorella vulgaris cultered on special conditions. Boll. Chim. Farm. 2001, 140, 165–168. [Google Scholar]
- Shalaby, E.S.; Shanab, S.M.M. Antiradical and antioxidant activities of different Spirulina platensis extracts against DPPH and ABTS radical assays. Indian J. Geomarine Sci. 2013, 42, 556–564. [Google Scholar] [CrossRef]
- Goiris, K.; Muylaert, K.; Voorspoels, S.; Noten, B.; De Paepe, D.; GJ, B.; De Cooman, L. Detection of flavonoids in microalgae from different evolutionary lineages. J. Phycol. 2014, 50, 483–492. [Google Scholar] [CrossRef]
- Jerez-Martel, I.; García-Poza, S.; Rodríguez-Martel, G.; Rico, M.; Afonso-Olivares, C.; Gómez-Pinchetti, J.L. Phenolic profile and antioxidant activity of crude extracts from microalgae and cyanobacteria strains. J. Food Qual. 2017, 4, 1–8. [Google Scholar] [CrossRef]
- Singh, D.P.; Prabha, R.; Verma, S.; Meena, K.K.; Yandigeri, M. Antioxidant properties and polyphenolic content in terrestrial cyanobacteria. 3 Biotech. 2017. [Google Scholar] [CrossRef]
- Liu, Y.; Nan, L.; Liu, J.; Yan, H.; Zhang, D.; Han, X. Isolation and identification of resveratrol-producing endophytes from wine grape Cabernet Sauvignon. SpringerPlus 2016. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zeng, Q.; Liu, Y.; Pan, Z. Alternaria sp. MG1, a resveratrol-producing fungus: isolation, identification, and optimal cultivation conditions for resveratrol production. Appl. Microbiol. Biotechnol. 2012, 95, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Baxter, R.A. Anti-aging properties of resveratrol: review and report of a potent new antioxidant skin care formulation. J. Cosmet. Dermatol. 2008, 7, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shi, J.; Liu, Y. Bioconversion of resveratrol using resting cells of non-genetically modified Alternaria sp. Biotechnol. Appl. Biochem. 2013, 60, 236–243. [Google Scholar] [CrossRef]
- An, S.M.; Koh, J.S.; Boo, Y.C. P-coumaric acid not only inhibits human tyrosinase activity in vitro but also melanogenesis in cells exposed to UVB. Phytother. Res. 2010, 24, 1175–1180. [Google Scholar]
- Lourith, N.; Kanlayavattanakul, M. Antioxidant activities and phenolics of Passiflora edulis seed recovered from juice production residue. J. Oleo. Sci. 2013, 62, 235–240. [Google Scholar] [CrossRef]
- Lopez-Burillo, S.; Tan, D.X.; Mayo, J.C.; Sainz, R.M.; Manchester, L.C.; Reiter, R.J. Melatonin, xanthurenic acid, resveratrol, EGCG, vitamin C and alpha-lipoic acid differentially reduce oxidative DNA damage induced by Fenton reagents: a study of their individual and synergistic actions. J. Pineal. Res. 2003, 34, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Satooka, H.; Kubo, I. Resveratrol as a kcat type inhibitor for tyrosinase: Potentiated melanogenesis inhibitor. Bioorg. Med. Chem. 2012, 20, 1090–1099. [Google Scholar] [CrossRef]
- Li, M.; Kildegaard, K.R.; Chen, Y.; Rodriguez, A.; Borodina, I.; Nielsen, J. De novo production of resveratrol from glucose or ethanol by engineered Saccharomyces cerevisiae. Metab. Eng. 2015, 32, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.G.; Fowler, Z.L.; Hueller, T.; Schaffer, S.; Koffas, M.A. High-yield resveratrol production in engineered Escherichia coli. Appl. Environ. Microbiol. 2011, 77, 3451–3460. [Google Scholar] [CrossRef]
- Sydor, T.; Schaffer, S.; Boles, E. Considerable increase in resveratrol production by recombinant industrial yeast strains with use of rich medium. Appl. Environ. Microbiol. 2010, 76, 3361–3363. [Google Scholar] [CrossRef]
- Balestrazzi, A.; Bonadei, M.; Calvio, C.; Mattivi, F.; Carbonera, D. Leaf-associated bacteria from transgenic white poplar producing resveratrol-like compounds: isolation, molecular characterization, and evaluation of oxidative stress tolerance. Can. J. Microbiol. 2009, 55, 829–840. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Zhang, Q.; Gao, Y.-Q.; Tang, J.-J.; Zhang, A.-L.; Gao, J.-M. Secondary metabolites from the endophytic Botryosphaeria dothidea of Melia azedarach and their antifungal, antibacterial, antioxidant, and cytotoxic activities. J. Agric. Food Chem. 2014, 62, 3584–3590. [Google Scholar] [CrossRef] [PubMed]
- Mou, Y.; Meng, J.; Fu, X.; Wang, X.; Tian, J.; Wang, M.; Peng, Y.; Zhou, L. Antimicrobial and antioxidant activities and effect of 1-hexadecene addition on palmarumycin C2 and C3 yields in liquid culture of endophytic fungus Berkleasmium sp. Dzf12. Molecules 2013, 18, 15587–15599. [Google Scholar] [CrossRef] [PubMed]
- Kerksick, C.; Willoughby, D. The antioxidant role of glutathione and N-acetyl-cysteine supplements and exercise-induced oxidative stress. J. Int. Soc. Sports Nutr. 2005, 2, 38–44. [Google Scholar] [CrossRef]
- Sonthalia, S.; Daulatabad, D.; Sarkar, R. Glutathione as a skin whitening agent: facts, myths, evidence and controversies. Indian J. Dermatol. Venereol. Leprol. 2016, 82, 262–272. [Google Scholar] [CrossRef]
- Fei, L.; Wang, Y.; Chen, S. Improved glutathione production by gene expression in Pichia pastoris. Bioprocess. Biosyst. Eng. 2009, 32, 729–735. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, J.; Wu, H.; Li, Z.; Ye, Q. Heterologous gshF gene expression in various vector systems in Escherichia coli for enhanced glutathione production. J. Biotechnol. 2015, 214, 63–68. [Google Scholar] [CrossRef]
- Palozza, P.; Krinsky, N.I. Astaxanthin and canthaxanthin are potent antioxidants in a membrane model. Arch. Biochem. Biophys. 1992, 267, 291–295. [Google Scholar] [CrossRef]
- Yamamoto, K.; Hara, K.Y.; Morita, T.; Nishimura, A.; Sasaki, D.; Ishii, J. Enhancement of astaxanthin production in Xanthophyllomyces dendrorhous by efficient method for the complete deletion of genes. Microb. Cell Fact. 2016, 15, 155. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, U.; Sarada, R.; Ravishankar, G.A. Effect of culture conditions on growth of green alga - Haematococcus pluvialis and astaxanthin production. Acta Physiol. Plant. 2002, 24, 323–329. [Google Scholar] [CrossRef]
- Mueller, L.; Boehm, V. Antioxidant activity of beta-carotene compounds in different in vitro assays. Molecules 2011, 16, 1055–1069. [Google Scholar] [CrossRef]
- Zhu, F.; Lu, L.; Fu, S.; Zhong, X.; Hu, M.; Deng, Z. Targeted engineering and scale up of lycopene overproduction in Escherichia coli. Process. Biochem. 2015, 50, 341–346. [Google Scholar] [CrossRef]
- Nelis, H.J.; De Leenheer, A. P, Reinvestigation of Brevibacterium sp. strain KY-4313 as a source of canthaxanthin. Appl. Environ. Microbiol. 1989, 55, 2505–2510. [Google Scholar]
- Sindhu, E.R.; Preethi, K.C.; Kuttan, R. Antioxidant activity of carotenoid lutein in vitro and in vivo. Indian J. Exp. Biol. 2010, 843–848. [Google Scholar]
- Del Campo, J.A.; Moreno, J.; Rodríguez, H.; Angeles Vargas, M.; Rivas, J.; Guerrero, M.G. Carotenoid content of chlorophycean microalgae: factors determining lutein accumulation in Muriellopsis sp. (Chlorophyta). J. Biotechnol. 2000, 76, 51–59. [Google Scholar] [CrossRef]
- Venugopalan, V.; Tripathi, S.K.; Nahar, P.; Saradhi, P.P.; Das, R.H.; Gautam, H.K. Characterization of canthaxanthin isomers isolated from a new soil Dietzia sp. and their antioxidant activities. J. Microbiol. Biotechnol. 2013, 23, 237–245. [Google Scholar] [CrossRef]
- Mahapatra, S.; Banerjee, D. Evaluation of in vitro antioxidant potency of exopolysaccharide from endophytic Fusarium solani SD5. Int. J. Biol. Macromol. 2013, 53, 62–66. [Google Scholar] [CrossRef]
- Zheng, L.P.; Zou, T.; Ma, Y.J.; Wang, J.W.; Zhang, Y.Q. Antioxidant and DNA damage protecting activity of exopolysaccharides from the endophytic bacterium Bacillus cereus SZ1. Molecules 2016, 21, 174. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Luo, J.; Ye, H.; Sun, Y.; Lu, Z.; Zeng, X. Production, characterization and antioxidant activities in vitro of exopolysaccharides from endophytic bacterium Paenibacillus polymyxa EJS-3. Carbohydr. Polym. 2009, 78, 275–281. [Google Scholar] [CrossRef]
- Liu, J.; Luo, J.; Ye, H.; Sun, Y.; Lu, Z.; Zeng, X. Medium optimization and structural characterization of exopolysaccharides from endophytic bacterium Paenibacillus polymyxa EJS-3. Carbohydr. Polym. 2010, 79, 206–213. [Google Scholar] [CrossRef]
- Xiao, R.; Zheng, Y. Overview of microalgal extracellular polymeric substances (EPS) and their applications. Biotechnol. Adv. 2016, 34, 1225–1244. [Google Scholar] [CrossRef]
- Chen, Y.; Mao, W.; Tao, H.; Zhu, W.; Qi, X.; Chen, Y. Structural characterization and antioxidant properties of an exopolysaccharide produced by the mangrove endophytic fungus Aspergillus sp. Y16. Bioresour. Technol. 2011, 102, 8179–8184. [Google Scholar] [CrossRef] [PubMed]
- Serrato, R.V.; Sassaki, G.L.; Cruz, L.M.; Pedrosa, F.O.; Gorin, P.A.; Iacomini, M. Culture conditions for the production of an acidic exopolysaccharide by the nitrogen-fixing bacterium Burkholderia tropica. Can. J. Microbiol. 2006, 52, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Trabelsi, L.; Chaieb, O.; Mnari, A.; Abid-Essafi, S.; Aleya, L. Partial characterization and antioxidant and antiproliferative activities of the aqueous extracellular polysaccharides from the thermophilic microalgae Graesiella sp. BMC Complement. Altern. Med. 2016. [Google Scholar] [CrossRef]
- Romay, C.; Gonzalez, R.; Ledon, N.; Remirez, D.; Rimbau, V. C-phycocyanin: a biliprotein with antioxidant, anti-inflammatory and neuroprotective effects. Curr. Protein Pept. Sci. 2003, 4, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Tasar, O.C.; Erdal, S.; Taskin, M. Chitosan production by psychrotolerant Rhizopus oryzae in non-sterile open fermentation conditions. Int. J. Biol. Macromol. 2016, 89, 428–433. [Google Scholar] [CrossRef]
- Asachi, R.; Karimi, K. Enhanced ethanol and chitosan production from wheat straw by Mucor indicus with minimal nutrient consumption. Process. Biochem. 2013, 48, 1524–1531. [Google Scholar] [CrossRef]
- Logesh, A.R.; Thillaimaharani, K.A.; Sharmila, K.; Kalaiselvam, M.; Raffi, S.M. Production of chitosan from endolichenic fungi isolated from mangrove environment and its antagonistic activity. Asian Pac. J. Trop. Biomed. 2012, 2, 140–143. [Google Scholar] [CrossRef]
- Ordonñez, L.; Garciía, J.; Bolanños, G. Producing chitin and chitin-glucan complexes from Aspergillus niger biomass using subcritical water. In Proceedings of the Ibero-american Conference on Supercritical Fluids, Cartagena, Colombia, 1–5 April 2013. [Google Scholar]
- Guo, J.; Rao, Z.; Yang, T.; Man, Z.; Xu, M.; Zhang, X. High-level production of melanin by a novel isolate of Streptomyces kathirae. FEMS Microbiol. Lett. 2014, 357, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Tarangini, K.; Mishra, S. Production of melanin by soil microbial isolate on fruit waste extract: two step optimization of key parameters. Biotechnol. Rep. 2014, 4, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Beckstead, A.A.; Zhang, Y.; Hilmer, J.K.; Smith, H.J.; Bermel, E.; Foreman, C.M. Ultrafast excited-state deactivation of the bacterial pigment violacein. J. Phys. Chem. A 2017. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, W.A.; Yusof, N.Z.; Nordin, N.; Zakaria, Z.A.; Rezali, M.F. Production and characterization of violacein by locally isolated Chromobacterium violaceum grown in agricultural wastes. Appl. Biochem. Biotechnol. 2012, 167, 1220–1234. [Google Scholar] [CrossRef]
- Wang, H.; Jiang, P.; Lu, Y.; Ruan, Z.; Jiang, R.; Xing, X.-H. Optimization of culture conditions for violacein production by a new strain of Duganella sp. B2. Biochem. Eng. J. 2009, 44, 119–124. [Google Scholar] [CrossRef]
- Pathak, J.; Sonker, A.S.; Richa, R.; Rajneesh, R.; Kannaujiya, V.K.; Singh, V.; Ahmed, H. Screening and partial purification of photoprotective pigment scytonemin from cyanobacterial crusts dwelling on the historical monuments in and around Varanasi, India. Microbiol. Res. 2017. [Google Scholar] [CrossRef]
- Chen, J.; Zhao, L.; Xu, J.; Yang, R.; He, S.; Yan, X. Determination of oxidized scytonemin in Nostoc commune Vauch cultured on different conditions by high performance liquid chromatography coupled with triple quadrupole mass spectrometry. J. Appl. Phycol. 2013, 25, 1001–1007. [Google Scholar] [CrossRef]
- Nakashima, T.; Anzai, K.; Kuwahara, N.; Komaki, H.; Miyadoh, S.; Harayama, S. Physicochemical characters of a tyrosinase inhibitor produced by Streptomyces roseolilacinus NBRC 12815. Biol. Pharm. Bull. 2009, 32, 832–836. [Google Scholar] [CrossRef]
- Kurbanoglu, E.B.; Ozdal, M.; Ozdal, O.G.; Algur, O.F. Enhanced production of prodigiosin by Serratia marcescens MO-1 using ram horn peptone. Braz. J. Microbiol. 2015, 46, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Boric, M.; Danevcic, T.; Stopar, D. Prodigiosin from Vibrio sp. DSM 14379; a new UV-protective pigment. Microb. Ecol. 2011, 62, 528–536. [Google Scholar] [CrossRef]
- Lawrence, K.P.; Gacesa, R.; Long, P.F.; Young, A.R. Molecular photoprotection of human keratinocytes in vitro by the naturally occurring mycosporine-like amino acid palythine. Br. J. Dermatol. 2017, 178, 1353–1363. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Incharoensakdi, A. Characterization of UV-screening compounds, mycosporine-like amino acids, and scytonemin in the cyanobacterium Lyngbya sp. CU2555. FEMS Microbiol. Ecol. 2014, 87, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Libkind, D.; Moline, M.; Sommaruga, R.; Sampaio, J.P.; van Broock, M. Phylogenetic distribution of fungal mycosporines within the Pucciniomycotina (Basidiomycota). Yeast (Chichester, England) 2011, 28, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Kogej, T.; Gostincar, C.; Volkmann, M.; Gorbushina, A.A.; Gunde-Cimerman, N.; Kogej, T.; Gorbushina, A.A. Mycosporines in extremophilic fungi - Novel complementary osmolytes? Environ. Chem. 2006, 3, 105–110. [Google Scholar] [CrossRef]
- Wei, S.; Xu, N.; Ji, Z. Identification of a kojic-acid producing Aspergillus flavus F52. Acta Microbiol. Sin. 2014, 1155–1160. [Google Scholar]
- El-Aasar, S.A. Cultural conditions studies on kojic acid production by Aspergillus parasiticus. Int. J. Agric. Biol. 2006, 8, 468–473. [Google Scholar]
- Mohamad, R.; Ariff, A.B. Biotransformation of various carbon sources to kojic acid by cell-bound enzyme system of A. flavus Link 44-1. Biochem. Eng. J. 2007, 35, 203–209. [Google Scholar] [CrossRef]
- Zhao, L.; Kim, J.C.; Paik, M.J.; Lee, W.; Hur, J.S. A multifunctional and possible skin UV protectant, (3R)-5-hydroxymellein, produced by an endolichenic fungus isolated from Parmotrema austrosinense. Molecules 2016, 22, 26. [Google Scholar] [CrossRef]
- Robledo, A.; Aguilera-Carbo, A.; Rodriguez, R.; Martinez, J.L.; Garza, Y.; Aguilar, C.N. Ellagic acid production by Aspergillus niger in solid state fermentation of pomegranate residues. J. Ind. Microbiol. Biotechnol. 2008, 35, 507–513. [Google Scholar] [CrossRef]
- Smith, W.P. The effects of topical L(+) lactic acid and ascorbic acid on skin whitening. Int. J. Cosmet. Sci. 1999, 21, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Maas, R.H.; Springer, J.; Eggink, G.; Weusthuis, R.A. Xylose metabolism in the fungus Rhizopus oryzae: effect of growth and respiration on L+-lactic acid production. J. Ind. Microbiol. Biotechnol. 2008, 35, 569–578. [Google Scholar] [CrossRef]
- Shih, I.L.; Van, Y.T. The production of poly-(gamma-glutamic acid) from microorganisms and its various applications. Bioresour. Technol. 2001, 79, 207–225. [Google Scholar] [CrossRef]
- Schallreuter, K.U.; Wood, J.W. A possible mechanism of action for azelaic acid in the human epidermis. Arch. Dermatol. Res. 1990, 282, 168–171. [Google Scholar] [CrossRef]
- Balina, L.M.; Graupe, K. The treatment of melasma, 20% azelaic acid versus 4% hydroquinone cream. Int. J. Dermatol. 1991, 30, 893–895. [Google Scholar] [CrossRef]
- Nazzaro-Porro, M.; Passi, S.; Morpurgo, G.; Breathnach, A.S. Identification of tyrosinase inhibitors in cultures of Pityrosporum and their melanocytotoxic effect. Pigment. Cell 1979, 4, 234–243. [Google Scholar]
- Yin, C.; Zhang, C.; Gao, M. Enzyme-catalyzed synthesis of vitamin E succinate using a chemically modified novozym-435. Chin. J. Chem. Eng. 2011, 19, 135–139. [Google Scholar] [CrossRef]
- Wicken, A.J.; Gibbens, J.W.; Knox, K.W. Comparative studies on the isolation of membrane lipoteichoic acid from Lactobacillus fermenti. J. Bacteriol. 1973, 113, 365–372. [Google Scholar]
- Tokiwa, Y.; Kitagawa, M.; Raku, T. Enzymatic synthesis of arbutin undecylenic acid ester and its inhibitory effect on mushroom tyrosinase. Biotechnol. Lett. 2007, 29, 481–486. [Google Scholar] [CrossRef]
- Mapari, S.A.; Meyer, A.S.; Thrane, U.; Frisvad, J.C. Identification of potentially safe promising fungal cell factories for the production of polyketide natural food colorants using chemotaxonomic rationale. Microb. Cell Fact. 2009. [Google Scholar] [CrossRef]
- Dufossé, L. Encyclopedia of Microbiology, 3rd ed.; Academic Press: New York, NY, USA, 2009; Microbial Pigments; pp. 457–471. [Google Scholar]
- Buenger, J.; Driller, H. Ectoin: An effective natural substance to prevent UVA-induced premature photoaging. Skin Pharmacol. Physiol. 2004, 17, 232–237. [Google Scholar] [CrossRef]
- Becker, J.; Schafer, R.; Kohlstedt, M.; Harder, B.J.; Borchert, N.S.; Stoveken, N. Systems metabolic engineering of Corynebacterium glutamicum for production of the chemical chaperone ectoine. Microb. Cell Fact. 2013. [Google Scholar] [CrossRef] [PubMed]
- Botta, C.; Di Giorgio, C.; Sabatier, A.S.; De Meo, M. Genotoxicity of visible light (400–800 nm) and photoprotection assessment of ectoin, L-ergothioneine and mannitol and four sunscreens. J. Photochem. Photobiol. B. 2008, 91, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Pugh, S.; McKenna, R.; Halloum, I.; Nielsen, D.R. Engineering Escherichia coli for renewable benzyl alcohol production. Metab. Eng. 2015, 2, 39–45. [Google Scholar] [CrossRef]
- Ni, J.; Tao, F.; Du, H.; Xu, P. Mimicking a natural pathway for de novo biosynthesis: natural vanillin production from accessible carbon sources. Sci. Rep. 2015. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Okada, Y.; Yoshida, T.; Nagasawa, T. Vanillin production using Escherichia coli cells over-expressing isoeugenol monooxygenase of Pseudomonas putida. Biotechnol. Lett. 2008, 30, 665–670. [Google Scholar] [CrossRef]
- Zhao, L.-Q.; Sun, Z.-H.; Zheng, P.; Zhu, L.-L. Biotransformation of isoeugenol to vanillin by a novel strain of Bacillus fusiformis. Biotechnol. Lett. 2005, 27, 1505–1509. [Google Scholar] [CrossRef]
- Ravasio, D.; Wendland, J.; Walther, A. Major contribution of the Ehrlich pathway for 2-phenylethanol/rose flavor production in Ashbya gossypii. FEMS Yeast Res. 2014, 14, 833–844. [Google Scholar] [CrossRef]
- Etschmann, M.M.W.; Schrader, J. An aqueous–organic two-phase bioprocess for efficient production of the natural aroma chemicals 2-phenylethanol and 2-phenylethylacetate with yeast. Appl. Microbiol. Biotechnol. 2006, 71, 440–443. [Google Scholar] [CrossRef]
- Willrodt, C.; David, C.; Cornelissen, S.; Buhler, B.; Julsing, M.K.; Schmid, A. Engineering the productivity of recombinant Escherichia coli for limonene formation from glycerol in minimal media. Biotechnol. J. 2014, 9, 1000–1012. [Google Scholar] [CrossRef]
- Dobler, L.; de Carvalho, B.R.; Alves, S.; Neves, B.C.; Freire, G.; Almeida, R.V. Enhanced rhamnolipid production by Pseudomonas aeruginosa overexpressing estA in a simple medium. PloS ONE 2017, 12, e0183857. [Google Scholar] [CrossRef]
- Desai, J.D.; Banat, I.M. Microbial production of surfactants and their commercial potential. Microbiol. Mol. Biol. Rev. 1997, 47–64. [Google Scholar]
- Castelblanco-Matiz, L.M.; Barbachano-Torres, A.; Ponce-Noyola, T.; Ramos-Valdivia, A.C.; Cerda García-Rojas, C.M.; Flores-Ortiz, C.M. Carotenoid production and gene expression in an astaxanthin-overproducing Xanthophyllomyces dendrorhous mutant strain. Arch. Microbiol. 2015, 197, 1129–1139. [Google Scholar] [CrossRef] [PubMed]
- Berman, J.; Zorrilla-Lopez, U.; Farre, G.; Zhu, C.; Sandmann, G.; Twyman, R.M.; Capell, T.; Christou, P. Nutritionally important carotenoids as consumer products. Phytochem. Rev. 2015, 14, 727–743. [Google Scholar] [CrossRef]
- Henríquez, V.; Escobar, C.; Galarza, J.; Gimpel, J. Carotenoids in microalgae. In Carotenoids in Nature; Strange, E., Ed.; Springer International Publishing: New York, NY, USA, 2016; pp. 219–237. [Google Scholar]
- Lau, N.-S.; Matsui, M.; Abdullah, A.A.-A. Cyanobacteria: photoautotrophic microbial factories for the sustainable synthesis of industrial products. J. Biomed. Biotechnol. 2015, 2, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Yoon, S.H.; Lee, S.H.; Kim, J.Y.; Oh, D.K.; Kim, S.W. An update on microbial carotenoid production: application of recent metabolic engineering tools. Appl. Microbiol. Biotechnol. 2007, 77, 505–512. [Google Scholar] [CrossRef]
- Mata-Gomez, L.C.; Montanez, J.C.; Mendez-Zavala, A.; Aguilar, C.N. Biotechnological production of carotenoids by yeasts: An overview. Microb. Cell Fact. 2014. [Google Scholar] [CrossRef]
- Liu, J.; Wang, X.; Pu, H.; Liu, S.; Kan, J.; Jin, C. Recent advances in endophytic exopolysaccharides: Production, structural characterization, physiological role and biological activity. Carbohydr. Polym. 2017, 157, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Ooi, V.E.; Chang, S.T. Free radical scavenging activities of mushroom polysaccharide extracts. Life Sci. 1997, 60, 763–771. [Google Scholar] [CrossRef]
- Jin, M.; Cai, Y.X.; Li, J.R.; Zhao, H. 1,10-phenanthroline-Fe2+ oxidative assay of hydroxyl radical produced by H2O2/Fe. Prog. Biochem. Biophys. 1996, 23, 553–555. [Google Scholar]
- Chen, B.; You, W.; Huang, J.; Yu, Y.; Chen, W. Isolation and antioxidant property of the extracellular polysaccharide from Rhodella reticulata. World J. Microbiol. Biotechnol. 2010, 26, 833–840. [Google Scholar] [CrossRef]
- Cirulis, J.T.; Scott, J.A.; Ross, G.M. Management of oxidative stress by microalgae. Can. J. Physiol. Pharmacol. 2013, 91, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Hassan, H.M. Cytotoxicity of oxyradicals and the evolution of superoxide dismutases. In Oxygen, Gene Expression and Cellular Function; Massaro, D., Clerch, L., Eds.; Marcel Dekker: New York, NY, USA, 1997. [Google Scholar]
- Bruno-Barcena, J.M.; Azcarate-Peril, M.A.; Hassan, H.M. Role of antioxidant enzymes in bacterial resistance to organic acids. Appl. Environ. Microbiol. 2010, 76, 2747–2753. [Google Scholar] [CrossRef] [PubMed]
- Rahman, K. Studies on free radicals, antioxidants, and co-factors. Clin. Interv. Aging 2007, 2, 219–236. [Google Scholar]
- Abbott, D.A.; Suir, E.; Duong, G.H.; de Hulster, E.; Pronk, J.T.; van Maris, A.J. Catalase overexpression reduces lactic acid-induced oxidative stress in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2009, 75, 2320–2325. [Google Scholar] [CrossRef]
- Shao, N.; Beck, C.F.; Lemaire, S.D.; Krieger-Liszkay, A. Photosynthetic electron flow affects H2O2 signaling by inactivation of catalase in Chlamydomonas reinhardtii. Planta 2008, 228, 1055–1066. [Google Scholar] [CrossRef]
- European Commission. Ban on Animal Testing. Available online: https://ec.europa.eu/growth/sectors/cosmetics/animal-testing_en (accessed on 14 April 2019).
- Gao, Q.; Garcia-Pichel, F. Microbial ultraviolet sunscreens. Nat. Rev. Microbiol. 2011, 9, 791–802. [Google Scholar] [CrossRef]
- Plonka, P.M.; Grabacka, M. Melanin synthesis in microorganisms-biotechnological and medical aspects. Acta Biochim. Pol. 2006, 53, 429–443. [Google Scholar]
- Nosanchuk, J.D.; Stark, R.E.; Casadevall, A. Fungal melanin: What do we know about structure? Front. Microbiol. 2015. [Google Scholar] [CrossRef]
- Sansinenea, E.; Ortiz, A. Melanin: a photoprotection for Bacillus thuringiensis based biopesticides. Biotechnol. Lett. 2015, 37, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K.; Nazifi, E.; Hirai, Y.; Wada, N.; Matsugo, S.; Sakamoto, T. The cyanobacterial UV-absorbing pigment scytonemin displays radical-scavenging activity. J. Gen. Appl. Microbiol. 2012, 58, 137–144. [Google Scholar] [CrossRef]
- Garcia-Pichel, F.; Sherry, N.D.; Castenholz, R.W. Evidence for an ultraviolet sunscreen role of the extracellular pigment scytonemin in the terrestrial cyanobacterium Chlorogloeopsis sp. Photochem. Photobiol. 1992, 56, 17–23. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Sonani, R.R.; Madamwar, D. Cyanobacterial sunscreen scytonemin: role in photoprotection and biomedical research. Appl. Biochem. Biotechnol. 2015, 176, 1551–1563. [Google Scholar] [CrossRef]
- Darshan, N.; Manonmani, H.K. Prodigiosin and its potential applications. J. Food Sci. Technol. 2015, 52, 5393–5407. [Google Scholar] [CrossRef]
- Duran, N.; Justo, G.Z.; Duran, M.; Brocchi, M.; Cordi, L.; Tasic, L. Advances in Chromobacterium violaceum and properties of violacein-Its main secondary metabolite: A review. Biotechnol. Adv. 2016, 34, 1030–1045. [Google Scholar] [CrossRef]
- Colabella, F.; Moline, M.; Libkind, D. UV sunscreens of microbial origin: mycosporines and mycosporine- like aminoacids. Recent Pat. Biotechnol. 2014, 8, 179–193. [Google Scholar] [CrossRef]
- Khosravi, S.; Khodabandeh, S.; Agh, N.; Bakhtiarian, M. Effects of salinity and ultraviolet radiation on the bioaccumulation of mycosporine-like amino acids in Artemia from Lake Urmia (Iran). Photochem. Photobiol. 2013, 89, 400–405. [Google Scholar] [CrossRef]
- Gorbushina, A.A.; Whitehead, K.; Dornieden, T.; Niesse, A.; Schulte, A.; Hedges, J.I. Black fungal colonies as units of survival: hyphal mycosporines synthesized by rock-dwelling microcolonial fungi. Can. J. Bot. 2003, 81, 131–138. [Google Scholar] [CrossRef]
- Gillbro, J.M.; Olsson, M.J. The melanogenesis and mechanisms of skin-lightening agents-existing and new approaches. Int. J. Cosmet. Sci. 2011, 33, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Smit, N.; Vicanova, J.; Pavel, S. The hunt for natural skin whitening agents. Int. J. Mol. Sci. 2009, 10, 5326–5349. [Google Scholar] [CrossRef] [PubMed]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: medicinal chemistry perspective of tyrosinase inhibitors. J. Enzyme Inhib Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef]
- Pillaiyar, T.; Namasivayam, V.; Manickam, M.; Jung, S.H. Inhibitors of Melanogenesis: An Updated Review. J. Med. Chem. 2018, 61, 7395–7418. [Google Scholar] [CrossRef] [PubMed]
- Ha, T.J.; Yang, M.S.; Jang, D.S.; Choe, S.U.; Park, G.H. Inhibitory activities of flavanone derivatives isolated from Sophora flavecens for melanogenesis. Bull. Korean Chem. Soc. 2001, 22, 97–99. [Google Scholar]
- Parvez, S.; Kang, M.; Chung, H.S.; Cho, C.; Hong, M.C.; Shin, M.K. Survey and mechanism of skin depigmenting and lightening agents. Phytother. Res. 2006, 20, 921–934. [Google Scholar] [CrossRef]
- Rosfarizan, M.; Mohd, S.M.; Nurashikin, S.; Madihah, M.S.; Arbakariya, B.A. Kojic acid: Applications and development of fermentation process for production. Biotech. Mol. Biol. 2010, 5, 24–37. [Google Scholar]
- Ortiz-Ruiz, C.V.; Berna, J.; Tudela, J.; Varon, R.; Garcia-Canovas, F. Action of ellagic acid on the melanin biosynthesis pathway. J. Dermatol. Sci. 2010, 5, 24–37. [Google Scholar] [CrossRef]
- Ventura, J.; Belmares, R.; Aguilera-Carbo, A.; Gutiérrez-Sanchez, G.; Rodríguez-Herrera, R. Fungal biodegradation of tannins from Creosote bush (Larrea tridentata) and Tar bush (Fluorensia cernua) for gallic and ellagic acid production. Food Technol. Biotechnol. 2008, 46, 213–217. [Google Scholar]
- Sepúlveda, L.; Ascacio, A.; Rodríguez-Herrera, R.; Aguilera-Carboó, A.; Aguilar, C.N. Ellagic acid: Biological properties and biotechnological development for production processes. Afr. J. Biotechnol. 2011, 10, 4518–4523. [Google Scholar]
- Breathnach, A.C.; Nazzaro-Porro, M.; Passi, S.; Zina, G. Azelaic acid therapy in disorders of pigmentation. Clin. Dermatol. 1989, 7, 106–119. [Google Scholar] [CrossRef]
- Sieber, M.A.; Hegel, J.K. Azelaic acid: Properties and mode of action. Skin Pharmacol. Physiol. 2014, 27, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Breathnach, A.S. Melanin hyperpigmentation of skin: melasma, topical treatment with azelaic acid, and other therapies. Cutis 1989, 7, 106–119. [Google Scholar]
- Usuki, A.; Ohashi, A.; Sato, H.; Ochiai, Y.; Ichihashi, M.; Funasaka, Y. The inhibitory effect of glycolic acid and lactic acid on melanin synthesis in melanoma cells. Exp. Dermatol. 2003, 12, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Jin, B.; Kelly, J.M. Production of lactic acid from renewable materials by Rhizopus fungi. Biochem. Eng. J. 2007, 35, 251–263. [Google Scholar] [CrossRef]
- Liu, X.; Liu, F.; Liu, S.; Li, H.; Ling, P.; Zhu, X. Poly-gamma-glutamate from Bacillus subtilis inhibits tyrosinase activity and melanogenesis. Appl. Microbiol. Biotechnol. 2013, 97, 9801–9809. [Google Scholar] [CrossRef]
- Mohorčič, M.; Friedrich, J.; Renimel, I.; André, P.; Mandin, D.; Chaumont, J.-P. Production of melanin bleaching enzyme of fungal origin and its application in cosmetics. Biotechnol. Bioprocess. Eng. 2007, 12, 200–206. [Google Scholar] [CrossRef]
- Antonopoulou, I.; Varriale, S.; Topakas, E.; Rova, U.; Christakopoulos, P.; Faraco, V. Enzymatic synthesis of bioactive compounds with high potential for cosmeceutical application. Appl. Microbiol. Biotechnol. 2016, 100, 6519–6543. [Google Scholar] [CrossRef]
- Sugimoto, K.; Nomura, K.; Nishimura, T.; Kiso, T.; Sugimoto, K.; Kuriki, T. Syntheses of alpha-arbutin-alpha-glycosides and their inhibitory effects on human tyrosinase. J. Biosci. Bioeng. 2005, 99, 272–276. [Google Scholar] [CrossRef]
- Sekhon Randhawa, K.K.; Rahman, P.K. Rhamnolipid biosurfactants-past, present, and future scenario of global market. Front. Microbiol. 2014. [Google Scholar] [CrossRef]
- Marchant, R.; Banat, I.M. Biosurfactants: a sustainable replacement for chemical surfactants? Biotechnol. Lett. 2012, 34, 1597–1605. [Google Scholar] [CrossRef] [PubMed]
- Lang, S.; Wullbrandt, D. Rhamnose lipids-biosynthesis, microbial production and application potential. Appl. Microbiol. Biotechnol. 1999, 51, 22–32. [Google Scholar] [CrossRef]
- Haba, E.; Pinazo, A.; Jauregui, O.; Espuny, M.J.; Infante, M.R.; Manresa, A. Physicochemical characterization and antimicrobial properties of rhamnolipids produced by Pseudomonas aeruginosa 47T2 NCBIM 40044. Biotechnol. Bioeng. 2003, 81, 316–322. [Google Scholar] [CrossRef]
- Piljac, G.; Piljac, T. Use of rhamnolipids in wound healing, treating burn shock, atherosclerosis, organ transplants, depression, schizophrenia and cosmetics. U.S. Patent No US7262171B1. 2000.
- Tuli, H.S.; Chaudhary, P.; Beniwal, V.; Sharma, A.K. Microbial pigments as natural color sources: current trends and future perspectives. J. Food Sci. Tech. 2015, 52, 4669–4678. [Google Scholar] [CrossRef] [PubMed]
- Caro, Y.; Venkatachalam, M.; Lebeau, J.; Fouillaud, M.; Dufossé, L. Pigments and colorants from filamentous fungi. Fungal Metab. 2017, 499–568. [Google Scholar] [CrossRef]
- Souza, P.N.; Grigoletto, T.L.; Abreu, L.M.; Guimarães, L.H.; Santos, C.; Galvão, L.R.; Cardoso, P.G. Production and chemical characterization of pigments in filamentous fungi. Microbiology 2016, 162, 12–22. [Google Scholar]
- Rao, N.; Prabhu, M.; Xiao, M.; Li, W.J. Fungal and bacterial pigments: Secondary metabolites with wide applications. Front. Microbiol. 2017. [Google Scholar] [CrossRef]
- Kuddus, M.; Singh, P.; Thomas, G.; Al-Hazimi, A. Recent developments in production and biotechnological applications of C-phycocyanin. Biomed. Res. Int. 2013. [Google Scholar] [CrossRef] [PubMed]
- Nicoletti, M. Microalgae Nutraceuticals. Foods 2016, 5, 54. [Google Scholar] [CrossRef] [PubMed]
- Graverholt, O.S.; Eriksen, N.T. Heterotrophic high-cell-density fed-batch and continuous-flow cultures of Galdieria sulphuraria and production of phycocyanin. Appl. Microbiol. Biotechnol. 2007, 77, 69–75. [Google Scholar] [CrossRef]
- Carroll, A.L.; Desai, S.H.; Atsumi, S. Microbial production of scent and flavor compounds. Curr. Opin. Chem. Biol. 2016, 37, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Longo, M.A.; Sanromán, M.A. Production of food aroma compounds: microbial and enzymatic methodologies. Food Technol. Biotechnol. 2006, 44, 335–353. [Google Scholar]
- Kempf, B.; Bremer, E. Uptake and synthesis of compatible solutes as microbial stress responses to high-osmolality environments. Arch. Microbiol. 1998, 170, 319–330. [Google Scholar] [CrossRef]
- Soccol, C.R.; Medeiros, A.B.; Vandenberghe, L.P.; Woiciechowski, A.L. Flavor compounds produced by fungi, yeasts, and bacteria. Handb. Food Prod. Manuf. 2007, 1, 179–191. [Google Scholar]
- Vandamme, E.J. Bioflavours and fragrances via fungi and their enzymes. Fungal Divers. 2003, 13, 153–166. [Google Scholar]
- Carrau, F.; Boido, E.; Dellacassa, E. Yeast Diversity and Flavor Compounds. In Fungal Metabolites; Mérillon, J.M., Ramawat, K., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 1–29. [Google Scholar]
- Priefert, H.; Rabenhorst, J.; Steinbüchel, A. Biotechnological production of vanillin. Appl. Microbiol. Biotechnol. 2001, 56, 296–314. [Google Scholar] [CrossRef]
- Kunjapur, A.M.; Tarasova, Y.; Prather, K.L.J. Synthesis and accumulation of aromatic aldehydes in an engineered strain of Escherichia coli. J. Am. Chem. Soc. 2014, 136, 11644–11654. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Gutierrez, J.; Chan, R.; Batth, T.S.; Adams, P.D.; Keasling, J.D.; Petzold, C.J. Metabolic engineering of Escherichia coli for limonene and perillyl alcohol production. Metab. Eng. 2013, 19, 33–41. [Google Scholar] [CrossRef]
- Okino, T.; Qi, S.; Matsuda, H.; Murakami, M.; Yamaguchi, K. Nostopeptins A and B, elastase inhibitors from the cyanobacterium Nostoc minutum. J. Nat. Prod. 1997, 60, 158–161. [Google Scholar] [CrossRef]
- Georgousaki, K.; DePedro, N.; Chinchilla, A.; Aliagiannis, N.; Vicente, F.; de Castro, P.; Fotinos, S.; Genilloud, O.; Fokialakis, N. High throughput screening of microbial biodiversity for the discovery of novel cosmeceutical agents. Panta Med. 2015. [Google Scholar] [CrossRef]
- Georgousaki, K.; Tsafantakis, N.; Cheilari, A.; Gumeni, S.; González, I.; González, V.; José, R.T.; Genilloud, O.; Fotinos, S.; Trougakos, I.; et al. Discovery of novel cosmeuceutical agents from endophytic microorganisms of Spanish biodiversity. Planta Med. 2017. [Google Scholar] [CrossRef]
- Sykiotis, G.P.; Bohmann, D. Stress-activated cap’n’collar transcription factors in aging and human disease. Sci. Signal. 2010. [Google Scholar] [CrossRef] [PubMed]
- Wedel, S.; Manola, M.; Cavinato, M.; Trougakos, I.P.; Jansen-Durr, P. Targeting protein quality control mechanisms by natural products to promote healthy ageing. Molecules 2018, 23, 1219. [Google Scholar] [CrossRef]
- Morimoto, R.I.; Cuervo, A.M. Proteostasis and the aging proteome in health and disease. J. Gerontol. A Biol. Sci. Med. Sci. 2014. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Chen, C.H. Proteasome regulators: activators and inhibitors. Curr. Med. Chem. 2009, 16, 931–939. [Google Scholar] [CrossRef]
- Peyrat, L.A.; Eparvier, V.; Eydoux, C.; Guillemot, J.C.; Litaudon, M.; Stien, D. Betulinic acid, the first lupane-type triterpenoid isolated from both a Phomopsis sp. and its host plant Diospyros carbonaria Benoist. Chem. Biodivers. 2017. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classes of Natural Products | Bioactive Compound | Microorganism Classification (Kingdom; Species; Family) | Habitat | Biological Activity | CosIng Inventory3 | References | ||
|---|---|---|---|---|---|---|---|---|
| Antioxidants1 | Hydroxycinnamic acids | p-Coumaric acid | Fungi; A. austroafricanus; Trichocomaceae | Isolated from Z. officinale rhizome. | Antioxidant (DPPH, hydroxyl and nitric oxide radical-scavenging methods) Skin-whitening (inhibition of human tyrosinase and melanogenesis). | [31,32] | ||
| Ferulic acid | Antioxidant (DPPH, hydroxyl and nitric oxide radical-scavenging methods) Photo-protective (SPFs: 1.3). | Antimicrobial | ||||||
| Cinnamic acid | Antioxidant (DPPH, hydroxyl and nitric oxide radical-scavenging methods). | Perfuming & skin conditioning agent | ||||||
| Chlorogenic acid | Cyanobacteria; Dichothrix sp.; Rivulariaceae | Antioxidant (DPPH radical scavenging activity, IC50: 6.41 μg/mL ABTS radical scavenging activity, IC50: 13.15 μg/mL Deoxyribose protective activity, IC50: 8.53 μg/mL) | Antioxidant, skin-conditioning & skin-protecting agent | [26] | ||||
| Caffeic acid | Cyanobacteria; Aulosira fertilissima; Fortieaceae | Antioxidant (DPPH radical scavenging activity, IC50: 6.34 μg/mL ABTS radical scavenging activity, IC50: 18.04 μg/mL Deoxyribose protective activity, IC50: 4.76 μg/mL) | Antioxidant & masking agent | |||||
| Stilbenes | Resveratrol | Fungi; Alternaria sp.; Pleosporaceae | Isolated from grapes of Vitis vinifera | Antioxidant (inhibition of 8-OH-dG formation in DNA, IC50: 10.9) [33]2 Skin-whitening (inhibition of mushroom tyrosinase and of melanogenesis) [34]2 Preventive effect on lipid peroxidation [29]2 | Antioxidant & skin protecting agent | [30] | ||
| Fungi; S. cerevisiae; Saccharomycetaceae | [35] | |||||||
| Bacteria; E. coli; Enterobacteriaceae | [36] | |||||||
| Fungi; S. cerevisiae; Saccharomycetaceae | Isolated from Sugarcane. | [37] | ||||||
| Bacteria; E. coli; Enterobacteriaceae | Obtained from a Genetic Stock Center, New Haven, CT. | [36] | ||||||
| Bacteria; Bacillus sp.; Bacillaceae | Isolated from leaves of Populus alba L. | [38] | ||||||
| Biphenyls | Altenusin | Fungi; Botryosphaeria dothidea; Botryosphaeriaceae | Collected from stems of white cedar (Melia azedarach). | Antioxidant (DPPH radical scavenging activity, IC50: 17.6 μM). | [39] | |||
| 5’, Methoxy-6- methylbiphenyl-3,4,3’-triol | Antioxidant (DPPH radical scavenging activity, IC50: 18.7 μM). | |||||||
| Naphthoquinone spiroketals | Palmarumycin C3 | Fungi; Berkleasmium sp.; Dematiaceae | Isolated from healthy rhizomes of the medicinal plant Dioscorea zingiberensis. | Antioxidant (DPPH radical scavenging activity, IC50: 37.57 μM). | [40] | |||
| Benzoic acids | Gallic acid | Cyanobacteria; Limnothrix obliqueacuminata; Pseudanabaenaceae | Antioxidant (DPPH radical scavenging activity, IC50: 3.53 μg/mL ABTS radical scavenging activity, IC50: 8.85 μg/mL Deoxyribose protective activity, IC50: 7.84 μg/mL) | Antioxidant | [26] | |||
| Vanillic acid | Cyanobacteria; Mastigocladus laminosus; Hapalosiphonaceae | Antioxidant (DPPH radical scavenging activity, IC50: 416.7 μg/mL ABTS radical scavenging activity, IC50: 132.1 μg/mL Deoxyribose protective activity, IC50: 91.1 μg/mL) | Registered with no reported functions | |||||
| Thiol peptides | Glutathione | Fungi; P. pastoris; Saccharomycetaceae | Antioxidant (minimizes lipid peroxidation in cellular membranes and other such targets that is known to occur with oxidative stress) [41]2 Skin-whitening (tyrosinase inhibitor) [42]2 | Reducing agent | [43] | |||
| Bacteria; E. coli; Enterobacteriaceae | [44] | |||||||
| Carotenoids | Astaxanthin | Fungi; X. dendrorhous; Filobasidiaceae | Antioxidant (in vitro protection of biological membranes by an antioxidant mechanism) [45]2 Skin-whitening (inhibition of pigmentation, inhibition of melanin-generation) | Skin conditioning agent | [46] | |||
| Chlorophyta; H. pluvialis; Haematococcaceae | Obtained from Algal Culture Center, Plant Physiology Institute, University of Gottingen, Germany. | [47] | ||||||
| β-carotene | Bacteria; E. coli; Enterobacteriaceae | Antioxidant (αTEAC, FRAP and CL assay) [48]2 | Skin conditioning agent | [1] | ||||
| Lycopene | Antioxidant | [49] | ||||||
| Canthaxanthin | Bacteria; Brevibacterium; Brevibacteriaceae | Antioxidant (in vitro protection of biological membranes by an antioxidant mechanism) [45]2 Pigment | Cosmetic colorant | [50] | ||||
| Lutein | Chlorophyta; Muriellopsis sp.; Chlamydomonadales incertae sedis | Isolated from the Natural Park of the Empordá Marshes in Catalonia, Spain. | Antioxidant (Superoxide radical scavenging activity, IC50: 21 μg/mL Hydroxyl radical scavenging activity, IC50: 1.75 μg/mL Inhibition of lipid peroxidation: 2.2 μg/mL DPPH radical scavenging activity, IC50: 35 μg/mL ABTS radical scavenging activity: >100μg/mL Nitric oxide radical scavenging activity, IC50: 3.8 μg/mL) [51]2 Photo-protective (absorption of UVA rays) | Skin conditioning agent | [52] | |||
| Cis-canthaxanthin | Actinobacteria; D. maris; Dietziaceae | Isolated from soil sample collected from the Kargil district of Jammu and Kashmir, India. | Antioxidant (inhibition of ROS generation in THP-1 cells, >80%) | [53] | ||||
| Polysaccharides (PSs) | Exopolysaccharides (EPSs) | EPS fraction (PS-I); rhamno galactan | Fungi; F. solani; Nectriaceae | Isolated from A. scholaris. | Antioxidant (scavenging potency, IC50: 578.54 μg/mL) | [54] | ||
| Unknown EPS | Bacteria; B. cereus; Bacillaceae | Isolated from A. annua L. | Antioxidant: (DPPH radical scavenging activity, EC50: 3–5 mg/mL Superoxide radical scavenging activity, EC50: 2.6 mg/mL) | [55] | ||||
| Crude EPS | Bacteria; P. polymyxa; Paenibacillaceae | Isolated from S. japonica (Blume) Miquel. | Antioxidant (hydroxyl radical scavenging activity: 87.58% at 1 mg/mL) | [56,57] | ||||
| Mannose: fructose: glucose (2.6:29.8:1) | Antioxidant (hydroxyl radical scavenging activity: 76.73% at 1 mg/mL) | [56] | ||||||
| Mannose: fructose: glucose (4.2:36.6:1) | Antioxidant (hydroxyl radical scavenging activity: 68.55% at 1 mg/mL) | |||||||
| Deproteinized EPS | Algae; R. reticulata; Rhodellaceae | Isolated from freshwater. | Antioxidant (superoxide anion radical scavenging activity: 328.48 U/L) | [58] | ||||
| Mannose: galactose (89.4:10.6) | Fungi; Aspergillus sp.; Trichocomaceae | Isolated from leaves of Ipomoea pes-caprae L. | Antioxidant | Mannose as humectant and galactose as skin-conditioning agent | [59] | |||
| Rhamnose: glucose: glucuronic acid (2:2:1) | Bacteria; B. tropica; Burkholderiaceae | Isolated from Sugarcane. | Rhamnose as humectant & masking, glucose as humectant and glucuronic acid as humectant, chelating & buffering agent | [60] | ||||
| Unknown EPS | Chlorophyta; Graesiella sp.; Chlamydomonadales incertae sedis | Isolated from the hot spring ‘Ain Echffa’ (water temperature of 60 °C), Tunisia. | [61] | |||||
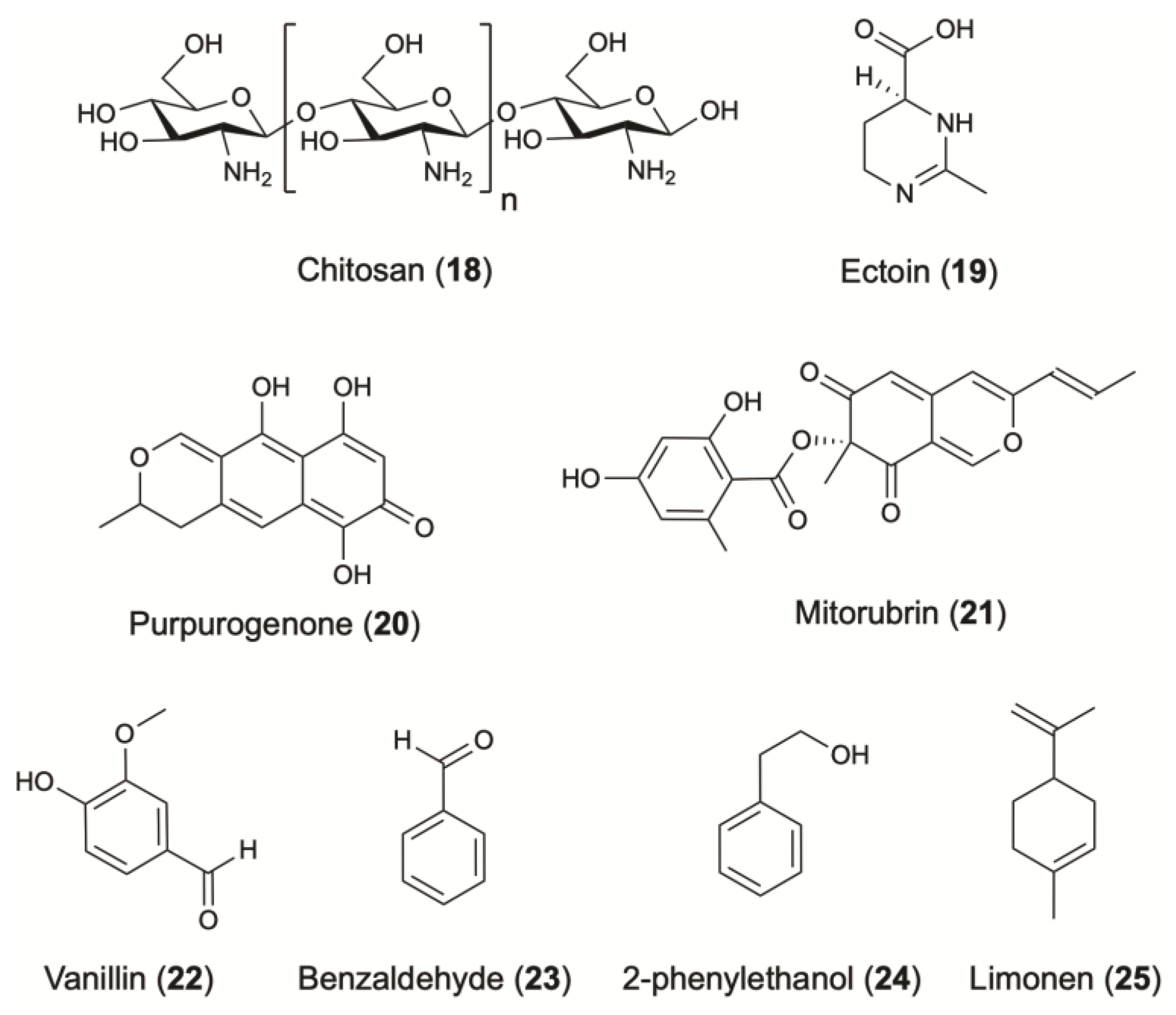
| Cell wall & exoskeleton PSs | Chitosan | Fungi; R. oryzae; Mucoraceae | Obtained from Culture Collection University of Gothenburg, Sweden. | Antimicrobial (higher activity on gram-positive bacteria, ex: Minimum inhibitory concentration (MIC) for S. aureus: 20 ppm) Moisturizing effect | Film forming & hair fixing agent | [62,63] | ||
| Fungi; R. japonicus; Mucoraceae | Shanghai Institute of Industrial Microbiology, China. | |||||||
| Fungi; M. indicus; Mucoraceae | Obtained from Culture Collection University of Gothenburg, Sweden. | [64] | ||||||
| Fungi; A. niger; Trichocomaceae | Isolated from the lichen R. montagnei. | [65] | ||||||
| Chitin-glucan | - | [66] | ||||||
| Photo-Protective agents1 | Melanins | - | Bacteria; S. kathirae; Streptomycetaceae | Isolated from soil samples. | Photo-protective (determination of SPF) | Skin protecting agent | [67] | |
| Bacteria; Bacillus safensis; Bacillaceae | [68] | |||||||
| Indole derivatives | Violacein | Bacteria; C. violaceum; Neisseriaceae | Isolated from soil sample collected from the vicinity of an oil refinery wastewater treatment plant in Negeri Sembilan, Malaysia. | Photo-protective: broad absorption band extended out to 700 nm [69] Antibacterial (more efficient on Gram positive bacteria, ex: S. aureus, IC50: 6.99 μM) | Antimicrobial, antioxidant & cosmetic colorant | [70] | ||
| Bacteria; Duganella sp.; Oxalobacteraceae. | Isolated from the glaciers of Tianshan, China. | [71] | ||||||
| Scytonemin | Bacteria; N. commune; Nostocaceae | Collected from sandy soil in Ningbo University, China. | Photo-protective: UV absorbent (UV-A and UV-B region) [72] Antioxidant (dose-dependent DPPH scavenging activity of 12%, 33%, and 57% at concentrations of 0.5, 1.0, and 2.0 mg/mL, respectively. Ascorbic acid used as positive control) | [73] | ||||
| Streptochlorin | Bacteria; S. roseolilacinus; Streptomycetaceae | Isolated from soil. | Skin-whitening (anti-tyrosinase activity, IC50: 9 mM) | [74] | ||||
| Prodigiosin | Bacteria; S. marcescens; Enterobacteriaceae | Isolated from fields contaminated with pesticides. | Photo-protective Antibacterial (more efficient on Gram positive bacteria, ex: S. aureus, IC50: 0.68 μM) | [75] | ||||
| Bacteria; Vibrio sp.; Vibrionaceae | Isolated from estuarine waters of the Northern Adriatic Sea. | [76] | ||||||
| Mycosporines | Palythine | Cyanobacteria; Lyngbya sp.; Oscillatoriaceae | Isolated from the bark of the rain tree Albizia saman (Jacq) Merr, Bangkok, Thailand. | Photo-protective (protection of HaCaT keratinocytes from solar-simulated radiation (SSR)-induced cell death), [77]2 | [78] | |||
| Asterina | Photo-protective | |||||||
| Unknown mycosporine-like amino acid | Antioxidant (DPPH scavenging activity of 14.5%, 53.0%, and 68.9% at 0.115, 0.230, and 0.460 mg/mL of MAAs, respectively. Ascorbic acid used as positive control) | |||||||
| Mycosporine-glutaminol-glucoside | Fungi; R. minuta; Sporidiobolaceae | Isolated from Patagonian natural environments | Photo-protective (UVB resistance of X. dendrorhous related to MGG production) | [79] | ||||
| Fungi; R. slooffiae; Sporidiobolaceae | ||||||||
| Fungi; R. lactosa; Sporidiobolaceae | ||||||||
| Fungi; C. liquefaciens; Tremellaceae | Isolated from a cold Arctic environment. | [80] | ||||||
| Mycosporine–glutamicol–glucoside | Fungi; C. cladosporioides; Cladosporiaceae | |||||||
| Skin-whitening agents1 | Pyrones | Kojic acid | Fungi; A. flavus; Trichocomaceae | Isolated from V. unguiculata. | Skin-whitening (anti-tyrosinase activity, IC50: 0.014 mM) | Antioxidant | [81] | |
| Fungi; A. oryzae; Trichocomaceae | - | [82,83] | ||||||
| Fungi; A. parasiticus; Trichocomaceae | Isolated from soil. | |||||||
| Fungi; A. candidus; Trichocomaceae | Isolated from soil. | |||||||
| Fungi; A. flavus; Trichocomaceae | Obtained from Department of Bioprocess Technology, University Putra, Malaysia. | |||||||
| (3R)-5-hydroxymellein | Endolichenic fungus | Isolated from the thalli of the lichen Parmotrema austrosinense (KoLRI no. 009806) collected from Jeju Island, Korea. | Photo-protective (damage recovery caused by UVB irradiation and inhibition of melanin synthesis) Antioxidant (DPPH radical scavenging, IC50: 30.8) | [84] | ||||
| Phenolic lactones | Ellagic acid (get by fungal bioconversion of ellagitannins) | Fungi; A. niger; Trichocomaceae | Obtained from DIA/UAdeC collection (Universidad Autonoma de Coahuila, Mexico). | Antioxidant (ABTS radical scavenging activity at 20 μg/mL: 93.9%). Skin-whitening (inhibition of melanogenesis) | Skin-conditioning agent | [85] | ||
| Carboxylic acids | Lactic acid | Fungi; R. oryzae; Mucoraceae | Obtained from CBS-Centraalbureau voor Schimmelcultures, Utrecht, The Netherlands. | Skin-whitening [86]2 pH adjuster Exfoliant | Humectant, buffering & skin-conditioning agent | [87] | ||
| Poly γ-glutamic acid | Bacteria; Bacillus sp.; Bacillaceae | Isolated from a soil sample collected at the Sugimoto campus of Osaka City University, Japan. | Skin-whitening Moisturizing (water-holding capacity: 56.9%) Antibacterial (more efficient on Gram-positive bacteria) | [88] | ||||
| Azelaic acid | Fungi; Malassezia furfur; Malasseziaceae | Skin-whitening (competitive inhibitor of tyrosinase: KI azelaic acid: 2.73x10-3 M) [89]2 Anti-bacterial and anti-acne effect Treatment of rosacea | Buffering & masking agent | [90,91] | ||||
| Tocopherols | Novel vitamine E succinate (bioconversion of vitamin E by modified Candida antarctica lipase B) | Fungi; C. antarctica; Saccharomycetacea | Skin-whitening effect | [92] | ||||
| Teichoic acids | Lipoteichoic acid | Bacteria; L. fermenti; Lactobacillaceae | Obtained from the National Collection of Type Cultures, Colindale, London. | Skin-whitening (inhibition of the intracellular activity of tyrosinase to 57.6% and 44.6% at 10 and 100 µg/mL of lipoteichoic acid) | [93] | |||
| Quinones | Arbutin undecylenic acid ester | Bacteria; B. subtilis; Bacillaceae | Skin-whitening (anti-tyrosinase activity, IC50: 4.10-4 M) | [94] | ||||
| Additives and other active ingredients1 | Purpurogenone | Fungi; P. purpurogenum; Trichocomaceae | Pigment | [95] | ||||
| Unknown Anthraquinone derivative | Fungi; P. oxalicum var. Armeniaca; Trichocomaceae | Obtained from soil. | [96] | |||||
| Peptides & amino acid derivatives | Ectoine | Bacteria; C. glutamicum; Corynebacteriaceae | Photo-protective (in vitro inhibition of UVA- induced signal transduction in human keratinocytes as well as inhibition of UVA-induced formation of mitochondrial DNA mutations in human dermal fibroblasts), [97]2 Moisturizing effect | Skin conditioning agent | [98,99] | |||
| Phenylalanine | Bacteria; E. coli; Enterobacteriaceae | New England Biolabs (NEB, Ipswich, MA). | Hair and skin conditioning effect | [100] | ||||
| Azaphilones | Mitorubrin | Fungi; P. purpurogenum; Trichocomaceae | Pigment | [95] | ||||
| Aromatic Aldehydes & alcohols | Benzaldehyde | Bacteria; E. coli; Enterobacteriaceae | New England Biolabs (NEB, Ipswich, MA). | Flavor, perfume (almond flavor) | Denaturant, flavoring, masking & perfuming agent | [100] | ||
| Benzyl alcohol | Bacteria; E. coli; Enterobacteriaceae | Flavor, perfume Preservative Bacteriostatic | Perfuming, preservative, solvent & viscosity controlling agent | |||||
| Vanillin | Bacteria; E. coli; Enterobacteriaceae | Flavor, perfume (vanilla flavor) | Masking agent | [101] | ||||
| Bacteria; E. coli; Enterobacteriaceae | [102] | |||||||
| Bacteria; B. fusiformis; Bacillaceae | Isolated from soil. | [103] | ||||||
| 2-phenylethanol | Fungi; A. gossypiii; Saccharomycetaceae | Flavor, perfume (rose flavor) Preservative | [104] | |||||
| Fungi; K. marxianus; Saccharomycetaceae | [105] | |||||||
| Terpenes | Limonene | Bacteria; E. coli; Enterobacteriaceae | Flavor, perfume (sweet citrus odor) | Deodorant, perfuming & solvent | [106] | |||
| Glycolipids | Rhamnolipid | Bacteria; P. aeruginosa; Pseudomonadaceae | Moisturizing and surfactant | Emollient, emulsifying & stabilizing agent | [107] | |||
| 2,3,4,2’-trehalose tetraester | Bacteria; R. erythropolis; Nocardiaceae | Isolated from soil. | Surfactant | [108] | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peyrat, L.-A.; Tsafantakis, N.; Georgousaki, K.; Ouazzani, J.; Genilloud, O.; Trougakos, I.P.; Fokialakis, N. Terrestrial Microorganisms: Cell Factories of Bioactive Molecules with Skin Protecting Applications. Molecules 2019, 24, 1836. https://doi.org/10.3390/molecules24091836
Peyrat L-A, Tsafantakis N, Georgousaki K, Ouazzani J, Genilloud O, Trougakos IP, Fokialakis N. Terrestrial Microorganisms: Cell Factories of Bioactive Molecules with Skin Protecting Applications. Molecules. 2019; 24(9):1836. https://doi.org/10.3390/molecules24091836
Chicago/Turabian StylePeyrat, Laure-Anne, Nikolaos Tsafantakis, Katerina Georgousaki, Jamal Ouazzani, Olga Genilloud, Ioannis P. Trougakos, and Nikolas Fokialakis. 2019. "Terrestrial Microorganisms: Cell Factories of Bioactive Molecules with Skin Protecting Applications" Molecules 24, no. 9: 1836. https://doi.org/10.3390/molecules24091836
APA StylePeyrat, L. -A., Tsafantakis, N., Georgousaki, K., Ouazzani, J., Genilloud, O., Trougakos, I. P., & Fokialakis, N. (2019). Terrestrial Microorganisms: Cell Factories of Bioactive Molecules with Skin Protecting Applications. Molecules, 24(9), 1836. https://doi.org/10.3390/molecules24091836